越冬期邓恩桉嫁接种子园叶绿素与丙二醛动态变化
2018-11-10邓紫宇郭东强陈健波刘媛
邓紫宇,郭东强,陈健波,刘媛
越冬期邓恩桉嫁接种子园叶绿素与丙二醛动态变化
邓紫宇1,2,3,郭东强1,2,3,陈健波1,2,3,刘媛1,2,3
(1.广西壮族自治区林业科学研究院,广西 南宁 530002;2.国家林业局 中南速生材繁育实验室,广西 南宁 530002;3. 广西优良用材林资源培育重点实验室,广西 南宁 530002)
以邓恩桉嫁接种子园抗寒能力不同且开花的5个家系为材料,测定其越冬期叶片中叶绿素与丙二醛(MDA)含量。结果表明:自然降温过程中,叶绿素含量随着温度的下降而降低,MDA含量随着温度的下降而增加,各家系MDA含量增加幅度不同;春季随着气温的回升,叶绿素含量略有上升,而MDA含量呈下降趋势。通过研究具开花能力的邓恩桉家系越冬期叶绿素与MDA含量动态变化,为选育抗寒能力强的邓恩桉F1代提供理论依据。
邓恩桉;嫁接种子园;叶绿素;丙二醛;自然降温
邓恩桉()原产澳大利亚,天然分布区面积极小,具有速生、耐寒优良性能,是重要的纸浆材和实木用材树种[1-4]。因其具有一定的耐寒性能,我国从20世纪80年代开始引种邓恩桉,并在南方桉树种植北缘区表现良好[5]。
邓恩桉无性繁殖困难,市面上仍未见组培苗,在我国大部分引种区,邓恩桉基本不开花结实,造林用的种子全部依赖进口,造林成本的增加限制了它在我国的进一步推广。邓恩桉的优良性能使其成为各方学者重点研究对象,主要集中在组织培养技术研究[6-7]、木材材性性能测定[8-11]、抗寒生理生化指标测定[12-16]及引种适应性调查[17-20]等方面。笔者所在的研究团队曾以开花与未开花的邓恩桉植株为材料,比较两者生理生化特性和光合生理,发现开花植株渗透调节物质明显高于未开花植株,且光合系数较小[21]。在此研究的基础上,本文针对具开花能力的邓恩桉植株,对其越冬期的表现,测定其叶片中叶绿素含量与MDA含量的变化规律,为邓恩桉嫁接种子园低温伤害与抗性机制研究提供参考,并为选育速生抗寒的邓恩桉F1代提供科学依据。
1 材料与方法
1.1 试验地概况
邓恩桉嫁接种子园位于广西环江县华山林场雅龙分场,2010年7月建立。试验地地理位置与气候因子参考文献[21]。
1.2 试验材料
以邓恩桉嫁接种子园具开花能力且生长正常、花蕾较多的5个家系为材料(编号A、B、C、D、E),采集树冠中上部不同方向当年生侧枝的完整功能叶,每个家系选取5株,每株30枚叶片,于2014年11月至2015年3月每月采集一次,以2014年11月第一次采集样品为对照。
1.3 试验方法
叶绿素含量的测定采用丙酮乙醇混合液浸提法,MDA含量的测定采用硫代巴比妥酸(TBA)比色法[22]。
2 结果与分析
2.1 邓恩桉嫁接种子园气候变化
2014年11月至2015年3月邓恩桉嫁接种子园的气候变化见表1,2015年1月为越冬期最低温月,平均低温为3℃,极端低温达-1℃。

表1 采样期邓恩桉嫁接种子园气候变化
2.2 叶绿素含量动态变化
由表2可知,越冬期,邓恩桉嫁接种子园5个家系叶片中叶绿素的含量随着温度的降低而降低,开春气温回暖,叶绿素的含量逐渐上升。2015年1月气温达到最低时,叶绿素含量降到最低,与对照相比,5个家系叶片中叶绿素含量分别下降了9.60%、5.08%、5.08%、9.70%、6.82%。叶绿素主要有叶绿素a和叶绿素b两种,不溶于水,但易溶于丙酮、酒精等有机溶剂。随着气温的下降,叶绿素a含量下降,叶绿素b含量上升,随着气温上升,叶绿素a含量上升,叶绿素b含量下降。正常情况下,叶绿素a与叶绿素b的比值约为3:1,随着气温下降,叶绿素a/b值下降,随着气温上升,叶绿素a/b值上升。
2.3 MDA含量动态变化
越冬期,邓恩桉嫁接种子园5个家系叶片中MDA的含量随着温度下降而升高,开春气温回暖,MDA的含量逐渐下降(表3)。2015年1月气温达到最低时,MDA的含量上升到最高,与对照相比,5个家系叶片中MDA的含量分别上升了53.15%、49.50%、61.65%、49.63%、56.20%。
表3 越冬期邓恩桉嫁接种子园5个家系MDA含量动态变化 μmol·g-1
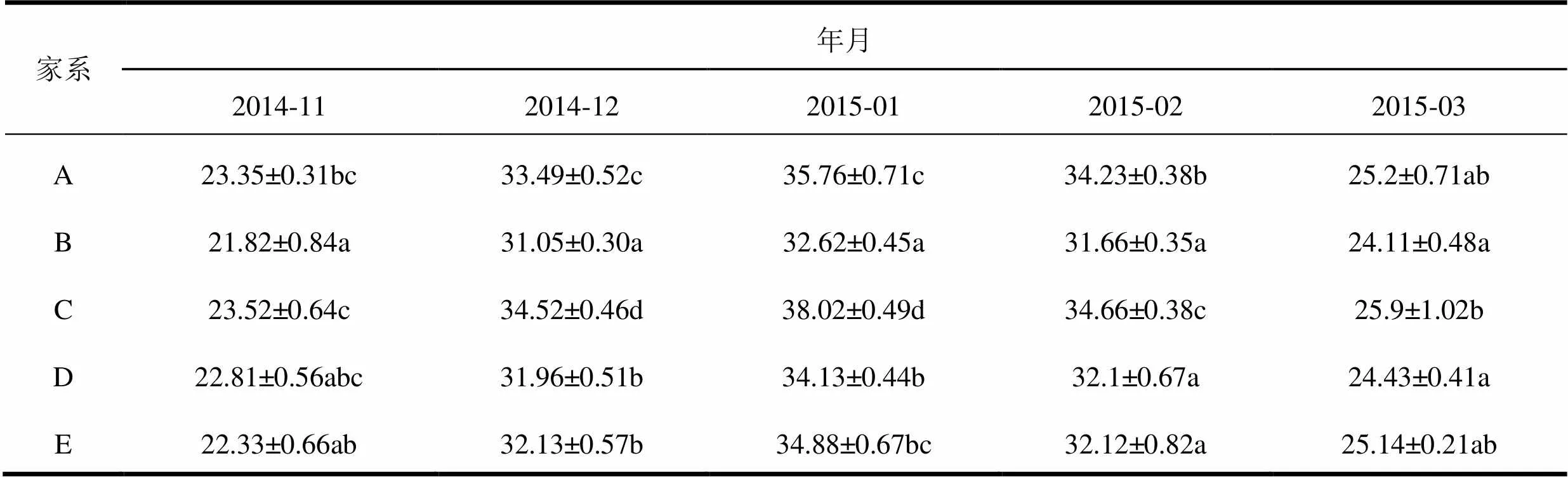
家系年月 2014-112014-122015-012015-022015-03 A23.35±0.31bc33.49±0.52c35.76±0.71c34.23±0.38b25.2±0.71ab B21.82±0.84a31.05±0.30a32.62±0.45a31.66±0.35a24.11±0.48a C23.52±0.64c34.52±0.46d38.02±0.49d34.66±0.38c25.9±1.02b D22.81±0.56abc31.96±0.51b34.13±0.44b32.1±0.67a24.43±0.41a E22.33±0.66ab32.13±0.57b34.88±0.67bc32.12±0.82a25.14±0.21ab
注:表中数据为平均值±标准差;小写字母表示不同家系差异性比较(˂0.05),下同。
表2 越冬期邓恩桉嫁接种子园5个家系叶绿素含量动态变化 mg·g-1
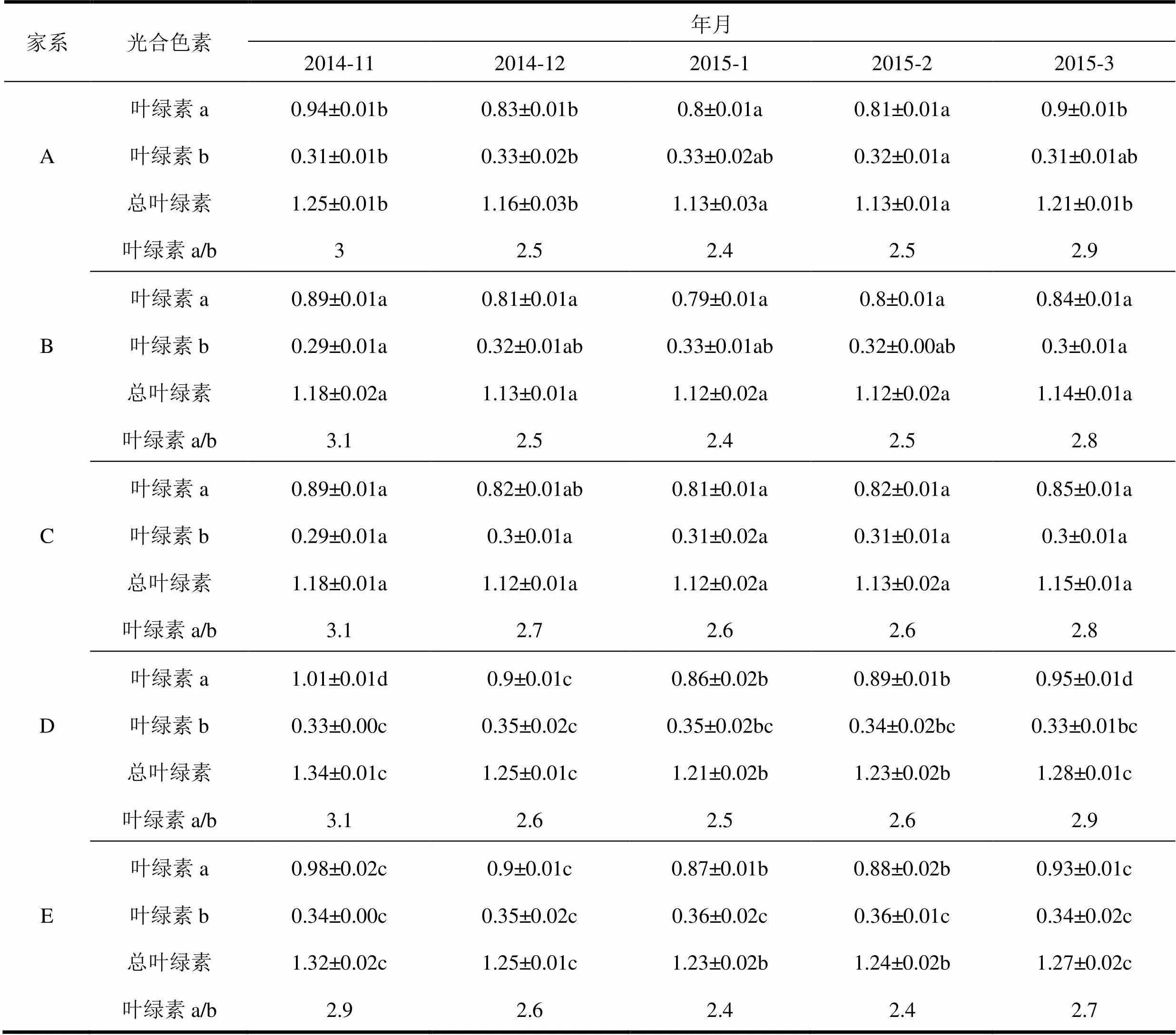
家系光合色素年月 2014-112014-122015-12015-22015-3 叶绿素a0.94±0.01b0.83±0.01b0.8±0.01a0.81±0.01a0.9±0.01b A叶绿素b0.31±0.01b0.33±0.02b0.33±0.02ab0.32±0.01a0.31±0.01ab 总叶绿素1.25±0.01b1.16±0.03b1.13±0.03a1.13±0.01a1.21±0.01b 叶绿素a/b32.52.42.52.9 叶绿素a0.89±0.01a0.81±0.01a0.79±0.01a0.8±0.01a0.84±0.01a B叶绿素b0.29±0.01a0.32±0.01ab0.33±0.01ab0.32±0.00ab0.3±0.01a 总叶绿素1.18±0.02a1.13±0.01a1.12±0.02a1.12±0.02a1.14±0.01a 叶绿素a/b3.12.52.42.52.8 叶绿素a0.89±0.01a0.82±0.01ab0.81±0.01a0.82±0.01a0.85±0.01a C叶绿素b0.29±0.01a0.3±0.01a0.31±0.02a0.31±0.01a0.3±0.01a 总叶绿素1.18±0.01a1.12±0.01a1.12±0.02a1.13±0.02a1.15±0.01a 叶绿素a/b3.12.72.62.62.8 叶绿素a1.01±0.01d0.9±0.01c0.86±0.02b0.89±0.01b0.95±0.01d D叶绿素b0.33±0.00c0.35±0.02c0.35±0.02bc0.34±0.02bc0.33±0.01bc 总叶绿素1.34±0.01c1.25±0.01c1.21±0.02b1.23±0.02b1.28±0.01c 叶绿素a/b3.12.62.52.62.9 叶绿素a0.98±0.02c0.9±0.01c0.87±0.01b0.88±0.02b0.93±0.01c E叶绿素b0.34±0.00c0.35±0.02c0.36±0.02c0.36±0.01c0.34±0.02c 总叶绿素1.32±0.02c1.25±0.01c1.23±0.02b1.24±0.02b1.27±0.02c 叶绿素a/b2.92.62.42.42.7
3 结论与讨论
3.1 自然降温与叶绿素含量变化
叶片是植物进行光合作用的主要器官,而叶绿体是植物进行光合作用的主要细胞器。高等植物的光合色素包括叶绿素和类胡萝卜素,类胡萝卜素比较稳定,叶绿素易受外界因素(降温、矿质元素缺失、病虫害等)及内在因素(叶片衰老)的影响而被破坏甚至降解。酶参与大部分叶绿素的生物合成过程,温度影响酶活动继而影响叶绿素的合成,低温抑制叶绿素的形成。一般情况下,2 ~ 4℃是植物叶绿素形成的最低温度,40℃是植物叶绿素形成的最高温度,而30℃左右是植物叶绿素形成的最适温度[23]。自然降温过程中,邓恩桉嫁接种子园5个家系叶绿素含量均下降,与雪松()[24]、红豆杉()[25]、猴樟()和芳樟()[26]等研究结论一致,低温下叶绿体结构遭到破坏、叶绿素合成受到限制,叶绿素含量下降。刘建等[15]利用人工低温处理邓恩桉幼苗,12 h和24 h后叶绿素含量上升,48 h后叶绿素含量下降,总体呈下降趋势,这是邓恩桉幼苗对低温的一种适应性表现。本研究是在整个自然降温过程中邓恩桉叶绿素含量的累积变化,叶绿素含量的减少,吸收的光能相应地减少,过剩光能产生的活性氧造成的伤害也减轻,这是邓恩桉对低温胁迫的反应也是长期适应低温的响应。
叶绿素a呈蓝绿色、叶绿素b呈黄绿色,叶绿素b分子与绝大部分叶绿素a分子具有收集和传递光能的作用,少数特殊状态的叶绿素a分子具有将光能转换为电能的作用,正常植株叶绿素a/b值稳定,约为3:1,使其叶片呈绿色[23]。自然降温过程中,邓恩桉嫁接种子园5个家系叶绿素a含量下降而叶绿素b含量上升,叶绿素a/b值下降,与雪松[24]研究结论一致。而在尾巨桉()[27]、猴樟和芳樟[26]等研究中,叶绿素、叶绿素a和叶绿素b均呈下降趋势,叶绿素b含量的上升与低温下叶绿素b的补偿效应有关,a/b值的下降,邓恩桉部分叶片绿色变浅或出现淡黄色。
3.2 自然降温与MDA含量变化
正常情况下,植株体内代谢平衡,自由基的产生与清除处于动态平衡,逆境胁迫下,平衡破坏,活性氧的产生大于清除,活性氧积累导致细胞膜脂过氧化,生物膜结构破坏,植株正常生长受到影响。MDA是细胞膜脂过氧化产物之一,是评价膜系统过氧化程度的重要指标。自然降温过程中,邓恩桉嫁接种子园5个家系叶片中MDA的含量均随着温度的下降而升高,与靖长柏等[16]、刘建[28]和黄月华等[29]的研究结论一致。低温胁迫下,邓恩桉植株体内活性氧代谢失衡,抗氧化调节能力下降,膜脂过氧化作用加剧,MDA含量上升。
自然降温过程中,邓恩桉嫁接种子园5个家系MDA含量变化存在一定差异,表明5个家系对低温的适应性不同。家系C增加幅度最大,其膜脂过氧化程度最严重,细胞膜受损也最严重,家系B与家系D增加幅度最小,其膜脂过氧化程度最轻,细胞膜受损也最轻,家系B与D在5个家系中抗寒能力较强,在今后邓恩桉核心嫁接种子园建设中可作为耐寒品种的选择。
3.3 存在不足与建议
邓恩桉应对低温的生理生化反应复杂多样,本研究仅从叶绿素含量与MDA含量探讨越冬期邓恩桉嫁接种子园的响应,而保护酶活性的变化有待作进一步深入研究。抗寒锻炼与合理施肥可以有效提高植株的抗寒能力,在今后工作中应加强这方面的研究。当前,分子生物学技术不断发展,植物抗寒研究已进入分子水平,邓恩桉抗寒的分子机理与抗寒基因工程研究也势在必行。
[1] BOLAND D J,BROOKER M I H,CHIPPENDALE G M,et al.Forest trees of Australia(5th ed) [M].Collingwood:CSIRO Publishing,2006.
[2] THOMAS D,HENSON M,JOE B,et al.Review of growth and wood quality of plantation-grownMaiden[J].Australian Forestry,2009,72(1):3-11.
[3] 王豁然.桉树生物学概论[M].北京:科学出版社,2010.
[4] 祁述雄.中国桉树:第2版[M].北京:中国林业出版社,2002.
[5] 罗建中.耐寒桉树良种—邓恩桉[J].桉树科技,2002 (2):1-8.
[6] 燕丽萍,夏阳,毛秀红等.邓恩桉的组织培养[J].林业科学,2011,47(5):157-161.
[7] 谭柏韬,李茂娟,邓少华.邓恩桉再生系统的研究[J].中国农学通报,2013,29(31):32-35.
[8] 郭东强,叶露,周维,等.2个种源邓恩桉木材纤维特性及变异[J].浙江农林大学学报,2014,31(4):502-507.
[9] 任世奇,罗建中,彭彦,等.17年生邓恩桉两个种源木材密度与干缩性研究[J].亚热带植物科学,2010,39(2):5-9.
[10] 叶露,NICHOLS J D, RAYMOND A D,等.邓恩桉生长特性与生长应变研究[J].安徽农业科学,2013,41(10):4409-4410,4418.
[11] 卢翠香,陈建波,刘媛,等.邓恩桉生材含水率、年轮宽度及木材密度研究[J].桉树科技,2014,31(2):23-27.
[12] 裘珍飞,曾炳山,刘英,等.应用电导法评价5个桉树抗寒性[J].广东林业科技,2011, 27(3):27-31.
[13] 郭祥泉,吴奇智,洪伟,等.应用电导率探讨邓恩桉不同优株的抗寒性[J].亚热带植物科学,2012,41(1):41-44.
[14] 郭祥泉.用膜透性、SOD酶活性和膜脂脂肪酸成分探讨林木抗寒性[J].西南林业大学学报,2012,32(4):6-11.
[15] 刘建,项东云,陈健波,等.低温胁迫对桉树光合和叶绿素荧光参数的影响[J].桉树科技,2009,26(1):1-6.
[16] 靖长柏,张利阳,童再康,等.自然低温胁迫对3种桉树生理生化特性的影响[J].防护林科技,2010(6):18-19, 35.
[17] 卢新瑛.2011年初春持续低温对邓恩桉生长的影响[J].亚热带农业研究,2011,7(2):100-104.
[18] 周修权,曹学良.桉树在郴州地区的抗寒性调查分析[J].湖南林业科技,2012,39(4):54-56.
[19] 郭祥泉,郭祥堆.邓恩桉在闽北生长规律与适应性探讨[J].西北林学院学报,2010,25(2):50-55.
[20] 郭祥泉,陈慈禄,洪伟,等.邓恩桉优株在闽北适应性种植决策探讨[J].西南林业大学学报,2012,32(3):26-30.
[21] 李昌荣,郭东强,周维等.邓恩桉种子园开花生理生化特性比较[J].中国农学通报,2014,30(25):20-23.
[22] 李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2000.
[23] 潘瑞炽.植物生理学:第5版[M].北京:高等教育出版社,2006.
[24] 郭爱华,左宝峰,姚延梼,等.自然降温对雪松叶片中叶绿素及电导率的影响[J].山西农业大学学报(自然科学版),2005,25(4):393-395.
[25]刘彤,祝佳媛,李鹏,等.秋冬季自然降温过程中东北红豆杉幼苗的生理生化特性[J].北京林业大学学报,2013,35(2):51-56.
[26]王宁,姚方,袁美丽,等.自然降温对猴樟和芳樟叶绿素荧光参数的影响[J].河南农业大学学报,2013,47(4):414-419.
[27] 刘奕清,李会合,陈泽雄.尾巨桉幼苗对低温胁迫的生理生化反应[J].福建林业科技,2007,34(4):24-26.
[28] 刘建.两种桉树对低温胁迫的响应机制研究[D].南京:南京林业大学,2008.
[29] 黄月华,徐建民,余雪标.低温胁迫对桉树代谢的影响[J].热带农业科学,2005, 25(5):24-28.
Dynamic Changes in Chlorophyll and Malondialdehyde in a Grafted Seed Orchard ofDuring Winter
DENG Ziyu1,2,3, GUO Dongqiang1,2,3, CHEN Jianbo1,2,3, LIU Yuan1,2,3
(1.2.3.)
Five families ofthat had been selected for flowering competence and a range of cold tolerance in a grafted seed orchard, had their leaves sampled at various times during the course of the winter months to examine their chlorophyll and malondialdehyde (MDA) contents. Chlorophyll contents were found to decrease as temperatures decreased as winter progressed, whilst MDA contents increased, but the increases in MDA differed among the five families. Then, with the onset of spring, chlorophyll contents increased while MDA contents decreased as the temperatures started to rise. This research on the dynamic changes in chlorophyll and MDA contents in the trees of thegrafted seed orchard during winter will help provide a theoretical basis for the breeding of this species and production of high quality F1seeds.
; grafted seed orchard; chlorophyll; MDA; natural temperature decrease
S718.43
A
广西科学研究与技术开发项目(桂科1347004-3);广西林科院基本科研业务费项目(林科201408号)。
邓紫宇(1984— ),男,硕士,工程师,主要从事林木遗传育种研究,E-mail:365204914@qq.com.
