甲状旁腺素相关肽蛋白核定位序列及C末端缺失对小鼠下颌磨牙发育的影响∗∗
2018-10-31颜成东蒋尚飞
刘 洪,颜成东,蒋尚飞
(江苏医药职业学院医学技术学院,江苏盐城 224005)
甲状旁腺素相关肽(parathyroid hormone related peptide,PTHrP)的结构和作用均和甲状旁腺素(PTH)相类似[1]。PTHrP和PTH有相同的受体,并且PTHrP通过自分泌/旁分泌方式发挥多种生理功能[2]。PTHrP的N-末端(1~36)和PTH的功能基本相同;而PTHrP核定位序列(87~106)和C-末端共同通过细胞内分泌的方式达到促进细胞增殖的功能[3]。为了研究PTHrP核定位序列及C末端的功能,利用基因工程在PTHrP的第84位氨基酸序列后敲入一个终止密码TGA,制备了不表达PTHrP核定位序列及C-末端的小鼠模型,称为PTHrP Knock-in(PTHrP KI)小鼠。这种小鼠的表型有生存时间变短,以及骨骼、脑组织和皮肤发育不良等表现[4]。为了研究PTHrP核定位序列及C末端缺失对小鼠下颌磨牙发育是否存在影响,本研究利用出生后2周的PTHrP KI小鼠和野生型小鼠的下颌磨牙进行比较,研究PTHrP核定位序列及C末端缺失对小鼠下颌磨牙发育和矿化的影响。
1 材料与方法
1.1实验动物 使用雄性PTHrP KI杂合子小鼠和雌性PTHrP KI杂合子小鼠交配(小鼠购自江苏省实验动物中心),获得同窝的野生型(wild type,WT)小鼠和PTHrP KI纯合子(PTHrP KI)仔鼠。仔鼠由母鼠喂养,母鼠使用普通饲料进行喂养。待小鼠生长至2周后取材,WT小鼠(WT组)和PTHrP KI小鼠(PTHrP KI组)分别取10只进行后续的影像学、组织学和分子生物学实验。本次实验经江苏医药职业学院伦理委员会审核通过。
1.2 方法
1.2.1动物基因型鉴定 小鼠出生后剪取鼠尾,使用蛋白酶K进行消化,提取DNA后利用PCR进行目标片段扩增反应,产物使用BstEⅡ内切酶进行酶切反应后行琼脂糖凝胶电泳,WT小鼠条带为424 bp,PTHrP KI小鼠条带为258 bp和166 bp[5]。
1.2.2取材 WT小鼠和PTHrP KI小鼠生长至2周后,颈椎脱臼处死,解剖分离左右两侧下颌骨,使用免疫电镜固定液固定后行CT扫描,使用乙二胺四乙酸(EDTA)脱钙后石蜡包埋,切片。
1.2.3影像学检查 使用SkyScan 1072 scanner CT扫描仪对WT小鼠和PTHrP KI小鼠下颌骨进行扫描,通过CT检查观察,沿着下颌第一磨牙近中根管方向进行三维重建。
1.2.4苏木精-伊红(HE)染色 石蜡切片分别使用100%、75%和50%乙醇进行脱蜡,苏木精染色8 min,伊红染色2 min,分别使用50%、75%、100%乙醇脱水,二甲苯透明,中性树胶封片。在光学显微镜下观察放大100倍HE染色切片,测量两种小鼠下颌第一磨牙长度,放大400倍HE染色切片测量两种小鼠的下颌第一磨牙前期牙本质占总牙本质的比例。
1.2.5免疫组织化学检查 石蜡切片使用牛睾丸透明质酸酶消化,Ⅰ型胶原(Collagen-Ⅰ)一抗为免抗鼠多克隆抗体(美国Santa Cruiz公司生产),二抗为生物素标记的羊抗兔二抗(美国Santa Cruiz公司生产),使用生物素-亲和素-桥联碱性磷酸酶酶标法(ABC-AP,美国Vector公司生产)孵育,碱性磷酸酶染色液染色后使用甲基绿复染。牙本质涎蛋白(dentin sialoprotein,DSP)、Ki-67和 p27一抗均为兔抗多克隆抗体(美国Santa Cruiz公司生产),二抗为辣根过氧化物酶标记的羊抗兔二抗(美国Santa Cruiz公司生产),链霉菌抗生物素蛋白-过氧化物酶连结法(Elite-AP,美国Vector公司生产)孵育后使用二氨基联苯胺(DAB)染色液染色,DSP使用苏木精复染。

2 结 果
2.1影像学检查 Micro-CT扫描三维重建结果显示,PTHrP KI组小鼠的下颌第一磨牙牙根长度明显小于WT组小鼠,见图1。
2.2HE染色 WT组小鼠的下颌第一磨牙长度较PTHrP KI组小鼠长,差异有统计学意义(t=6.109,P<0.01);WT组小鼠的前期牙本质占总牙本质比例小于PTHrP KI组小鼠,差异有统计学意义(t=9.178,P<0.01),见图2、表1。
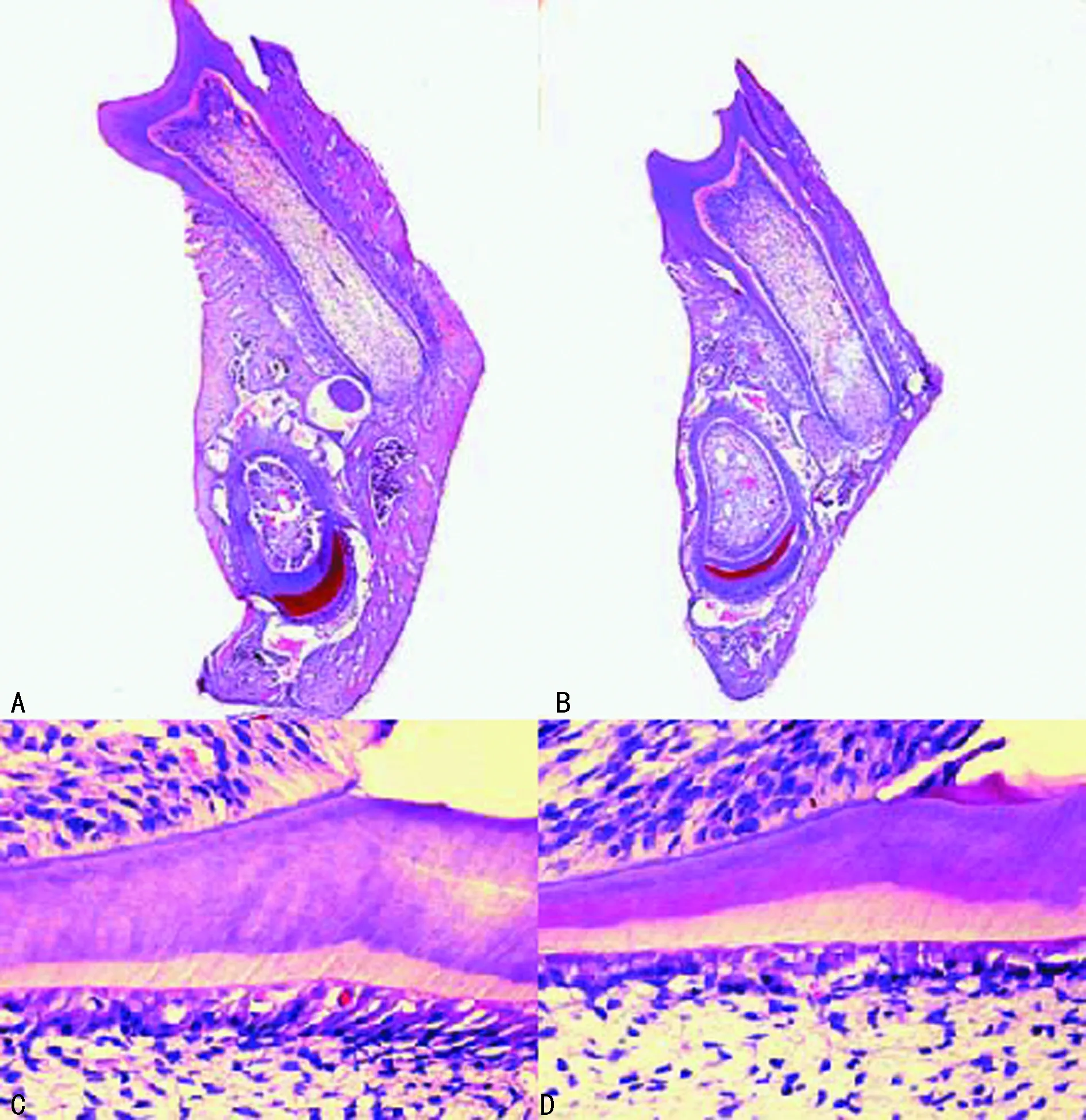
A:WT组小鼠下颌骨(×100);B:PTHrP KI组小鼠下颌骨(×100);C:WT组小鼠下颌第一磨牙牙颈部(×400);D:PTHrP KI组小鼠下颌第一磨牙牙颈部(×400)图2 两组小鼠HE染色
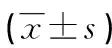
表1 两组小鼠下颌第一磨牙长度和前期牙本质占总牙本质比例比较
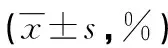
表2 两组小鼠下颌第一磨牙Collagen-Ⅰ、DSP、Ki-67和p27阳性表达比较
2.3免疫组织化学染色 WT组小鼠Collagen-Ⅰ占下颌第一磨牙阳性比例、Ki-67阳性细胞比例和DSP占牙本质阳性比例均高于PTHrP KI组小鼠,p27阳性细胞比例低于PTHrP KI组小鼠,差异均有统计学意义(P<0.05),见图3、表2。
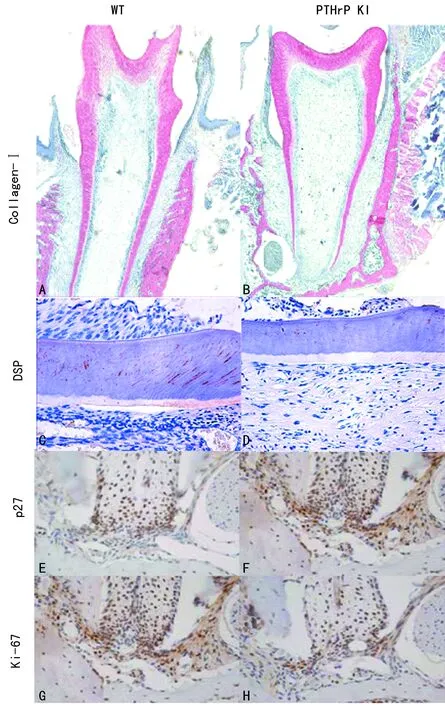
A、B:Collagen-Ⅰ(×100);C、D:DSP(×400);E、F:p27(×400);G、H:Ki-67(×400)图3 两组小鼠磨牙中Collagen-I、DSP、Ki-67和p27阳性表达
3 讨 论
在牙齿的发育、萌出和矿化过程中,多种钙、磷调节激素如1,25(OH)2D3[6]、成纤维细胞生长因子-23(fibroblast growth factor-23,FGF-23)[7]的缺乏或者过表达都有可能干扰牙齿细胞外基质的分泌和矿化,最终导致牙齿发育异常。
本研究结果显示,CT扫描后重建结果表明与WT小鼠比较,PTHrP KI小鼠下颌第一磨牙的长度缩短;通过对下颌第一磨牙进行HE染色、分析,结果发现PTHrP KI小鼠下颌第一磨牙的长度明显缩短;进一步分析表明,PTHrP KI小鼠下颌第一磨牙前期牙本质的比例明显高于WT小鼠。在牙本质的发育阶段,成牙本质细胞分泌细胞外基质形成前期牙本质,前期牙本质在不断矿化之后形成牙本质,如果前期牙本质的矿化过程受阻,则前期牙本质占总牙本质的比例就会明显升高[6]。本研究HE染色结果表明,小鼠PTHrP核定位序列和C末端缺失可能导致了牙本质基质生成减少和矿化的异常。
应用免疫组织化学法进行分析,结果发现,PTHrP KI小鼠的Collagen-Ⅰ在磨牙区的阳性表达比例明显小于WT小鼠。Collagen-Ⅰ是成牙本质细胞分泌的细胞外基质的主要成分,并且在牙本质矿化过程中能够作为羟基磷灰石沉积的框架[8]。研究结果表明,小鼠PTHrP核定位序列和C-末端缺失导致了小鼠磨牙细胞外基质形成减少。PTHrP KI小鼠下颌第一磨牙牙本质中DSP的阳性面积明显小于WT小鼠,DSP是由牙本质涎磷蛋白(dentin sialophosphoprotein,DSPP)基因转录后进行编辑合成,不仅能够促进成牙本质细胞的分化,而且还能够在牙本质的矿化过程中调节羟基磷灰石晶体的形成,而且能够促进前期牙本质形成牙本质,因此,DSP也被认为是牙本质矿化的重要指标[9]。本研究结果表明,PTHrP核定位序列和C末端缺失导致了DSP合成减少,进而阻止了前期牙本质的矿化过程。免疫组织化学结果发现,PTHrP KI小鼠下颌第一磨牙牙髓和上皮根鞘处p27的阳性细胞比例明显增加,而Ki-67的阳性细胞比例则有所降低。在有丝分裂期,Ki-67表达水平最高,因此可以把Ki-67作为细胞增殖状态重要细胞因子[10-11]。p27是细胞周期蛋白依赖性激酶抑制因子(cyclin dependent kinase inhibitors,CDKI)家族成员,CDKI能够和细胞周期蛋白-细胞周期蛋白依赖性激酶(CDK)复合物结合,抑制CDK的活性,从而阻断细胞的增殖过程[12]。而p27是PTHrP的下游靶标,当PTHrP KI小鼠体内表达PTHrP蛋白中核定位序列和C-末端缺失,导致残缺的PTHrP蛋白不能进入细胞核,丧失了对p27的调控抑制作用,从而导致了p27表达水平上调[13]。
PTHrP发挥促进细胞增殖、抑制细胞凋亡的作用是通过其蛋白的NLS和C-末端以细胞内分泌的方式进入细胞核后实现的,而PTHrP KI小鼠体内的PTHrP蛋白缺失了核定位序列和C-末端。体外研究表明,PTHrP缺失核定位序列和C-末端片段能调控细胞周期相关细胞因子的表达,从而使得细胞凋亡明显增加[14]。研究发现,PTHrP在牙齿间充质细胞、牙乳头、上皮根鞘等处均有表达[15],而PTHrP KI小鼠分泌的PTHrP蛋白因为缺失核定位序列和C-末端,失去了通过细胞内分泌方式进入细胞核来发挥促细胞增殖的作用,导致牙髓p27蛋白表达水平上调,Ki-67蛋白表达水平下降,导致牙髓间充质细胞的凋亡增加和向成牙本质细胞的分化减少,进而导致成牙本质细胞胞外基质分泌减少,并且进一步影响了牙本质的生成和矿化;此外,牙根的形成和上皮根鞘密切相关[16-17],同样,PTHrP蛋白核定位序列和C-末端缺失还可能导致了小鼠磨牙上皮根鞘处的细胞凋亡增加,进而导致了PTHrP KI小鼠磨牙牙根发育障碍。
综上所述,PTHrP核定位序列和C-末端缺失导致小鼠磨牙细胞增殖受阻,牙本质细胞外基质分泌减少,矿化障碍,最终导致了小鼠磨牙的发育异常。
