Anourosorex squamipes染色体次缢痕多态性研究
2018-11-07李凤君徐含峰王旭明韦海雪陈顺德
李凤君, 徐含峰, 王旭明, 韦海雪, 张 琪, 王 琼, 陈顺德*
(1.四川师范大学生命科学学院,四川成都610101; 2.广西大学生命科学与技术学院,广西南宁530004)
染色体是遗传信息的载体,其次缢痕是一对同源染色体除着丝粒(主缢痕)外的另一个收缢部分.国内外对染色体次缢痕进行了相关研究[1-4],一些研究证实染色体次缢痕存在多态性[5-6].King[5]对澳大利亚12种雨蛙进行了带型分析,并对雨滨蛙属(Litoria)中出现的多态次缢痕进行了分类.刘万国等[6]也对4种蛙属两栖动物的次缢痕多态性进行了研究,并对次缢痕、银染核仁组织区(Ag-NORs)和结构异染色质间的相互关系进行了分析.
四川短尾鼩(Anourosorex squamipes)在分类学上属于鼩形目(Soricomorpha)鼩鼱科(Soricidae),栖息于海拔300~2 500 m的农田、灌丛等多种环境,地下及地面生活,杂食性[7],主要分布在四川、重庆、陕西、甘肃、湖北、贵州和云南[8].台湾短尾鼩(Anourosorex yamashinai)曾作为四川短尾鼩的亚种[9],Motokawa 等[10]依据染色体的不同将台湾短尾鼩定为独立种,同时报道了台湾短尾鼩的染色体次缢痕多态性.作为台湾短尾鼩的近缘种,四川短尾鼩的染色体次缢痕是否具有多态性尚未见报道.本文旨在研究四川短尾鼩的染色体核型,分析其染色体次缢痕是否存在多态性,为进一步研究染色体次缢痕多态性发生的原因及其功能提供基础资料.
1 材料与方法
1.1 实验材料 本研究于2016年在四川省成都市四川师范大学校园内,采用笼捕的方法共捕捉到成年四川短尾鼩12只,其中雄性9只,雌性3只,体重21 ~35 g,平均体重 30.3 g.
1.2 实验方法
1.2.1 染色体制备 采用骨髓细胞制片,参考文献[11-12]骨髓细胞染色体制备的方法并稍作修改.腹腔注射质量分数0.1%秋水仙素溶液(按照4μg/g体重注射),经过3~4 h处理后,采用颈椎脱臼法处死四川短尾鼩.通过取骨髓、低渗处理、固定、空气干燥法制片及染色后,在显微镜油镜下进行染色体的照相和记数分析.
1.2.2 染色体核型与次缢痕分析 观察分裂相好的中期细胞以及对染色体形态正常且分散良好的10个中期分裂相细胞进行计数、测量、显微摄像等.计算染色体的相对长度、臂比和着丝粒指数等的平均值和标准差,按文献[13]标准划分染色体形态类型,并编号排成核型图,将放大的染色体照片逐条裁剪、分类,并根据染色体形态特征,按照染色体长度递减顺序(X、Y染色体除外)配对排序.排序计算公式参考文献[14].选取77个形态清晰、分散较好的细胞(2n=48)对染色体次缢痕进行观察并计数,以分析四川短尾鼩次缢痕多态性.
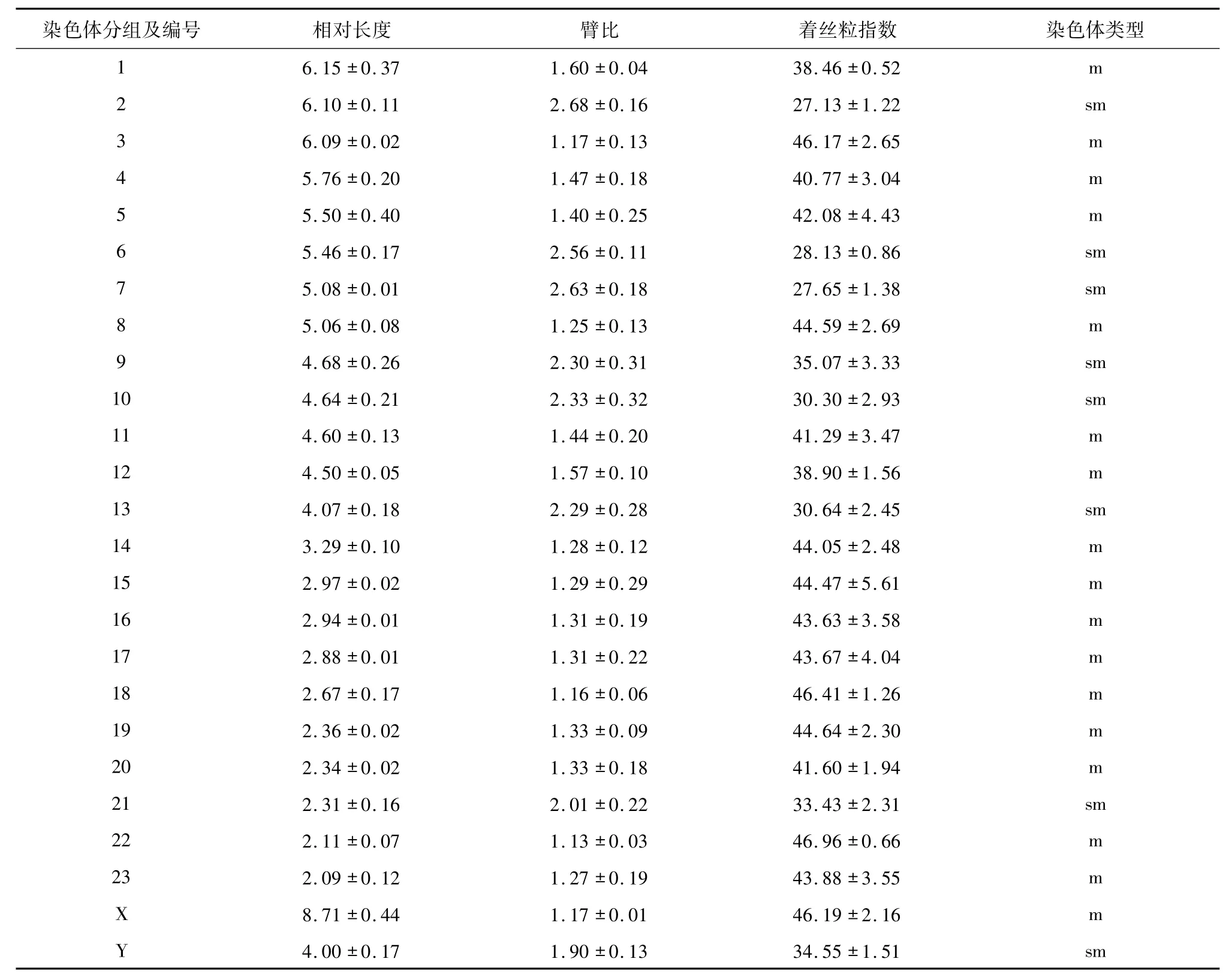
表1 四川短尾鼩染色体类型及测量Tab.1 Measurement and types of karyotype of Anourosorex squamipes
2 结果
2.1 染色体数目和染色体核型 通过对77个分裂相好的中期细胞的观察和对10个染色体细胞的拍照测量,结果表明四川短尾鼩染色体为二倍体,染色体数为2n=48.10个形态清晰、分散较好的细胞的染色体相对长度、臂比、着丝粒指数的平均值和方差分别见表1.按文献[13]的染色体形态及命名法则,四川短尾鼩23对常染色体中有16对染色体臂比在1.0~1.7,为中部着丝粒染色体,其余7对臂比皆在1.7~3.0,为亚中部着丝粒染色体,无亚端着丝粒染色体和端着丝粒染色体.在性染色体对中,X染色体为中部着丝粒染色体,为最大的染色体;Y染色体为亚中部着丝粒染色体,其相对长度介于第13号和第14号染色体之间,在所有的亚中部着丝粒染色体中,Y染色体相对长度仅大于第21号染色体.从相对长度上看,四川短尾鼩的最大染色体(X)是最小染色体(No.23)的4倍左右.根据以上分析,四川短尾鼩的染色体核型公式为
2.2 染色体次缢痕 在24对同源染色体(包括X、Y染色体)中,任何一对非同源染色体在长度、形态等方面存在着明显差异.在所有染色体对中,第21号、第22号染色体在形态结构上与其它染色体存在着非常显著的差异,在其染色体臂上存在独特的二级收缩结构(次缢痕),且染色体次缢痕数量在1~3条之间波动(表2和图1).在77个细胞(2n=48)中,10个细胞有1个次缢痕,50个细胞具有2个次缢痕,17个细胞具有3个次缢痕(表2).

表2 四川短尾鼩染色体次缢痕数量分布Tab.2 Distribution of secondary constriction structure numbers in Anourosorex squamipes
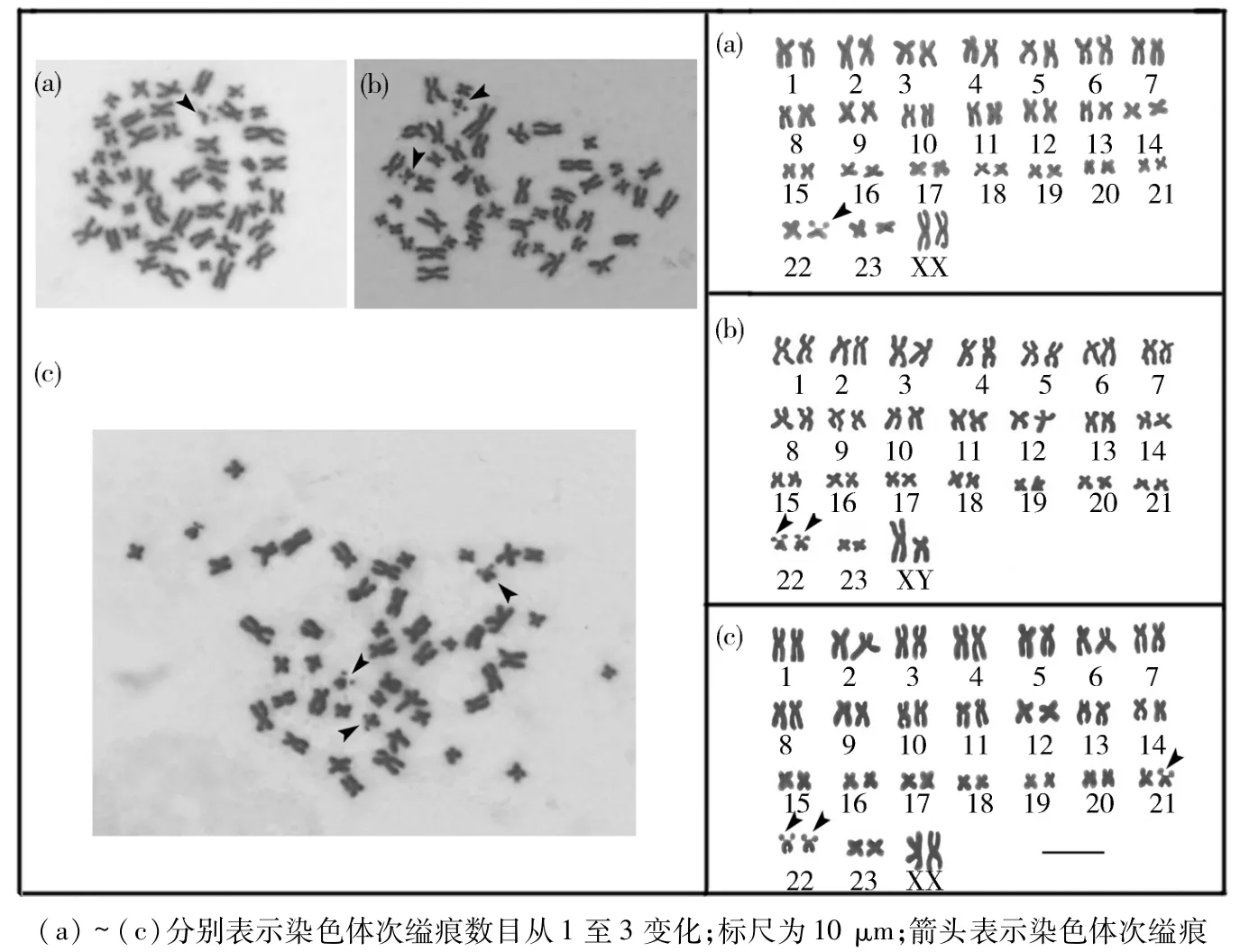
图1 四川短尾鼩染色体组型(2n=48)Fig.1 Conventional karyotype of Anourosorex squamipes(2n =48)
3 讨论
染色体数目与结构变异是物种分化和新物种产生的重要原因.四川短尾鼩的染色体组型(2n=48)、次缢痕位置(No.21、No.22)、次缢痕数量多态性(1~3条)与 Motokawa等[10]报道的台湾短尾鼩不同,后者2n=50,次缢痕位于第20号染色体上,数量1~2条.这些差异说明四川短尾鼩与台湾短尾鼩在长时间的物种进化过程中,由于大陆和台湾之间海峡的地理隔离,导致2个物种间形成了不同的染色体和次缢痕.在鼩鼱科中,物种内具有染色体次缢痕数量多态性的物种仅有3种,除四川短尾鼩和台湾短尾鼩外,还有日本水鼩鼱(Chimarrogale platycephalus)[15].次缢痕数量多态属于次缢痕多态性的一种,对于物种内染色体次缢痕数量多态产生的原因尚不清楚,还有待进一步研究.
次缢痕是染色体上的一个缢缩部位,其数量、位置和大小是染色体的重要形态特征,一般认为染色体的次缢痕区域与核仁组织区的形成有关.在次缢痕多态的遗传机制的研究中,陈广文等[16]认为次缢痕多态和染色体数目的变化是物种进化的重要因素之一.King[5]对12种两栖动物次缢痕多态性进行了研究,认为次缢痕位置的改变来源于潜在的核仁组织区的扩大.刘万国等[6]探讨染色体次缢痕(SC)、核仁组织区(NORs)和结构异染色质三者间的关系,认为这三者之间的关系是错综复杂的,应视物种的不同而定.本文从细胞遗传学的角度证实了相同地区的四川短尾鼩染色体次缢痕数量多态性的现象,研究结果将对深入研究染色体次缢痕多态性的发生原因及其遗传机制具有重要意义.
