药用植物四环三萜皂苷生物合成及代谢研究进展*
2018-10-30孟珍贵龙光强李龙根张广辉杨生超陈军文
杨 玲,范 伟,孟珍贵,龙光强,李龙根,张广辉,杨生超,陈军文**
(1.云南农业大学农学与生物技术学院 昆明 650201;2.云南农业大学云南省药用植物生物学重点实验室 昆明 650201;3.云南农业大学西南中药材种质创新与利用国家地方联合工程研究中心 昆明 650201)
三萜皂苷是以糖苷键将三萜皂苷元和糖结合从而形成的配糖体,其中包括四环三萜(达玛烷型为主)和五环三萜(齐墩果烷型为主)两大类,图1所示分别为达玛烷和齐墩果烷以及各自代表性的三萜皂苷:20(S)-原人参二醇和黄甘草皂苷。近几十年还发现了由于氧化、环裂解、甲基转位、重排及降解等产生的高度氧化的新骨架类型三萜皂苷。三萜皂苷结构复杂,从植物中直接提取或通过化学合成间接获得都较为困难[1]。因此,阐明四环三萜皂苷的生物合成途径及代谢调控对于了解三萜皂苷的整体代谢框架具有重要意义。
四环三萜皂苷(Tetracyclic triterpenoid saponins)作为三萜皂苷中重要的一类,其主要包括达玛烷型(Dammaranes)、羊毛脂烷型(Lanostanes)、环菠萝蜜烷/环阿屯烷型(Cycloartanes)、大戟烷型(Euphane)、葫芦烷型(Cucurbitanes)、楝烷型(Meliacanes)、甘遂烷型(Tirucallanes)等。目前四环三萜皂苷主要存在于双子叶植物中,灵芝属中也有大量报道发现,如许多常用药用植物如人参(Panax quinquefolius)[2]、黄芪(Astragalus membranaceus)[3,4]、三七(Panax pseudoginseng)[5]、麦 冬(Ophiopogon japonicus)[6,7]、升麻(Cimicifuga foetida)[8,9]、灵芝(Ganoderma lucidum)[10,11]等都含有四环三萜皂苷类成分。
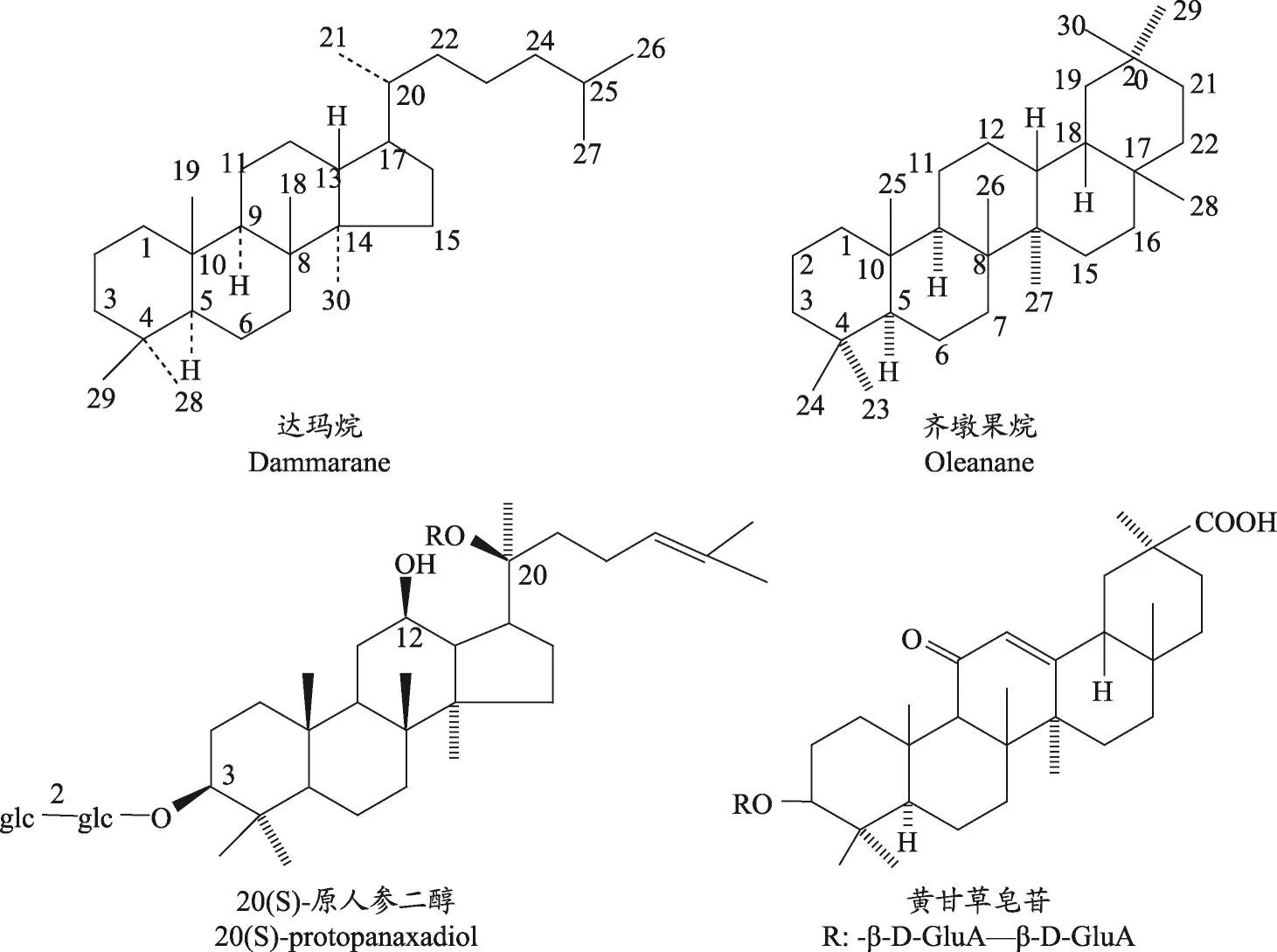
图1 三萜皂苷中达玛烷与齐墩果烷型皂苷结构示意图
虽然很多药用植物的有效成分属于四环三萜皂苷化合物,但在药用植物中其往往含量偏低,天然药用植物资源十分有限,严重影响了其活性成分的开发利用。药用植物资源匮乏及其种植所面临的一系列问题推进了基因工程等技术在药用植物领域的广泛利用。目前三萜皂苷代谢工程研究仍处于前期初步探索过程中,对大多数四环三萜皂苷的合成途径和代谢调控机制尚未进行深入研究,相关酶基因及其表达调控的研究报道还比较少,而通过代谢工程或基因工程进行四环三萜皂苷生物合成的工厂化生产,具有广阔的市场前景。本文综述了药用植物四环三萜皂苷的一些代表性成果,探究其在次生代谢工程及基因工程领域内的研究方法与策略,以期对药用植物四环三萜皂苷后续研究提供一定的参考价值。
1 四环三萜皂苷及其生物合成
目前研究发现,四环三萜皂苷的代谢合成主要通过甲羟戊酸途径(Mevalonate pathway,MVA途径)和类异戊二烯途径(Isoprenoid Biosynthetic Pathway,MEP途径),形成异戊烯焦磷酸(IPP)和二甲烯丙基焦磷酸(DMAPP)前体物,生成2,3-氧化鲨烯(2,3-oxidosqua⁃lene),再经后续的一系列关键酶及相关酶,如达玛烯二醇合成酶(DS)、环阿屯合酶(CS)、羊毛甾醇合酶(LS)、细胞色素P450单加氧酶(CYP450)、糖基转移酶(UGT)等进行三萜骨架修饰,从而合成多种结构和功能各异的四环三萜皂苷[12,13]。
1.1 达玛烷型三萜皂苷
达玛烷型如五加科植物人参、三七和西洋参(Panax quinquefolius)等的根、茎、叶、花、果实中均含有多种人参皂苷[14-18],酸枣(Ziziphus jujuba)仁中的酸枣仁皂苷(jujuboside)A和B[19],以及绞股蓝中的绞股蓝(Gy⁃nostemma pentaphyllum)皂苷[20,21],其苷元绝大多数属于达玛烷型四环三萜,达玛烷型三萜皂苷结构从环氧鲨烯由全椅式构象形成达玛烷骨架,主要分为20(S)-原人参二醇和20(S)-原人参三醇。达玛烷型皂苷在心脑血管系统、抗衰老及抗肿瘤等方面具有广泛应用[22-25],迄今为止其研究仍关注在少数几种植物中,深入挖掘达玛烷型皂苷生物合成及代谢机理将有助于后续对达玛烷型皂苷的开发利用。
1.2 环阿屯烷型三萜皂苷
环阿屯烷型基本骨架在C19位甲基与C9位脱氢形成三元环,其母核结构在C13,C14位分别连接有β、α-CH3,环阿屯烷型三萜皂苷在豆科、五味子科、毛茛科、大戟科、楝科、仙茅科等植物中均有报道其化学结构及药理活性的相关研究[26-29],如黄芪具有补气强壮之功效,从其中分离鉴定出20个左右的皂苷,多数皂苷元为环黄芪醇[30];环阿屯烷型三萜除在中药黄芪中发现外,在升麻族植物中广泛存在[31];白芨(Bletilla striata)鉴定出环阿屯型三萜并且研究了其抗血管生成活性[32]。
1.3 其他四环三萜皂苷

图2 人参皂苷Rb1和Rg1生物合成途径
羊毛脂烷亦称羊毛脂甾烷,与达玛烷型极为相似,不同点在于其C18甲基连在C13位,有研究表明在灵芝中分离出羊毛脂烷型三萜皂苷,是羊毛甾烷的高度氧化化合物[33-35]。此外,从葫芦科雪胆属雪胆(Hemsleya chinensis)中雪胆甲素、雪胆乙素均属于葫芦烷型[36-39];有研究发现无患子(Sapindus mukorossi)中含有大戟烷型三萜皂苷[40];楝烷型三萜皂苷主要在楝科植物中有所报道[41-43];而甘遂烷型三萜皂苷在植物分布中比较罕见,在臭椿属(Ailanthus Desf.)中有分离得到[44-47],从生源合成来看,甘遂烷型三萜等原柠檬苦素是形成四降三萜的前体物质[48]。目前羊毛脂烷型、葫芦烷型、大戟烷型、楝烷型、甘遂烷型三萜皂苷相关文献报道较少,大多数研究还停留在其化学成分的提取分离鉴定及生物活性分析方面,对于生物合成与代谢调控的研究尚浅。
1.4 四环三萜皂苷生物合成
在四环三萜皂苷生物合成过程中,值得注意的是2,3-氧化鲨烯经环氧角鲨烯环化酶(OSCs)的催化发生环化反应这一关键步骤[49-51]。不同四环三萜化合物其立体构型是复杂多样的,而作为多基因家族的OSCs,可使环氧角鲨烯发生环化,从而生成各类骨架的三萜化合物,然后通过细胞色素P450单加氧酶、糖基转移酶和糖苷酶一系列相关酶的作用等对骨架进行氧化置换和糖基化等化学修饰,最终生成不同类型的四环三萜皂苷类化合物。由于受到多种因素的调节,且在这一合成的动态变化过程中所涉及到的各种中间产物和酶具有复杂性,因此目前对四环三萜皂苷的合成机理认识尚不清晰。
以典型的四环三萜皂苷——人参皂苷Rb1和Rg1为例[52]由人参皂苷Rb1和Rg1生物合成途径(图2),可见以,2,3-氧化鲨烯为分支点,不同的OSCs催化2,3-氧化鲨烯,获得达玛烯二醇(Dammaranediol),再经过糖基转移酶的化学修饰,生成人参皂苷Rb1和Rg1[53-55]。全化学合成在人参皂苷生产中尚未得到实现,因此对那些含量少而活性强的人参皂苷单体的研究具有一定的难度。虽然目前对四环三萜皂苷生物合成途径的研究逐渐深入,但由于其中、下游合成途径中的多基因家族酶的结构及类型复杂多样,导致离其合成具体步骤的阐明还有一段距离。因此,若要将生物代谢工程及基因工程的方法应用到四环三萜皂苷的合成中,还需对四环三萜皂苷类化合物生物合成过程中在碳环骨架建立后所涉及的各种复杂的官能团反应进行深入研究。
2 皂苷的代谢调控
目前有大量文献报道关于五环三萜皂苷的合成及代谢调控,相对四环三萜皂苷研究而言,要更为深入[56]。根据四环三萜皂苷相关研究发现,在其代谢合成途径中,一方面与外界环境因素息息相关,这些因素包括激素、诱导子和环境刺激等[57-60],而这些作用具体是通过哪种途径实现的目前尚未研究清楚,需要进一步探索。另一方面关键酶基因从基因转录水平及转录后或翻译水平上对其进行调控[61],即通过基因工程调节合成路径中一些关键化合物的代谢流,利用调节基因和转录因子来调控关键酶编码基因的表达水平,或使关键酶编码基因过表达,都有可能有利于四环三萜皂苷类化合物的大量合成。在代谢途径中酶的调节作用是极为重要的,但由于目前路径研究尚未清楚,特别是受一些下游合成路径中涉及的复杂酶类的影响,导致四环三萜皂苷类化合物全合成研究成为一大难题。
2.1 四环三萜皂苷的代谢调控
2.1.1 关键酶基因的调控
根据已推断出的四环三萜皂苷类化合物生物合成路径[62],人们对其中部分关键酶及其基因进行了深入研究。参与四环三萜皂苷生物合成路径的关键酶,总体上可分为上游催化IPP和DMAPP合成的关键酶;中游催化IPP、DMAPP生成和萜类碳环骨架化合物以及中间体形成的关键酶;下游催化中间体和萜类化合物进行复杂的结构修饰从而获得不同四环三萜皂苷的关键酶。近年来,众多学者关注与研究的热点仍与四环三萜皂苷整个生物合成途径的阐明相关,特别是关于三萜碳环骨架合成及环上复杂官能团化等相关酶基因的鉴定与克隆,了解这些相关酶及其基因在合成途径中的作用对四环三萜皂苷类化合物的代谢调控及更好发挥其药用价值具有深远意义。
迄今为止,关于四环三萜皂苷类化合物的代谢过程中上游关键酶研究已经比较清楚,而中下游关键酶对代谢的影响尚未明确。有研究表明,过量表达SS基因,可促进刺五加(Eleutherococcus senticosus)中三萜皂苷类化合物的大量合成,表明SS基因在绞股蓝皂苷生物合成途径中具有关键性地位[63]。蒋军富等利用RTPCR和RACE技术,对绞股蓝SE基因进行了克隆及序列分析[64]。Han等发现PgSQE1的表达受阻将导致人参皂苷的产生减少,其参与调节人参皂苷的生物合成[65]。Liu等研究2年生人参中PgSS和PgSQE1基因表达水平最高,可间接调控皂苷合成,PgDS转录水平与人参皂苷Rg1,Re和Rf显著相关,PgSS基因表达与人参皂苷Rb1合成呈负相关[66]。Dhar等对催眠睡茄(Withania somnifera)进行分析确定了角鲨烯合酶、角鲨烯环氧化酶、环蒿醇合酶、细胞色素P450还原酶等在其不同发育阶段的代谢产物中的表达模式[67]。缺乏相关基因的克隆及分析,是了解其生物合成、代谢调控及限制生物技术手段干预合成的主要瓶颈之一。
氧化鲨烯环化酶(OSC)是调控代谢流进入四环三萜皂苷途径的重要分支位点,而DS是OSC家族成员之一,是合成达玛烷型人参皂苷的关键靶酶,也是导致三萜产物多样性的一个重要原因。Luo在研究中表明DS在四年生三七根部高表达,并获得了参与三萜皂苷合成的候选基因,包括CYP450s和UGTs[68]。此外,在这些关键酶当中,值得注意的是P450s和GTs这两类在植物代谢过程中占据重要地位的多基因家族,其中P450s可以利用氧气和NAD(P)H催化三萜骨架羟基化,使其实现区域专一性及立体专一性,是四环三萜皂苷生物合成路径中的关键酶[69-73]。Han等在报道中提到,原人参二醇合成酶CYP716A47属于一个细胞色素P450单加氧酶,其参与催化达玛烯二醇II羟基化在C12位产生原人参二醇[74]。目前,对CYP450的晶体结构以及相应的蛋白结合位点并未给出明确的说明,导致有关四环三萜皂苷生物合成途径的下游修饰机制未能完全阐明。
2.1.2 其他调控因素
研究表明,某些激素,如茉莉酸甲酯(MeJA)等,对四环三萜皂苷合成代谢中关键酶基因的表达具有调节作用[75]。在人参不定根组织培养中,培养基中施加一定浓度的MeJA后,促进了人参SS基因的表达[76]。Choi等研究表明SE等三个基因与三萜皂苷合成高度相关,而MeJA通过诱导SE基因高表达从而提高三萜皂苷合成量[77]。Oktae等利用MeJA处理人参不定根,发现PgSS、PgSE和DS基因表达水平改变从而对人参皂苷Rb1、Rg1含量造成不同影响[78]。关于激素对合成代谢的调控目前尚在研究初始阶段,大部分研究集中在少数几种激素,然而不同激素对四环三萜皂苷的生物合成影响需要更进一步研究,并且激素之间的互作也要引起大家的重视。
其他外界刺激如光照、土壤水分、金属离子、外源物质等对药用植物中四环三萜皂苷合成的影响也逐渐引起人们重视[79-81]。例如,阳生植物对光照的需求较高,光强较大时三萜皂苷的合成较为旺盛,如绞股蓝在相对光照度为70%左右时绞股蓝总苷量最高[82]。光反应产物是次生代谢的必需物质,因此光照强度影响植物体三萜皂苷生物合成机理极有可能与光反应产物相关。李佳洲等研究发现土壤水分含量为0.45W时有利于人参皂苷Rg1的积累,人参皂苷Rb1则在0.70W处理下积累最多[83],可见土壤不同水分含量下四环三萜皂苷积累有所差异。此外,有研究表明,金属镉(Cd)胁迫诱导内源NO的产生,而NO促进SE的表达,抑制DS的表达水平,即抑制了2,3-氧化鲨烯向达玛烷型三萜皂苷的代谢流[84]。胡向阳等利用壳聚糖处理从而提高人参皂苷合成中皂苷的积累,发现其SE转录水平提高[85]。由于研究尚未透彻,需要人们更进一步对于这些因素做出准确合理的分析。
四环三萜皂苷类化合物种类较多且合成途径长而复杂,这给控制其代谢流的遗传操作带来极大挑战。明确相关酶基因表达在组织和发育过程中的调控机制,对基因时空调控作用的各种顺式和反式作用因子进行研究,从而进一步补充和完善四环三萜皂苷类化合物生物合成途径及其网络,将为今后实现大规模生产四环三萜皂苷类化合物奠定基础。
3 结论与展望
近年来,药用植物皂苷在医药领域的应用越来越广泛,市场需求日益增高,这一现状导致如何提高药用植物中皂苷含量成为目前研究热点。在药用植物次生代谢产物生产中,制约四环三萜皂苷产量提高的原因是多方面的,但归根结底还是由于其生物合成途径和代谢机制尚未明确、基因工程手段还有待完善。通过对药用植物四环三萜皂苷合成途径的研究,进一步明确其代谢机制,从而推动药用植物皂苷代谢工程的发展,促进药材种植规范化、品种优良化、资源可持续化的发展。因此,未来可以从以下几个方面深入研究。
3.1 药用植物四环三萜皂苷类次生代谢物多样性
药用植物分布广泛,而四环三萜皂苷种类多且生物合成途径复杂,导致目前研究较为清楚的仍只是其中的很小一部分,仅鉴定和克隆了少数基因,对其代谢途径的整体调控以及次生代谢途径之间的协调作用等方面依旧存在很多盲区,而利用系统生物学方法开展预见性代谢工程将会拓宽未来的研究领域。
3.2 合成诱导机制多样性
一种四环三萜皂苷类化合物的合成可能受几种因素的诱导,一种因素又可诱导不同类型皂苷的合成积累,同一因素对不同药用植物的诱导作用也具有差异性,这给药用植物四环三萜皂苷在代谢调控机制方面的研究带来极大挑战,使得实际应用药用植物次生代谢调控生产皂苷类化合物增加了一定难度。探究药用植物次生代谢的具体路径及其调控网络,从不同侧面整合与分析代谢过程及其调控手段,提高各种调控之间的联合使用,在未来药用植物研究中具有一定指导意义。
3.3 基因工程手段局限性
某些药用植物繁殖周期长,在遗传育种上难度大,不同类型的四环三萜皂苷合成可能具有单独的合成途径和独特的酶促反应机制,导致基因工程手段的实施具有一定的盲目性和局限性,极大影响了研究工作的进程。未来研究中要运用各种代谢调控手段干扰代谢流,将四环三萜皂苷的化学成分研究与基因表达分析充分结合,不断将新兴技术融合应用到实验研究中。
虽然对很多药用植物四环三萜皂苷生物合成途径已有了一定的研究,但由于其合成中、下游途径中的多基因家族酶的结构及类型的复杂性,大多数合成具体步骤尚未得到清晰的阐明。随着对药用植物四环三萜皂苷生物合成关键酶基因调控研究的深入,阐明其生物合成途径中的关键酶基因及其协同性,利用人工手段调控关键酶基因表达水平,尝试经细胞培养、生物转化、发根培养等一系列工业化生产去实现大规模生产药用植物皂苷,这将是未来可能实现的一大突破。可以预见,随着对药用植物四环三萜皂苷合成代谢网络的深入全面解析和相关关键酶基因的克隆,以及现代生物技术手段的不断完善推进,药用植物四环三萜皂苷代谢工程将更加有效、更好地满足人类的需求。