不同种群密度地区长足大竹象生殖系统的超微结构研究
2018-10-29梁爽龙文聪梁梓罗朝兵刘安萱廖鸿杨瑶君
梁爽, 龙文聪, 梁梓, 罗朝兵, 刘安萱, 廖鸿, 杨瑶君
(乐山师范学院生命科学学院,竹类病虫防控与资源开发四川省重点实验室,四川乐山614004)
长足大竹象Cyrtotrachelusbuqueti是竹林毁灭性害虫,自2003年一直被列为我国林业危险性有害生物之一。据调查,四川省乐山市丛生竹林面积667 km2,长足大竹象平均虫口密度为0.49头/笋(杨瑶君等,2011),而贵州省赤水市丛生竹林面积820 km2,平均虫口密度低于0.01头/笋。两地相距不足300 km,虫口密度却相差48倍。生殖是维持昆虫种群密度的关键环节之一,生殖系统结构的变化可能导致昆虫繁殖能力或种群数量的显著变化。2014—2017年,本课题组在田间调查时观察到赤水地区长足大竹象雌虫产卵数量较少,其生殖器官的结构是否发生了显著变化?其变化特点如何?
近年对长足大竹象的研究主要集中在头部感器的超微结构(杨瑶君等,2010)、成虫体表信息化学物质的提取和鉴定(忙定泽,2012)、营养成分(蒲远凤等,2015)、繁殖性能(杨桦等,2015)与生物学特性及防治(聂学文,2010)等方面。在长足大竹象生殖系统方面,汪淑芳等(2009)对生殖结构形态解剖及梁梓等(2016)对生殖系统中雄性附腺的抑菌作用进行了研究。本文应用解剖及扫描电镜技术观察乐山高种群密度、赤水低种群密度长足大竹象生殖系统的形态特征和超微结构,比较二者的差异,以期分析赤水地区长足大竹象种群密度低的原因,为研究长足大竹象的种群特征和发生规律提供依据。
1 材料与方法
1.1 试验地概况及试验材料
贵州省赤水市大同镇位于市境西面,与四川省叙永县、合江县毗邻,低中山地、丘陵地形,属中亚热带温暖湿润季风气候区,年均温度18.2 ℃,年均降水量1 280 mm,海拔228~1 280 m,主要竹种有毛竹Phyllostachyspubescens、慈竹Neosinocalamusaffinis、撑绿竹Bambusapervariabilis×Dendrocalamusdaii、麻竹Dendrocalamuslatiflorus、孝顺竹Bambusamultiplex等(王润秋,2014)。四川省乐山市沐川县大楠镇属浅丘区兼有低山,年平均温度17 ℃,年均降水量1 333.9 mm,海拔336~818 m,主要竹种有慈竹、撑绿竹、绵竹Lingnaniaintermedia、苦竹Pleioblastusamarus等(陈其勇等,2014)。2016年7—8月长足大竹象出土盛期,分别在四川省乐山市沐川县大楠镇(104°0′6.93″E,28°58′5.06″N,海拔532 m)和贵州省赤水市大同镇(105°40′40.12″E,28°30′28.33″N,海拔358 m)慈竹林中采集雌雄成虫各15只,用取自两地的新鲜慈竹笋分开成对喂养,交配后收集受精卵。饲养环境:温度25 ℃、相对湿度75%、光周期12L∶12D,每2 d更换1次竹笋。
1.2 主要仪器及试剂
离子溅射仪Cressington108(Cressington,英国)、扫描电镜Inspect F50(FEI,美国)、Leica EZ4 HD体视镜(Leica,德国)等。所用试剂均为分析纯,购自成都科龙化工试剂厂。
1.3 解剖及扫描电镜观察
长足大竹象内、外生殖器的解剖方法参照汪淑芳等(2009),雌雄成虫各取15只进行解剖、扫描电镜观察。解剖后的内、外生殖器用足量生理盐水清洗3次后,放入4%多聚甲醛-磷酸二氢钠/氢氧化钠固定液中固定。参照杨瑞等(2014)的方法,将拟电镜观察的卵粒、雄性睾丸、雌雄性外生殖器等用固定液固定24 h以上,经30%、50%、70%、90%、100%乙醇进行逐级脱水,乙酸异戊酯置换处理10 min,经临界点干燥仪干燥,将干燥好的样品观察面向上粘台,用Cressington108 离子溅射仪真空喷金。将经上述处理过的样品置于Inspect F50扫描电镜下,观察其生殖结构形态特征并拍照。
1.4 数据统计与分析
用SPSS 16.0、Excel对数据进行统计分析,运用t检验进行差异显著性分析。利用ImageJ对卵细胞和精细胞直径、密度及外生殖器的长度等进行测量,数据以平均值±标准误表示。
2 结果与分析
2.1 生殖系统形态对比
乐山、赤水两地长足大竹象雌性成虫生殖结构均由1对卵巢、1对侧输卵管、中输卵管、交配囊、受精囊及产卵器组成,卵巢左右并列,呈乳白色,每个卵巢由2只卵巢管组成,卵巢管长约2.8 cm,每只卵巢管下端卵粒大而饱满,向上逐级减小,5~6粒卵,每侧卵巢管端丝汇成悬带;雄性成虫生殖结构均由1对睾丸、输精管、射精管、附腺及外生殖器组成,每侧睾丸由2只扁球状睾丸管组成(图1)。两地雌雄成虫生殖系统在形态结构上未观察到显著差异。
2.2 内生殖器超微结构
2.2.1雌虫卵黄颗粒对雌性长足大竹象受精卵内部进行电镜扫描,可见大量呈椭圆形或圆形的卵黄蛋白充塞在原生质网络内。电镜放大300倍,观察到乐山地区卵黄颗粒(图2:A)较赤水地区(图2:D)密实,放大至3 000倍、10 000倍后,发现乐山雌虫卵黄颗粒饱满且表面光滑(图2:B、C),而赤水雌虫卵黄颗粒表面粗糙、有镂空(图2:E、F)。测定卵黄颗粒密度、直径,每万立方微米内卵黄颗粒的平均密度为:乐山地区10.4个、赤水地区11.9个;卵黄颗粒平均直径为:乐山地区20.76 μm±1.33 μm、赤水地区15.26 μm±1.99 μm,二者差异有高度统计学意义(P=0.000 145)。
2.2.2雄虫睾丸乐山、赤水两地雄虫睾丸内部有明显差异,乐山地区睾丸内壁较为光滑(图3:A、B、C),赤水地区睾丸内壁上有许多褶皱(图3:D、E、F),两者内部均可观察到精细胞。每万立方微米内精细胞的数量为:乐山地区约400个、赤水地区约1 500个,相差近3倍;精细胞直径为:乐山地区5.60 μm±0.51 μm、赤水地区3.31 μm±0.27 μm,二者差异有高度统计学意义(P=0.000 004)。
2.3 外生殖器超微结构
2.3.1雌虫外生殖器扫描电镜结构乐山、赤水两地的产卵器鞘上均有成丛的刚毛,由产卵器鞘末端向上逐渐变短(图4:A、D),刚毛上着生刺状凸起,刚毛区基部均匀着生芽状小刺(图4:B、E),外生殖器中部有一片绒毛区(图4:C、F)。分别测量其外生殖器长刺和小刺的长度、密度、绒毛区宽度等,两地差异无统计学意义。

图1 乐山(A、B)、赤水(C、D)地区长足大竹象生殖系统Fig. 1 Reproductive system of Cyrtotrachelus buqueti from Leshan (A, B) and Chishui (C, D)
A, C. 雌虫female, B, D. 雄虫male; ovp. 产卵器ovipositor, C. 中输卵管common oviduct, L. 侧输卵管lateral oviduct, ov. 卵巢ovary, bc. 交配囊bursa copulatrix, S. 受精囊spermatheca, li. 悬带ligament, pe. 阳茎penis, mu. 肌纤维鞘muscle fiber sheath, va. 输精管vasa deferentia, sp. 睾丸管spermatic tube, te. 睾丸testis, ac. 附腺accessory gland

图2 乐山(A~C)、赤水(D~F)地区长足大竹象雌性成虫卵黄颗粒电镜扫描Fig. 2 The egg of female Cyrtotrachelus buqueti from Leshan (A-C) and Chishui (D-F) as determined by using scanning electron microscopy
A, D. 300×, B, E. 3 000×, C, F. 10 000×

图3 乐山(A~C)、赤水(D~F)地区长足大竹象雄性成虫睾丸电镜扫描Fig. 3 The testis of male Cyrtotrachelus buqueti from Leshan (A-C) and Chishui (D-F) as determined by using scanning electron microscopy
A, D. 5 000×, B, C, E, F. 10 000×

图4 乐山(A~C)、赤水(D~F)地区长足大竹象雌性成虫产卵器鞘电镜扫描Fig. 4 External reproductive organs of female Cyrtotrachelus buqueti from Leshan (A-C) and Chishui (D-F) as determined by using scanning electron microscopy
A, D. 200×, B, E. 2 000×, C, F. 5 000×
2.3.2雄虫外生殖器超微结构乐山、赤水两地雄虫外生殖器末端的阳茎均为黑褐色骨化管状结构,向腹面弯曲,背面上半部骨化,下端为膜质区带(图5:A、E),表面局部着生成丛的纤毛(图5:B、F)。横切面扫描电镜观察到阳茎为双层结构,内层萎缩形成层间中空(图5:C、G),内层不规则卷迭,呈空腔结构,内阳茎壁不光滑,具有突起状结构(图5:D、H)。分别测量阳茎壁厚度、内阳茎壁凸数目等,两地差异无统计学意义。
3 讨论
形态解剖表明,乐山、赤水两地不同种群密度的长足大竹象雌性成虫生殖系统形态结构未观察到显著差异,扫描电镜观察两地成虫外生殖器超微结构也未观察到显著差异,但两地成虫卵黄颗粒、精细胞在形态结构上的差异有高度统计学意义。超微结构研究表明,赤水地区低密度种群雌虫卵黄颗粒表面粗糙有镂空,直径显著小于乐山地区高密度种群,赤水地区雄虫睾丸内壁粗糙有褶皱,精细胞直径极显著小于乐山地区,每万立方微米内精细胞的数量超过乐山地区的2倍。由此可知,与乐山地区相比,赤水地区成虫生殖系统卵黄颗粒、睾丸的结构和大小及精细胞的数量均发生了显著变化,这可能导致赤水地区种群的繁殖力下降,种群数量低。
环境因素、遗传变异或遗传变异与环境互作均可引起长足大竹象生殖系统结构发生变异。赤水地区大面积竹林为长足大竹象的种群繁育提供了充足的食物来源。若是遗传变异导致种群数量大幅度降低,那么幸存个体在环境适宜、食源丰富的条件下应能大量繁殖,且长足大竹象是迁飞性昆虫,通过迁飞,亦可使其种群数量增加(Östmanetal.,2001;Eckertetal.,2008;Spearetal.,2010;He & Hubbell,2011)。由此推测,由环境因素引起赤水地区成虫睾丸、卵黄颗粒结构及精细胞大小产生变异的可能性较大。温度是影响昆虫生长发育和繁殖的重要因素之一,相关研究发现低温会延缓雌性苜蓿叶象甲Hyperapostica(张娜等,2010)和甜菜夜蛾Spodopteraexigua(韩兰芝等,2003)的性腺发育及卵子成熟;高温影响雄性棉铃虫Helicoverpaarmigera精细胞囊、睾丸和附腺的发育,导致畸形精子束产生(郭慧芳等,2002)。纬度偏南的广西南宁,年均温度高于赤水,但慈竹林生长环境相似,长足大竹象种群数量仍高于赤水地区(梁祖昂,2017)。本研究发现,年均温度相近的乐山和赤水两地的长足大竹象成虫在卵黄颗粒、睾丸的结构和大小及精细胞的数量等方面却有显著差异。由此推测,温度等气候因子导致两地长足大竹象成虫生殖结构变化的可能性不大。课题组实验检测还发现,赤水地区长足大竹象蛹室土壤中重金属Hg的含量相对较高,为乐山地区的1.5倍以上(资料未发表)。大量研究表明,Hg影响酶活性,对生命活动、昆虫种群可能造成毒害(沈维干,陈彦,2000;董杰影等,2004;楼哲丰等,2007)。长足大竹象的老熟幼虫在地下蛹室化蛹羽化的时间长达11个月,蛹室土壤Hg含量显著增加很可能会影响成虫生殖器官的分化,进而导致生殖细胞结构发生变异,降低赤水地区种群的繁殖能力和种群数量。因此,土壤重金属Hg含量的差异可能是导致两地长足大竹象生殖系统结构发生显著差异的重要原因,但还需进一步验证。
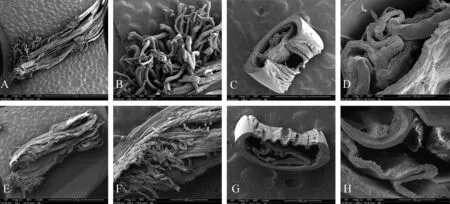
图5 乐山(A~D)、赤水(E~H)地区长足大竹象雄性成虫外生殖器电镜扫描
Fig. 5 External reproductive organs of maleCyrtotrachelusbuquetifrom Leshan (A-D) and Chishui (E-H) as determined by using scanning electron microscopy
A.阳茎基phallobase 43×, B. 阳茎基phallobase 150×, C. 阳茎横切面penis transection 150×, D. 阳茎横切面penis transection 1 000×,
E. 阳茎基phallobase 50×, F. 阳茎基phallobase 200×, G. 阳茎横切面penis transection 200×, H.阳茎横切面penis transection 1 000×
本研究首次比较了高、低种群密度地区地下生活史较长昆虫长足大竹象生殖系统超微结构的变化特点,研究结果对研究长足大竹象种群变化、种群发生等具有现实参考价值。
