大花序桉组培快繁技术及其无性系造林效果的初步评价
2018-11-10唐再生王建忠莫继有熊涛张磊李丽芳兰俊
唐再生,王建忠,莫继有,熊涛,张磊,李丽芳,兰俊
大花序桉组培快繁技术及其无性系造林效果的初步评价
唐再生,王建忠,莫继有,熊涛,张磊,李丽芳,兰俊*
(广西国有东门林场,广西 扶绥 532108)
采用16 ~ 30 a生大花序桉优树伐桩萌芽,经无菌芽诱导、丛芽增殖、继代培养、生根培养、甁苗移栽、组培苗造林等技术环节的研究,筛选出一组适合大花序桉生长的培养基,建立以芽繁芽的组培快繁体系,获得实用的移栽和造林方法,形成一整套可面向规模化生产和推广造林的大花序桉组培快繁技术,其中无性系CL1的生根率达到88.9%,甁苗移栽成活率达到82.7%,试生产组培苗10万株,出圃造林近70 hm2,初具组培苗工厂化生产规模;对幼林观测测定结果表明:遗传性状稳定,再生植株能保持原株的优良遗传性状,林相整齐,速生丰产效果显著。
大花序桉;组培快繁;无性系造林
大花序桉()又叫昆士兰桉,天然分布在澳大利亚昆士兰州。它生长迅速,树高可达30 ~ 40 m,直径1 ~ 2 m,干形通直圆满,自然整枝良好,是一种重要实木利用的桉树树种。大花序桉的木材黄褐色,硬度高,纹理通直,结构均匀,耐久持重,锯板性能优良,是一种天然高贵木材[1]。因为生长快,干形好,材质优良,是商品经营培育大径材的优良树种。其轮伐期比尾巨桉()稍长,可以改善尾巨桉短周期经营超强收获而引起的生态环境问题,对于实现桉树产业可持续发展具有重要意义。广西国有东门林场从1983年中澳东门桉树项目开始,3次引种大花序桉,建立种源试验,结果表明大花序桉适合东门及附近地区生长,并表现出优良特性[2]。
目前大花序桉未能实现规模推广种植,主要原因在于苗木繁育困难。一是结实量少,种子发芽率低。造林用种依赖进口,育苗成本高昂,而且实生苗造林后林相不整齐,分化明显。二是大花序桉属于难生根树种,扦插生根极难。组培育苗虽然经过不少探索,陆续发表了一些大花序桉组培研究的报告,并取得了一些进展[3],但到目前为止,尚无面向生产推广的完整技术,未见大花序桉组培苗造林的报道。
育苗能否成功,决定了大花序桉的种植规模和能否持续发展。为打破这制约发展的瓶颈,东门林场林科所在2002年开展了大花序桉组培快繁技术的探索,并取得了初步成果,证明大花序桉能够通过组培进行无性繁殖[4]。2014年,在原来研究的基础上,再次进行深入研究,离体快繁技术取得成功,培育了无性系CL1和CL8,初步实现大花序桉组培无性系造林的目标。
1 材料与方法
1.1 试验材料
从东门林场1983年、1989年和2001年建立的大花序桉种源试验林中,选出优良种源/家系,再选出优良单株。单株选择时,注重生长量和干形指标,并考虑树体周围林木生长情况等综合因素,排除孤立木、林缘木、畸形木,通过伐倒促萌的方法,获得组培快繁原材料—优树萌芽,并对优树编号(CL1、CL2、CL3、…CL12)。
1.2 试验方法
1.2.1 优树萌芽的处理
1.2.1.1 观察记录萌芽形态特征及分类
观察记录优树萌芽的丛数,芽条数,枝和叶的形态特征,生长势等情况,汇总后分类。
1.2.1.2 外植体选择和表面消毒灭菌
对伐桩萌芽作必要的修剪、病虫害防治处理。当萌条适合作组培时,选取生长正常,无病虫害,无畸形,具有半木质化茎段的萌条,采集带回试验室,然后修剪去掉叶片和侧芽,留少许叶柄,切取长2 ~ 3 cm,含有潜伏芽的顶稍幼嫩茎段和半木质化茎段,用纯净水清洗10 min,然后在超净工作台上,用75%酒精浸泡10 ~ 20 s,无菌水清洗3次;再用0.1%升汞溶液浸泡5 ~ 8 min,无菌水清洗5次。用经过灭菌处理的干滤纸吸去茎段表面附着的水分,在接种碟上,用手术刀或手术剪将茎段切割成含有1 ~ 2个潜伏芽,长度1.0 ~ 1.5 cm的小段,接种到无菌芽诱导培养基上。观测记录污染、褐化、萌芽其情况,累计多次诱导的结果。
1.2.2 外植体无菌芽促萌方法
诱导无菌芽选用改良MS培养基。添加6-BA 1.0 mg∙L-1+NAA 0.2 mg∙L-1,并根据不同无性系特点,调整6-BA和NAA的浓度,蔗糖3%,琼脂4.0 ~ 5.0 g∙L-1,按琼脂强度大小调整用量。pH值调至5.8 ~ 6.2。接种后全暗培养9 ~ 12 d,萌发芽点后,移至自然散射光下培养,光照强度由弱逐渐增强。
1.2.3 无菌芽继代增殖培养
1.2.3.1 预备试验
通过改良MS培养基,添加6-BA 0.4 mg∙L-1+NAA 0.2 mg∙L-1,蔗糖3%,琼脂4.0-5.0 g∙L-1,pH值5.8 ~ 6.2,并根据继代苗生长情况调整6-BA和NAA的浓度。
1.2.3.2 深化试验
对于预备试验培育后生长不理想的无性系,选用改良ER(微量元素用MS的量),WPM做基础培养基,添加6-BA、NAA和KT,改变光照条件,接种后先暗培养10 d,试验材料作幼化处理等。
1.2.3.3 增殖倍数的确定方法
因为继代培养的芽苗大小不一致,各代芽苗数出现重叠,用切割芽苗数来计算,理论上比较准确,但在实际中难以操作。所以采用连续继代培养3代,以最后一次继代苗瓶数和起始瓶数,按=3计算增殖倍数。为最后一次继代苗瓶数;为初始继代苗瓶数;为增殖系数。要求接种时每瓶芽苗数尽量一致。
1.2.4 生根培养
1.2.4.1 预备试验
剪切1.2 ~ 1.5 cm长的单芽,接种到生根培养基1/2改良MS + NAA 1.0 mg∙L-1+ IBA 0.5 mg∙L-1,蔗糖3%,琼脂4.0 ~ 5.0 g∙L-1,pH值5.8 ~ 6.2,每个无性系接种40瓶,每瓶10株。
1.2.4.2 深化试验
选用B2(广西钦州市林科所研制)、1/2改良ER、1/2改良MS作为基础培养基,添加ABT1、IBA作为生根激素,按照生根率和出根特征,调整激素浓度,再添加根太阳(一种分子信号促生根物质)进行试验。
1.2.5 大花序桉离体培养和植株再生的培养条件
1.2.5.1 培养温度
培养室温度26 ± 2℃,生根苗在大棚炼苗时温度18 ~ 35℃。
1.2.5.2 光照条件
继代培养以自然散射光为主,每天光照时间为10 ~ 12 h,光照强度1 500 ~ 2 000 lux,刚接种时弱光条件下培养,随后逐步增强。自然光照不足时,用人工光源补足。生根培养中,光照也由弱光条件向强光条件递增。大棚炼苗时,光照强度1 500 ~ 7 000 lux。
1.2.6 生根甁苗移栽
1.2.6.1 炼苗和洗苗方法
开盖后,灌注冷开水到培养基再炼苗。洗苗时,将培养基放入水中捏碎,取出苗根。
1.2.6.2 栽培基质的选择
①育苗用轻型基质,椰糠+泥炭土(5:3);②黄心土+蛭石(5:1);③轻基质(椰糠+泥炭土)+黄心土(3:1)。基质在移栽前2 d,用0.3%高锰酸钾溶液消毒。
1.2.7 大花序桉组培苗造林试验
1.2.7.1 试验设计
选大花序桉组培无性系苗CL1造林,用大花序桉实生苗作对照,株行距2 m × 4 m,宽行窄株布置。以组培苗3行,实生苗3行交替块状种植,机耕全垦整地。
1.2.7.2 试验数据调查
造林2个月后调查统计成活率,造林1 a后调查测定树高和胸径,观测记录幼树形态和干形,每年测定树高胸径1次。
2 结果与分析
2.1 优树萌芽处理的效果
2.1.1 优树萌芽形态特征观察与分类
对优树萌芽观察后表明,砍伐的优树都具有萌芽能力。萌芽条具有区别于尾巨桉等其他桉树的固有特征,各优树间又有明显的差异。结合观察间伐的大量大花序桉萌芽形态特征,大致可划分分为3类。由表1可知,大花序桉萌芽形态差异大,表明各优树之间基因性状的差异也大,这为在群体中选择到相对易于繁殖的个体提供了依据。进行分类组培研究,有可能获得通过观察萌芽特征,初步判断繁殖能力大小的方法。
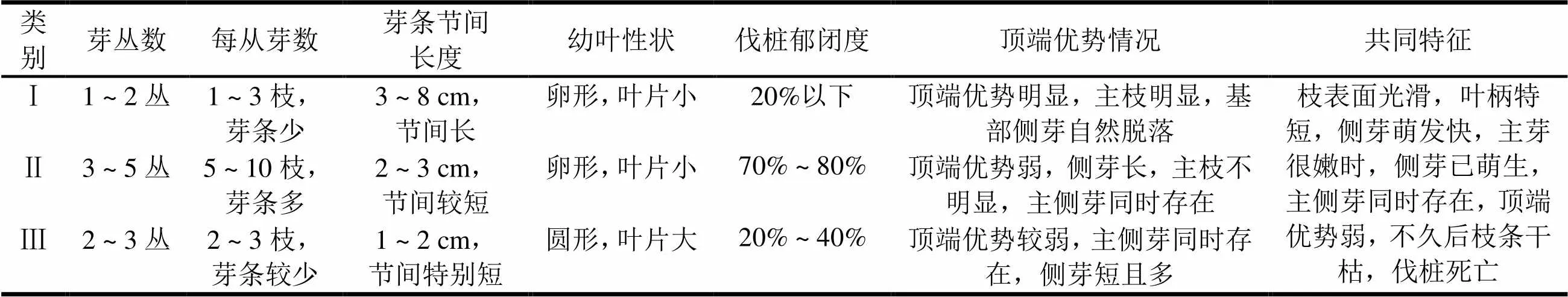
表1 大花序桉优树萌芽形态特征及分类
2.1.2 外植体选择与灭菌方法对无菌芽诱导的影响
按照外植体在萌芽条上节位的高低和木质化程度,分为3类:A、萌条上部,仅含有芽点或小侧芽的嫩茎段;B、萌条中段,已萌发较长侧芽的半木质化茎段;C、萌条中下部,已萌发较长侧芽的木质化茎段。依据萌条木质化程度采用相应的消毒灭菌时间,统计多次消毒灭菌后12 d的检查结果。表2显示,含有芽点或小侧芽的A类外植体,因为太嫩,木质化程度低,经短时间消毒灭菌处理后,外植体褐化枯死率高,而且萌芽质量不理想。C类外植体比较老,接种后污染率偏高,茎切口处培养基褐化比较重,萌芽质量差。B类外植体褐化、污染率较低,相对比较易萌芽,萌芽质量比较好,是大花序桉诱导无菌芽宜选用的外植体,但第一批侧芽已经萌发,并在消毒灭菌前已修剪,外植体萌发的是第二批潜伏芽。这与尾巨桉有一定差别,也增加了无菌芽诱导的难度。
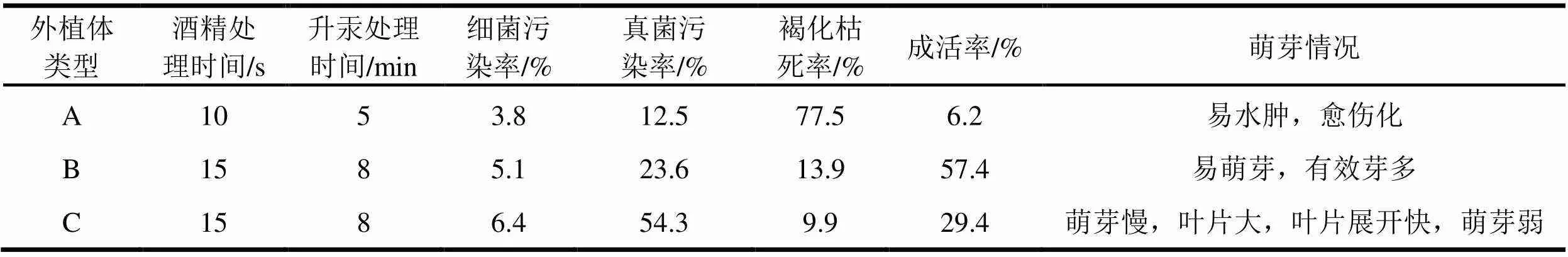
表2 大花序桉不同类型外植体消毒灭菌情况表
2.2 诱导培养基对外植体萌芽的影响
选用半木质化茎段B类外植体接种,除去污染、褐化枯死者,以接种后36 d为界。累积多次外植体诱导无菌芽的调查统计结果,得出各类优树萌芽情况。从表3可知,不同类型优树的外植体其萌生无菌芽存在较大差异。Ⅱ类易萌生丛芽,Ⅲ类萌生的无菌芽节间太短,仅见叶不见茎,几乎成无效芽。Ⅰ类芽少,节间长。体现了优树萌芽的特征,即伐桩芽多,外植体萌芽也多。
为改善第Ⅲ类优树外植体萌芽的不足,调整了生长调节剂浓度与配比,将诱导培养基调节剂改为改良MS+6-BA 1.0 mg∙L-1+NAA 0.5 mg∙L-1;虽然结果表明总的萌芽数仍然少,但提高了无菌芽质量,节间伸长,有效芽数增加。所以,针对不同伐桩的萌芽特征,需选择相应的诱导培养基。Ⅲ类优树萌芽作外植体诱导无菌芽,宜选用细胞分裂素和生长素比值较低的培养基,以6-BA 1.0 mg∙L-1+NAA 0.5 mg∙L-1较合适。

表3 各类型大花序桉优树外植体萌发无菌芽情况
2.3 增殖培养基对丛芽增殖和继代培养的影响
2.3.1 预备试验结果
从未污染外植体上切取无菌芽,第一代转接时需连带一小段母枝,利于无菌芽生长,转入继代增殖培养基后,萌芽逐渐增多,形成侧芽和腋芽共同增殖的态势。表4显示,3类不同的原材料形成的继代芽苗外观形态差异明显。瓶苗的生长情况反映了伐桩萌芽的生长特点。伐桩萌芽多,生长旺盛者,组培繁育后也表现增殖多,生长旺盛,具有基因依赖性。
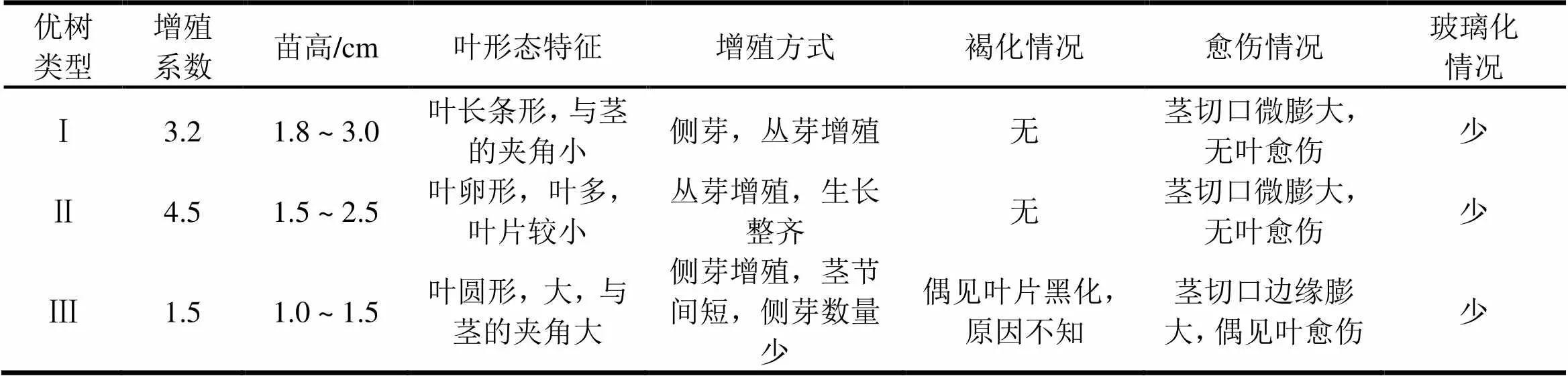
表4 各类大花序桉优树无菌芽继代增殖情况
不断继代培养,Ⅰ类材料保持稳定的增殖,因为激素在植物体内的积累,芽苗逐渐增多,增殖系数有一定提高,五代之后,处于稳定状态。Ⅱ类材料保持高增殖状态,五代之后丛生芽过多,处于团芽中间部分的芽苗变得细弱,需要降低激素浓度与比值,用6-BA 0.2 mg∙L-1+NAA 0.1 mg∙L-1。结果向人为需要的方向发展,反应较灵敏。当需要接种生根时,对于增殖系数过高,芽苗细弱者,还需要作一次壮苗培养,才有利于生根。Ⅰ、Ⅱ类材料适合在预备试验的增殖培养基上。Ⅲ类材料则表现出增殖系数低,芽苗节间难伸长,苗高1.5 cm以下,叶片大,在单位面积的培养基上芽苗数量太少等问题,需要作进一步深化试验。
2.3.2 深化试验结果
2.3.2.1 改变基础培养基和激素浓度
表5的数据表明,改良MS培养基,调高6-BA/NAA的比值,Ⅲ类材料的侧芽增多,但芽苗短小,如未开的桃花着生在叶腋处,多数是无效芽。需要用6-BA和NAA浓度比值为1的壮苗培养基培养,芽苗的茎节间才伸长,而且高生长仍有限,低于1.5 cm。继续往下培养,出现侧芽少,增殖系数不足1.5,苗高叶达不到要求。改变基础培养基,改良ER与改良MS的差异不明显,用WPM培养基,芽苗生长表现不如改良MS,芽苗增殖有限,难以拔节长高。以改良MS作基础培养基,添加KT 0.2mg∙L-1,芽苗有一定的高生长,侧芽也有伸长,拔节作用较为明显,但很少见有丛生芽,仍以侧芽增殖为主。
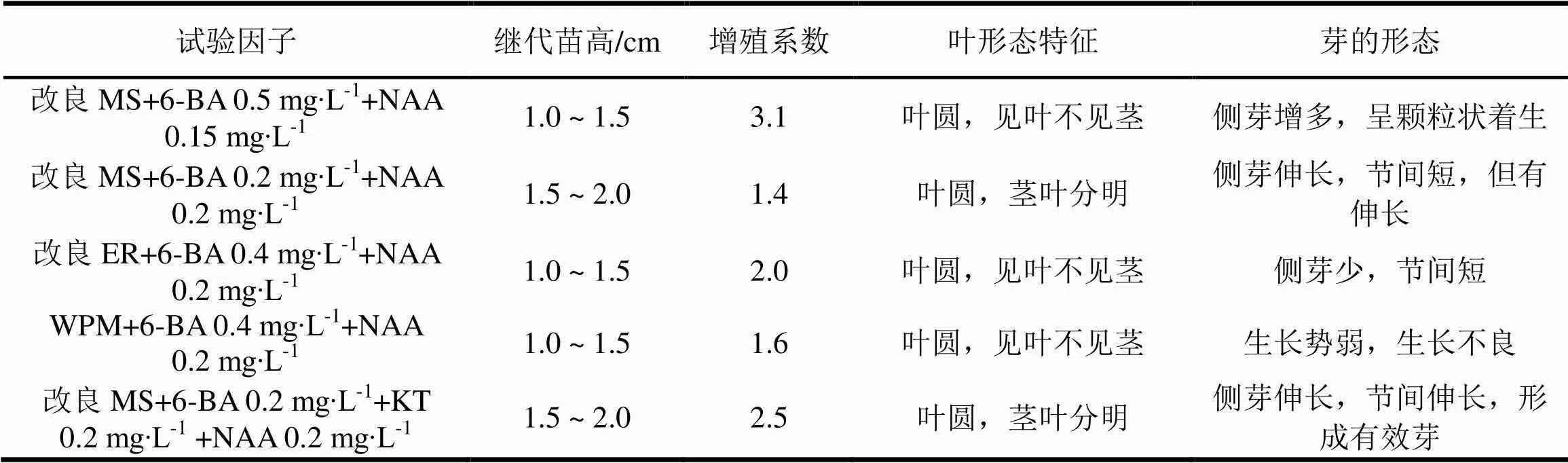
表5 3类大花序桉优树无菌芽继代增殖情况
2.3.2.2 改变光照条件以促进苗高生长
接种继代后,在继代初期实行全暗培养,在7 ~ 12 d内,芽苗新生芽白化,稍有伸长,老叶仍绿,但部分老叶黑化死亡,包括部分嫩梢黑化死亡,总的比率达到20%;见光培养后,维持原状,高生长不显著。接种继代后,用日光灯作光源,四周遮光,光线从顶部照射,利用植物的向光性促进拔节伸长。其结果亦不理想,高生长有限,而且苗木生长势比较弱。部分叶片黑化死亡,原因不知。
2.3.2.3 进行幼化处理促进苗木生长
考虑到优树树龄大,萌条位置虽然是在根茎附近,但需要作幼化处理,以解除原材料对无菌芽的束缚。切取芽苗茎尖,长度0.5 cm作继代培养。结果新芽仍不能长高,增殖也同原来一样,叶片圆形,叶片与茎的夹角比较大,只是叶片变小了一些。
以上试验结果表明,Ⅲ类材料增殖不够和苗高有限的特点,具有基因依赖性,对外部条件改善难以奏效。针对没有丛芽、仅有侧芽,而且节间不伸长的特点,采用无菌短枝型分化方式[5],将继代芽切成带侧芽的小段,转接到含有KT的改良MS培养基,通过不断调整细胞分裂素与生长素的比例以及浓度,经反复多次培养之后,苗木增殖和苗高生长都得到改善,增殖系数达到2.5倍,苗高达到1.5 ~ 2.0 cm,能反复继代培养,结果比较稳定,芽苗为有效芽,能进行批量生产组培苗,规模生产组培苗的方案是先利用较高细胞分裂素和生长素浓度比例的培养基,形成较多数量的芽点,再用较低比例的培养基让小芽伸长,这样做会使培养周期延长,但能够达到一定的增殖系数并产生有效芽,进入批量生产,从而解决这类材料的快繁问题。实现这个目标,与KT的使用有关,单纯用6-BA,茎节间难以伸长,另一个原因可能是经多代培养后,芽苗内源激素与外源激素逐渐平衡有关。
2.4 生根培养基对大花序桉生根的影响
2.4.1 预备试验结果
经多次继代培养,芽苗生长稳定之后,选取健壮的单芽切割,接种到预备试验培养基,培养20 d后,调查生根情况,结果见表6。初步试验结果显示,大花序桉Ⅱ、Ⅲ类材料,有少许出根,相对Ⅰ类材料易于生根,但与尾巨桉DH32-29相比,生根率很低,而且根系质量与苗的质量都远不如尾巨桉。大花序桉不同品系之间,差异比较明显。Ⅰ类材料,易形成类根组织且比较突出。类根组织外形上与根相似,易与真根混淆,其主要特征:长0.3 ~ 1.0 cm,圆柱形,表面光滑,无根毛,无侧根,但有分叉。当激素浓度较高时,形成鹿角状类根组织,与愈伤团有差别。移栽到育苗基质后,成活率在5%以下。在预备培养基上,Ⅰ类材料未生根,Ⅱ、Ⅲ类材料生根率很低,离规模生产要求差距较大。
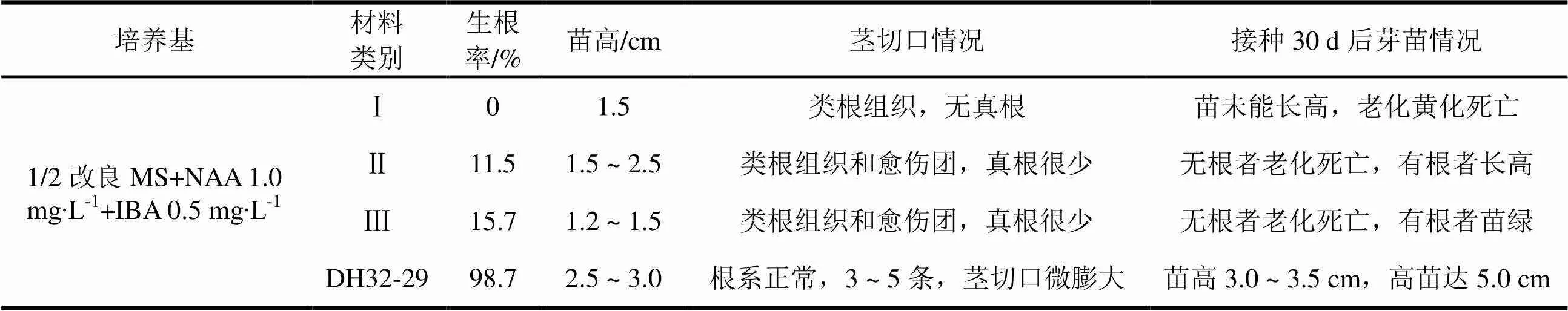
表6 三种大花序桉在预备生根培养基的生根情况
2.4.2 深化试验结果
2.4.2.1 生根剂的调试
以1/2改良MS培养基和ABT1促根剂的配合,利于大花序桉产生真根。其根比较细长,但结实,苗的主干也变得结实,叶片更绿。单因子多浓度梯度试验,以0.2 mg∙L-1为一个梯度,浓度范围0.3 ~ 3.0 mg∙L-1。结果表明,ABT10.5 mg∙L-1以下,没有根出现,茎切口处无反应;当浓度达到2.5 mg∙L-1时,切口处愈伤特别大,叶片碰到培养基时,也形成愈伤组织或类根组织。用ABT1促根,浓度应选择在0.5 ~ 2.5 mg∙L-1之间,多次对不同无性系调试试验,综合评价后,以1.0 ~ 1.5 mg∙L-1有比较好的出根率,Ⅱ、Ⅲ类材料出根率高于Ⅰ类材料,Ⅱ类材料出根率最高,超过60%。
再用ABT1和IBA协同促生根处理,按新配方多浓度配制培养基,接种后最佳处理表现为茎切口微膨大,出根条数增加,每株出3 ~ 5条真根。Ⅰ类材料有15%的出根率,Ⅲ类材料出根率上升至78.5%。主要原因是ABT1和IBA组合,充分发挥了互补作用。ABT1主要成分是吲哚乙酸(IAA)和萘乙酸(NAA),关键成分是新型植物生长调节剂。ABT1的生理活性与NAA、IBA等调节不同,它具有通过强化调控植物内源激素含量和重要酶的活性,促进生物分子合成的作用,诱导植物不定根的形态建成[6]。IBA促根作用强,但在植物体内移动性比较差,所以用生理活性强的ABT1和低浓度的IBA配合,具有相互促进与互补功效,当浓度使用得当,就产生好的生根效果。
2.4.2.2 新型生根剂的应用
在ABT1和IBA协同作用下,生根率大幅提升,但芽苗切口有一定的愈伤组织,部分生根和愈伤组织相距很近,当愈伤组织在洗苗时发生脱落,那么根在受到外力碰撞时,也易于脱落。为解决这一不足,添加新型生根剂—根太阳,取得了较好效果。
根太阳是一种新型的分子信号诱导生根剂,它可以通过诱导植物体内相关基因的表达,促进根原基发出新根。在大花序桉诱导生根过程中,有一典型特征就是诱导茎的皮层发根,离茎切口比较远。这类根在洗苗时不易脱落,移栽成活率较高,只要浓度应用得当,效果明显。
2.4.2.3 改良基础培养基
选取Ⅲ类材料的健壮芽苗,每瓶接15株,每个处理共接20瓶,接种到B2、1/2改良ER、1/2改良MS培养基,添加ABT1和IBA,其生根情况如表7。B2培养基的优点是根细而结实,但显著的缺点就是苗高生长不足。1/2改良ER与1/2改良MS差异不大,苗能够长高,但木质化不够,根系虚大。在B2和1/2改良MS培养基的无机盐浓度水平上,调整部分矿物质营养浓度,特别是NH4+的含量,结果取得较好的效果,出根率提高,苗高达到2.0 cm以上,根系比较结实。再经过基础培养和激素浓度的配合调试,最终形成大花序桉生根培养基:1/2改良MS+ABT1+IBA+根太阳。

表7 不同基础培养基的生根情况
2.4.3 驯化作用对生根的影响
随着大花序桉无菌芽继代培养代数的增加,切取健壮芽苗接种到相同配方的生根培养基上,生根率和根的条数都有逐步提高的趋势。结果见表8。生根的驯化作用,可能来源于继代苗的驯化。随着继代次数的增加,芽苗生长稳定,而且对培养基成分变化的缓冲作用增强了,苗木生长整齐,大小一致,只是在生根培养基中表现突出,易于观测。在继代初期,选取规格一致的优势芽苗接种生根,生根率很低,到45代之后,即诱导无菌芽,继代培养后2 a时间,生根率提高,出根条数增加,其中表现较好的CL1号的平均生根率达到88.9%,出根数3 ~ 5条,苗高生长也达到要求。在不同无性系之间,驯化作用的表现又有差异,Ⅰ类材料在继代多代之后,生根率提高的幅度仍然有限,没有达到规模化生产的要求。
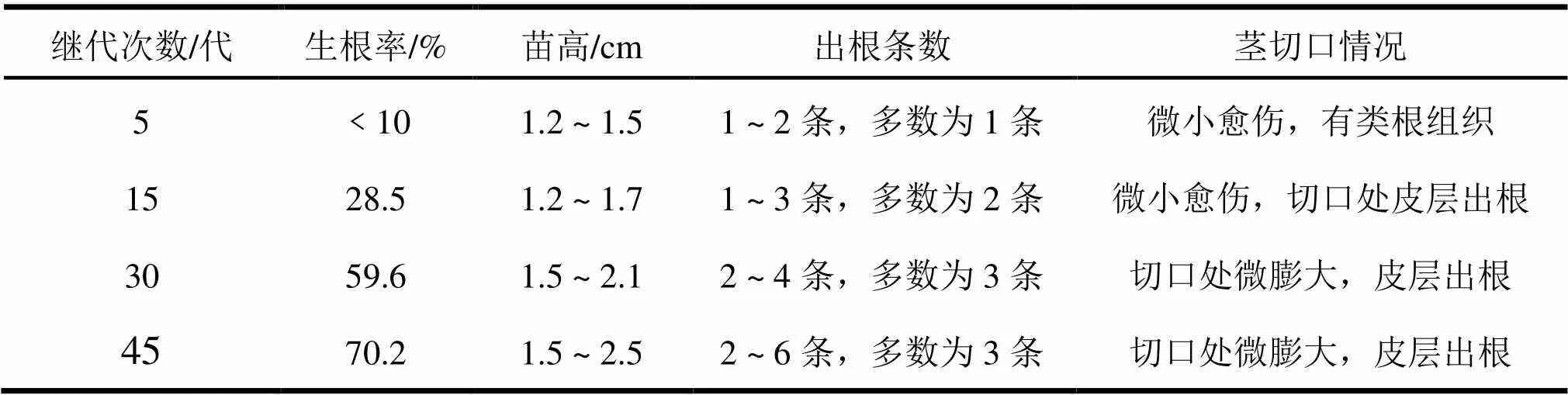
表8 大花序桉CL1随着继代次数增加生根情况统计表
2.5 大花序桉生根苗移栽
2.5.1 炼苗洗苗方法对移苗质量的影响
移栽前通过揭开瓶盖,降低瓶内湿度,有利于增强小苗叶片革质化,减小蒸腾对水分的消耗,增强小苗对自然环境的适应能力,大花序桉开盖炼苗后,出现叶片萎焉现象,移栽后成活率降低。考虑到因为蒸腾作用强,吸干培养基原有水分,加注自来水,然后再炼苗,结果培养基污染严重,这表明培养基中还有较多糖和其他有机物未吸收利用完,容易滋生细菌。因此只能加冷开水,炼苗2 d。
在洗苗去除培养基时,很容易断根,按照洗尾巨桉的方法,一手拿苗,一手捏碎培养基,再向上提苗的步骤,断根约30%以上,验收生根率达到90%的甁苗,洗苗移栽后仅剩60%左右。所以必须一手拿苗,一手托住培养基,放在水中捏碎培养基,轻轻晃动,将培养基去掉,虽然这样比较费时,但根系保存完整,移苗后成活率高。
2.5.2 基质对大花序桉移栽的影响
在春季,移栽后45 d,检查苗木成活率和生长情况(表9)。
轻基质移栽后易出现苗木枯死且较多,黄心土作基质成活率高,但是生长缓慢,长势不佳。轻基质+黄心土是最好基质,苗木易成活,生长也好。说明黄心土和轻基质对大花序桉根的生长影响比较大,原因可能在于土壤胶体贮存水分,易于被根系吸收,轻基质提供部分养料加上空隙大,能够提供氧气,而桉树生根过程中需要大量的氧气[7]。
2.5.3 生理性立枯病对大花序桉移栽的影响
小苗移栽后,前期用薄膜覆盖保湿,或者仅盖上遮阴网。在晴朗干燥的天气,小苗大面积干枯死亡,大花序桉实生苗也出现类似的情况,外观症状与立枯病十分相似。苗高5 ~ 8 cm的大苗也有立枯现象。采用杀菌剂无效,而且基质本身已严格消毒处理,深入检查发现,死亡植株没有病原物,干枯死亡系生理性失水,采取叶面补水措施后,症状缓解,死亡株数减少。提前采取补水措施,立枯病症状可以控制或者不发生。原因可能是大花序桉小苗叶面积大,蒸腾作用过强和根系吸水和输水功能障碍引起,导致水分亏缺。在实际生产中,为防止生理性立枯危害,移栽小苗生长稳定后,将小苗连同无纺布基质杯植入直径12 cm的黄心土基质杯中,即可减少生理性立枯病的发生。此外,病理性立枯病对大花序桉组培苗移栽危害也比较严重,是一种土壤中病原菌侵染引发的病害,表现为幼苗植株基部腐烂、矮化、黄化,用手拔植株时根部易断,多发生在春末夏初,空气潮湿的天气,采用精甲∙噁霉灵水剂或噁霉灵水剂进行灌根或喷施防治,有较好的效果。
2.5.4 虫害防治
用轻基质移栽小苗后,发现一种虫害症状:前1 d苗木生长正常,第2 d出现萎焉死亡,不倒伏。当拔苗出来观测时,见苗茎与根部断开,而且断裂处有缺口,夜间观测,可看到白色蠕虫啃食苗茎。经查证,这是尖眼蕈蚊的幼虫为害,其成虫俗称“小黑飞”。虫源可能来自轻基质原料中的泥炭土,成虫产卵其中,在育苗大棚内湿热环境中孵化,幼虫啃食小苗。另一个原因是大花序桉幼苗茎秆肥嫩,易被蠕虫啃食,同时移栽的尾巨桉小苗鲜见受害。防治措施:①用黑光灯诱杀成虫,在大棚内悬挂杀虫灯诱杀已孵化的成虫。②用杀虫剂毒杀幼虫。在移栽的育苗杯基质表面,撒上长制缓释药肥颗粒,毒杀幼虫。两种措施配合使用,效果显著,虫害得到有效控制。
2.6 大花序桉组培苗造林
2.6.1 造林成活率
选择大雨后,林地充分湿透后造林,如果雨量不够,种植后用人工灌水补足,使土壤湿润,造林20 d后检查成活率98%以上,偶有死株,需及时补植。大花序桉组培苗叶面积较大,蒸腾作用强,种植难度比较大,有条件可以采用保水剂,每株0.5 ~ 1.0 kg,效果更好,只要造林措施得当,成活率高。
2.6.2 早期生长量调查结果
造林14个月后测定胸径和树高,以实生苗作对照,结果见表10。
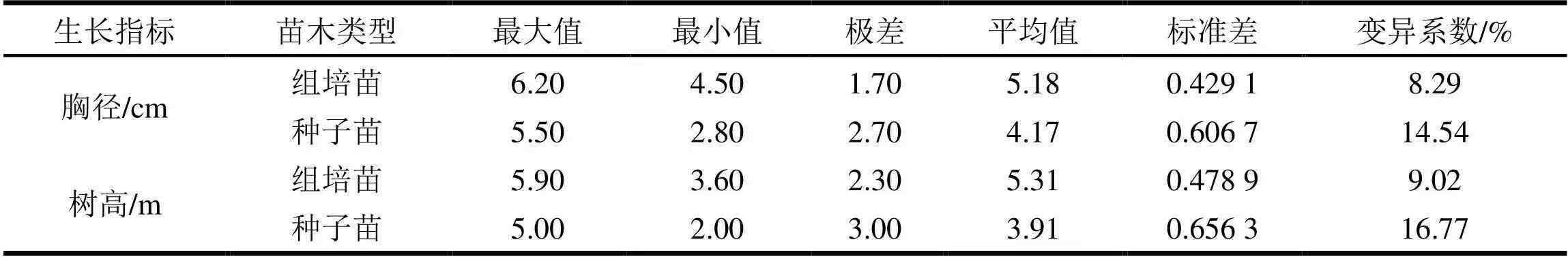
表10 大花序桉组培苗和实生苗林分生长情况表
从表10可知,大花序桉组培苗林分胸径和树高平均值为5.18 cm和5.31 m,而大花序桉实生苗林分胸径和树高平均值为4.17 cm和3.91 m,通过胸径和树高的极差和变异系数分析,大花序桉组培苗林分比实生苗林分在生长量、整齐度方面具有明显优势。原因是组培苗有优树无性繁殖成苗,具有好的基因优势并且基因相同,所以苗木生长快而且整齐一致,实生苗来源于不同基因的个体,生长量大小不一致,出现分化。
2.6.3 幼林形态观察
2.6.3.1 林相整齐,树体大小一致
组培无性系林分,通过肉眼观察,生长整齐,树高相差很小,树体大小一致,表现了无性系林分的特征;而实生苗林分,则已表现出分化趋势,主要在树体大小的差异比较明显,冠幅的大小,树高都有差异,总体参差不齐。
2.6.3.2 各植株形态正常,干形好
逐株观察,组培苗植株呈尖塔形,主干明显,枝条分布均匀,没有出现畸形植株,而实生苗林分出现部分干形不良植株,表现在1.8 ~ 2.0 m处出现丛生枝,形成没有主干的状况,近地面主干生长硕大,另外还见到少量树高和胸径生长量很小的植株。
2.6.3.3 枝条多,自然整枝慢
这是组培苗和实生苗共有的形态特征,基部枝条多,枝条比较长,易受台风影响造成风倒。对于台风影响频繁的地区,需要人工整枝。
从幼林形态观察,大花序桉组培苗表现出大花序桉固有的形态特征,无生长畸形株,林相整齐,干形好,保持了优树原株的优良特性。
3 结论与讨论
大花序桉不同种源/家系的优树,萌芽形态差异比较大,按照芽条形态特征分类,各类材料在诱导无菌芽、继代增殖培养、生根培养等方面都表现了一定的差异。特别在生根性状上,有较多的基因依懒性。按照不同类型的萌芽,有针对性的选择材料繁殖,有利于组培繁育获得成功。
大花序桉萌芽条的侧芽萌生快,顶端优势比较弱,诱导无菌芽时,只能依靠第二批潜伏芽获得无菌繁殖体系。因此,需要选育细胞分裂素浓度和生长素浓度比值较低的培养基,有利于无菌芽的萌发。
继代增殖培养过程中,Ⅰ类和Ⅱ类材料易于建立继代增殖体系,芽苗增殖系数高,生长质量好。Ⅲ类材料鲜有见丛生芽,增殖系数小,叶片大,苗高生长不足。通过添加激动素KT,按照无菌短枝型方式,以侧芽增殖,可以达到规模生产的要求。
生根培养中,大花序桉普遍表现生根困难,组培快繁的瓶颈在生根环节,这是由大花序桉本质特性决定的。大花序桉组培快繁能否进入规模化生产,繁殖材料的基因类型起决定作用,通过外源激素促进生根起辅助作用,根太阳这一新型分子信号促根剂具有一定的促生根作用。所以,选择易于繁殖的无性系,调试合适的培养基,是目前大花序桉组培快繁的实用选择。
大花序桉组培苗移栽也比较困难,主要是受到立枯病的危害。这是大花序桉特有的生理性病害。实生苗也有发生,导致育苗损失严重。原因在于叶片蒸腾作用过强和根系吸水障碍引起,解决办法就是保湿。采用喷雾状水和使用较大的育苗杯,使根系扩大,很好地解决了这一难题。移栽大花序桉的育苗基质,采用轻基质和黄心土按3:1比例混合,有利于小苗成活和生长。但目前在制作育苗杯/袋时,不能够机械化作业,需要人工装填基质,增加了育苗成本,当大规模生产大花序桉组培苗之际,应定制专门的机械来制杯。
大花序桉组培苗造林后,表现出无性系造林的优势,林相整齐,能保持优树的良好特性,生长快、干形好、无分化。无性系造林是大花序桉下一步发展的方向。通过侧芽和顶芽形成以芽繁芽的组培快繁方式,没有经过愈伤组织途径,无脱分化过程,获得的再生植株遗传性状稳定[8],实现大规模造林是可靠、安全的。
大花序桉组培快繁是可行的,培养技术在不断完善,生根率不断提高,但大花序桉难生根这一特性没有改变。在同一培养基中,接种相同质量的芽苗,结果有出根和不出根的,生根情况差异大。再加上移栽环节对技术要求高,要实现大花序桉组培苗无性系造林,需要提高技术水平,还要提高管理水平,做到产量规模化,操作专业化,管理精细化。
本研究中Ⅱ类和Ⅲ类优树已有无性系进入规模生产组培苗,两个无性系已累计生产组培苗10万株,出圃造林累计近70 hm2,用于推广试验。Ⅰ类优树因为生根率较低,多数是类根组织,需要继续研究,提高生根率才能实现无性系造林。下一步需要继续改进和完善Ⅱ类和Ⅲ类材料培养基配方,探索环境条件对组培繁育的影响,不断提高苗木质量,降低育苗成本。继续探索Ⅰ类材料的促根技术,最终实现对每一株优树都能够繁育成无性系造林的目标。
[1] 李昌容,陈奎,周小金,等.大花序桉研究现状与发展趋势[J].桉树科技,2012,29(2):40-46.
[2] 王建忠,熊涛,张磊,等.25年生大花序桉种源生长与形质性状的遗传变异及选择[J].林业科学研究,2016,29(5):705-713.
[3] 姚瑞玲,王胤,项东云,等.大花序桉组织培养研究进展[J].安徽农业科学,2009,37(29):14030-14031.
[4] 唐再生.大花序桉芽器官离体组培快繁技术研究初报[J].广西林业科学,2006,35(S1):22-23+33.
[5] 曹孜义,刘国民.实用植物组织培养技术教程[M].兰州:甘肃科学技术出版社,1999.
[6] 唐庆兰.大花序桉组织培养的研究[D].南宁:广西大学,2006.
[7] 宋建英.邓恩桉的组织培养和植株再生[J].林业科学,2010,46(6):138-145.
[8] 曾炼武.巨尾杂交桉芽器官离体培养快速繁殖的研究[J].广西林业,1990(S1):16-23.
Tissue Culture Techniques forandField Evaluation Resultant Clones
TANG Zaisheng, WANG Jianzhong, MO Jiyou, XIONG Tao, ZHANG Lei, LI Lifang, LAN Jun
(,,)
Buds from 16 to 30 year old elite trees ofwere used to study aspects of tissue culture propagation of clones of these trees, including: sterile bud induction, cluster bud proliferation, successive transfer culture,rooting, transplanting into nursery growing tubes and early field establishment. In this study suitable tissue culture media for growth ofwas identified along with protocols necessary for rapidtissue culture propagation, starting from buds collected off the original trees. These protocols, along with methods developed for subsequent nursery culture, can provide for rapid propagation ofelite trees and are suited to the needs of large-scale production to service plantation establishment programs. The rooting rate of the clone CL1 reached 88.9%, transplanting survival rate reached 82.7% and initial test production yielded one hundred thousand tissue culture seedlings, enough for establishment of 70 hm2of commercial plantations. Early results from field trials show that thetissue cultured clones can keep the superior traits of the original mother trees (i.e. the ortets), with the young clonal trees showing remarkably fast early growth.
; tissue culture; clonal afforestation
S722.3+7
A
广西林业科技项目“大花序桉良种组培快繁技术研究”(桂林科研〔2015〕第18号)。
唐再生(1972— ),男,学士,高级工程师,主要从事桉树育种、组培育苗及优良无性系开发及推广研究。
兰俊(1981— ),男,硕士,高级工程师,主要从事桉树遗传改良及无性系开发研究,E-mail:57714915@qq.com
