黄瓜MADS-box基因家族生物信息学分析及CsMADS-box AGL62的克隆
2018-11-08周秀艳秦智伟
赵 阳 周秀艳 辛 明 秦智伟
(农业农村部东北地区园艺作物生物学与种质资源创制重点实验室,东北农业大学园艺园林学院,黑龙江哈尔滨 150030)
黄瓜是我国保护地栽培的第一大蔬菜作物,2015年全国栽培面积125.8万hm2(沈辰 等,2017)。黄瓜的性别分化、花期的早晚对黄瓜的产量及上市的早晚有着不可忽视的影响,这些特点决定了黄瓜品种的经济价值。目前,黄瓜性别分化、花期及果实发育成为了研究的热点。MADS-box基因是一类广泛存在于植物中序列特异的同源异型基因,参与调控花和果实发育及成熟的多个过程。MADS-box基因作为转录因子含有一段高度保守的DNA结合结构域,通过该结构域MADS-box基因识别并结合在下游目标基因的特定靶位点,从而调控下游目标基因的表达(张则婷和李学宝,2007)。植物含有众多MADS-box基因,如拟南芥有100多个MADS-box基因(De Bodt et al.,2003;Parenicov á et al.,2003),它们形成一个庞大的基因家族。根据系统进化关系、基因结构和蛋白结构的不同,MADS-box 基因分为I型(Type I)和II型(Type II)两大类型(Becker & Theissen,2003)。Type I可 进 一 步 分 为Ma、Mβ、Mγ 和Mδ;Type II可进一步分为MIKCC和MIKC*(Henschel et al.,2002)。植物I型MADS-box基因研究较浅,已有的研究表明,该类型的基因主要参与雌配子体、胚胎和种子的发育进程,并与不同物种间生殖器官分界的决定有关(Masiero et al.,2011)。植物II型MADS-box基因研究比较深入,该类型的基因大多与花形态形成有关,如已知的ABCDE模型。黄瓜中共发现43种MADS-box基因。已有的研究表明黄瓜中不含有FLC、AGL12和Bs 3种亚型的MADS-box基因,并且除GL亚型外,黄瓜其他亚型的MADS-box基因与拟南芥等其他植物中基因具有相似的功能(Gan et al.,2014)。
本试验在对黄瓜几组不同花期的杂交组合田间花期调查的基础上,由转录组测序并通过荧光定量验证的方法获得了1个黄瓜MADS-box家族的基因,通过生物信息学分析的手段来预测该基因的功能,以期为进一步基因功能验证奠定基础。
1 材料与方法
1.1 试验材料
以黄瓜自交系D0708-2为父本(雌雄异花同株),自交系D1158♀-2(强雌系)、D1104-2-4(全雌系)、D0528-2(雌雄异花同株)为母本配制杂交组合。F1分别为东农812(D1158♀-2×D0708-2)、C15-114(D1104-2-4×D0708-2)、C17-31(D0528-2×D0708-2)。父母本及F1均由东北农业大学园艺园林学院黄瓜课题组提供,采用穴盘育苗并且于2017年7月幼苗两叶一心时定植于东北农业大学园艺站塑料大棚。每小区2行,每行14株。随机区组设计,3次重复。
逆转录酶、DNA聚合酶、d NTP、T/A克隆载体均购自TaKaRa公司,TRIzol购自Invitrogen公司,质粒小量提取试剂盒、DNA凝胶回收试剂盒和荧光定量染料均购自北京全式金生物技术有限公司。
1.2 试验方法
1.2.1 田间性状调查及转录组测序 于黄瓜开花初期、根瓜期、盛果期时每小区随机抽取3株进行定株调查,分别考察各组合不同时期的雌花数、雌花节率、第1雌花节位及最高发育节位,最高发育节位指最顶端开放的花(不分雌花雄花)所在位置距离茎尖的节位。开花初期指黄瓜单株至少3朵花开放,小区内至少50%植株开花的时期;根瓜期指有50%黄瓜植株开始采收商品瓜的时期;盛果期指黄瓜单株商品瓜产量达到最大的时期(路洪凤,2014)。试验数据采用Excel 2010进行整理,DPS 7.05软件进行统计分析。
通过田间试验结果挑选1个组合进行转录组测序。测序及分析交由华大基因(深圳,中国)使用Illumina HiSeq平台完成,参照数据库 为 NCBI数 据 库(https://www.ncbi.nlm.nih.gov/genome/?term=cucumber)。
1.2.2 引物设计与合成 使用软件Primer Premier 5.0(Premier Bio-soft International,Palo Alto,CA)设计克隆基因全长引物AGL62,并通过http:/biodb.swu.sdu.cn/qprimerdb网站下载荧光定量引物qAGL62,使用CsEF1α基因作为内参(Lu et al.,2018)。克隆引物及荧光定量引物见表1。1.2.3 CsMADS-box AGL62的克隆及荧光定量 PCR扩增体系为20.0μL,其中:10×PCR Buffer(含 20 mmol·L-1Mg2+)2.0 μL,dNTPs(10 mmol·L-1)2.0μL,Taq 酶(2U)0.2μL,cDNA模板2.0μL,上下游引物各0.5μL,最后加12.8μL的ddH2O。PCR反应程序为:94 ℃预变性5 min;94 ℃变性 30 s,55 ℃退火 1 min(表 1),72℃延伸1 min,35个循环;72 ℃延伸10 min;4 ℃保存。PCR产物经1%琼脂糖凝胶电泳分析后,回收目的片段,连接到pGEM-T克隆载体上,采用热激法转化到Trans1-T1感受态细胞中,在氨苄抗性平板上进行蓝白斑筛选,挑白色菌落摇菌后送至苏州金唯智生物科技有限公司测序。

表1 引物序列
荧光定量体系为20.0μL:染料SYBR Green PCR Master Mix 10.0μL,上下游引物各0.5μL,cDNA模板2.0μL,ddH2O 7.0μL。荧光定量在BIO-RAD iQ5荧光定量PCR仪上进行PCR扩增,反应程序为:95 ℃ 3 min;95 ℃ 10 s,55 ℃ 30 s,72 ℃ 30 s,40 个循环;72 ℃延伸步骤收集荧光信号,每个样品设置4个技术性重复,3个生物学重复。采用2-ΔΔCT相对定量分析方法计算出基因的相对表达量,并用DPS 7.05 数据处理系统软件进行方差及显著性分析。
1.2.4 CsMADS-box AGL62的生物信息学分析 通过 NCBI的 BLAST(https://blast.ncbi.nlm.nih.gov/Blast.cgi)对基因进行保守结构域的分析;利用MEGA 5.2对基因进行建树分析。利用ProtParam(http://web.expasy.org/protparam/)计算蛋白质的理论分子质量、理论等电点、原子组成、稳定系数和脂肪系数。利用蛋白质亚细胞定位工具PSORT(http://psort.hgc.jp/form2.html) 预 测 蛋 白 质 的 定位。通过SignalP 4.1(http://www.cbs.dtu.dk/services/SignalP/)及 IPSORT(http://ipsort.hgc.jp/)进行蛋白质信号肽预测。利用TMHMM(http://www.cbs.dtu.dk/services/TMHMM/)对蛋白质进行跨膜区域预测。通过 ProtScale(http://web.expasy.org/protscale/)分析蛋白质的亲水性与疏水性。用SOPMA(http://sopma.expasy.org/)及 Phyre 2(http://sbg.bio.ic.ac.uk/phyre2/)对蛋白质的二级结构、三级结构进行预测。
2 结果与分析
2.1 田间调查结果
亲本及F1调查结果见表2。从表中可以看出,3个时期的雌花数、雌花节率显示了F1能够很好地遗传亲本性状,如D1158♀-2、D1104-2-4为雌性系,其杂种一代东农812和C15-114也为雌性系,并且C15-114的雌性表现比母本D1104-2-4还要突出(表2)。D0528-2为雌雄异花同株,杂种一代C17-31也为雌雄异花同株。
根据杂种优势指数的计算方法:

其中F1表示杂种一代,P1、P2表示亲本,Hp表示亲本中较优良的亲本,Mp表示双亲的中亲值,OPH表示超亲优势,MPH表示超中优势。
通过和非雌性系父本杂交,以强雌系D1158♀-2为母本的杂种一代东农812的雌花数和雌花节率均低于母本,这在3个时期表现一致(图1),但由全雌系D1104-2-4作母本的C15-114在雌花数和雌花节率上产生了超亲优势(图1-A),表明利用全雌系可以获得在雌性上具有杂种优势的新品种。但是通过杂交笔者并没有获得具有早花超亲优势性状的杂种一代(图1-A),不过通过和具有早花性状的亲本杂交,F1能够很好地综合亲本的优良性状兼具早熟和雌性的特点(图1-B)。

表2 3个组合田间性状表现
2.2 黄瓜MADS-box基因家族测序结果分析
根据田间调查结果,以D1104-2-4为母本,D0708-2为父本配制的C15-114组合在雌花数、雌花节率上具有超亲优势,这一特点有利于研究黄瓜MADS-box基因家族在雌性杂种优势形成中的作用,因此挑选该组合进行转录组测序。
测序结果显示(表3),黄瓜37个MADS-box家族基因中大部分基因在父母本及子代中无差异表达。C15-114和母本D1104-2-4之间MADS-box家族基因差异倍数在-3.20~7.04,其中31个基因无差异表达,只有6个有差异表达,其中1个基因差异显著(差异倍数>4)。而C15-114和父本D0708-2之间MADS-box家族基因差异倍数在-5.83~5.55,其中有17个基因差异表达,20个无差异表达(表3)。根据C15-114、D1104-2-4及D0708-2的性状表现可知,C15-114、D11104-2-4属于全雌系而D0708-2属于雌雄异花同株,转录组的测序结果进一步体现了杂种一代的雌性偏向。

图1 3个F1的超亲及超中优势
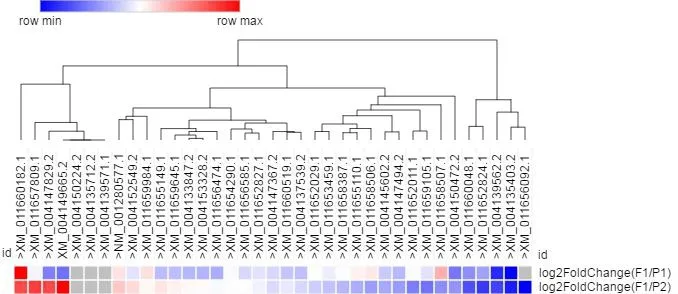
图2 黄瓜MADS-box家族基因转录组测序热图聚类分析
对黄瓜MADS-box家族基因进行聚类分析发现,差异表达的基因富集在相似的类别中,上调差异表达的基因和下调差异表达的基因分别聚集在2组不同的分支中(图2)。这表明杂种一代与亲本雌性产生差异的结果是由于部分基因差异表达。为了进一步分析差异基因特点,对黄瓜37个MADS-box家族基因构建系统发育树并通过比对到黄瓜基因组数据库(http://www.icugi.org/)分析各个基因的外显子分布(图3)。这些MADS-box家族基因的2种类型主要集中在2个区域(图4)。说明黄瓜中该家族基因进化上具有很大的保守性。通过转录组测序得到的差异较大且在2个亲本中差异表达相反的2个基因ID:101215235和ID:101206073均属于Type Ι( 图 3、4)。ID:101215235有 1个外显子区域,ID:101206073有2个外显子区域,二者都属于AGL62型。有研究表明,AGL62的基因功能与AGL61功能类似,AGL61可以与AGL80协作,共同参与雌配子体内中心细胞的分化(Bemer et al.,2008),并且AGL62同时还参与胚乳细胞的形成(Kang et al.,2008),这些都与植物的雌性分化密切相关。综上分析表明:引起杂交种C15-114与亲本间雌性差异及花期差异的原因很可能是这2个基因差异表达的结果。
为验证这一结论的准确性,挑选ID:101215235基因在3个组合中进行荧光定量验证分析(图5)。将每个组合的杂种一代作为参照,C15-114组合中父母本与子代均有差异,母本表现为近5倍的差异表达量,父本表现为近3倍的差异表达量,荧光定量结果和转录组测序结果基本一致(表3,图5)。分析发现2个雌性系组合父母本与杂种一代均有显著差异表达,而非雌性系C17-31组合中父母本与杂种一代差异不显著。表明基因ID:101215235在杂交组合中的差异表达可能参与黄瓜的雌性分化。
2.3 CsMADS-box AGL62基因克隆及分析
从黄瓜茎尖提取总RNA后,用琼脂糖凝胶电泳检测(图6-A)。结果显示RNA提取良好。进行PCR克隆(图6-B)后在745 bp左右出现条带,回收后与pGEM-T克隆载体相连转化到Trans1-T1感受态细胞中,挑白色菌落摇菌后测序。测序结果与NCBI上基因ID:101215235序列一致,表明克隆成功。
Protparam分析结果显示基因ID:101215235编码的蛋白含有187个氨基酸;理论等电点为9.18;预测的分子量为21.583 kDa;分子式为C970H1541N267O278S6;不稳定指数为54.96,属于不稳定蛋白(>40属于不稳定);脂肪族指数为84.49。NCBI比对结果表明该蛋白为MADS-box家族蛋白,类别为AGL62(图7),并且具有SRF-TF结构域,所以基因ID:101215235还是1个转录因子,把该基因命名为CsMADS-box AGL62。PSORT预测结果显示该蛋白可能为线粒体蛋白。
信号肽预测结果表明该蛋白没有明显的信号肽序列(图8-A、B),而TMHMM跨膜区域预测发现该蛋白在第30~50个氨基酸位置有跨膜,表明该蛋白是膜蛋白,且前30个氨基酸位于膜内,第50个氨基酸之后位于膜外(图8-C)。

表3 黄瓜MADS-box基因转录组测序结果

图3 黄瓜MADS-box家族基因发育树及外显子分布

图4 黄瓜MADS-box家族基因分类

图5 ID:101215235基因在3个F1组合中的荧光定量结果
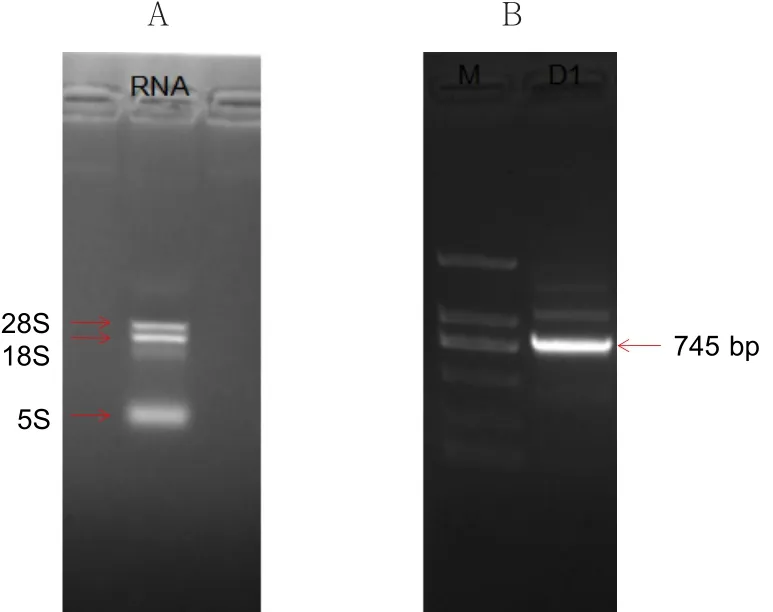
图6 RNA提取及PCR电泳

图7 CsMADS-box AGL62基因保守结构域

图8 CsMADS-box AGL62蛋白信号肽及跨膜区域预测
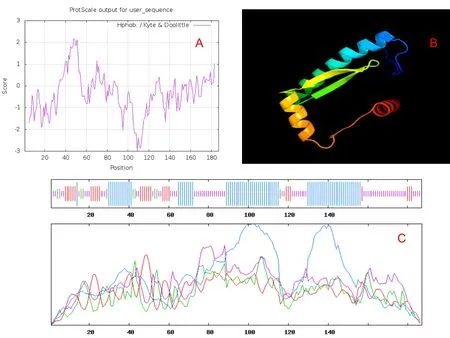
图9 CsMADS-box AGL62的疏水性及高级结构预测

图10 CsMADS-box AGL62系统进化分析
对蛋白质的疏水性预测发现,主要的疏水位点为第30~50个氨基酸(图9-A),该位点与跨膜预测结果中的跨膜位点一致。结合跨膜预测结果发现,氨基酸的膜外和膜内区域均为亲水性氨基酸,这些结果均与跨膜蛋白特征相似。蛋白质的二级结构分析表明,该蛋白共有76个α螺旋,占整个多肽链的40.64%;28个延伸主链占整个多肽链的14.97%;10个β折叠占整个多肽链的5.35%;73个无规则卷曲占整个多肽链的39.04%(图9-C)。用Phyre2同源建模预测蛋白的三级结构如图9-B所示。
不同物种的AGL62亚型基因系统发育分析发现黄瓜与甜瓜的同源性最高(图10-A),不同物种不同亚型的MADS-box家族基因系统发育分析表明,同一个物种不同亚型的MADS-box家族基因进化程度更为相近(图10-B)。结合图3和图4分析,CsMADS-box AGL62基因同一物种中相同的亚型具有最高的同源性,表明该基因在黄瓜进化中保守性很高。
3 讨论
本试验通过田间调查及转录组分析筛选出杂交种中与黄瓜雌花发育有关的差异表达基因,分析了MADS-box家族基因在黄瓜杂交种中的表达模式,并挑选1个该家族显著差异的基因ID:101215235进行生物信息学分析,并命名为:CsMADS-box AGL62。
MADS-box家族在植物中参与性别决定与花发育的机理已经有了深入的研究(Lin et al.,2015;Dreni & Zhang,2016;Zeng et al.,2018),但是该家族对植物雌性形成的杂种优势研究还很少。本试验发现MADS-box家族基因在杂交组合中表达模式显著偏向母本,这可能是F1的雌性相关农艺性状与母本相似的原因。有研究发现MADS-box家族基因参与植物早花的形成(de Oliveira et al.,2014),在本试验中发现所有的组合开花时间均介于母本与父本之间,并且更偏向父本,与雌花分化等性状偏向母本相反。MADS-box家族基因在杂交种中的表达调控机理还有待进一步研究。
AGL62基因属于MADS-box家族中I型基因,本试验在黄瓜中获得1个CsMADS-box AGL62基因。有研究表明,该基因调控胚乳的发育及细胞形成(Hehenberger et al.,2012),生物信息学分析表明,黄瓜中该基因位于线粒体膜上,属于转录因子,这说明CsMADS-box AGL62基因可能在黄瓜中调控线粒体基因的表达。
