混播种类和群体结构对豆禾牧草混播系统氮素利用效率的影响
2018-10-19朱亚琼关正翾郑伟王祥
朱亚琼,关正翾,郑伟,2*,王祥
(1.新疆农业大学草业与环境科学学院,新疆 乌鲁木齐 830052;2.新疆维吾尔自治区草地资源与生态重点实验室,新疆 乌鲁木齐830052)
豆科牧草与禾本科牧草混播/间套作(异行混播)/轮作能利用豆科牧草生物固氮作用增加豆禾牧草混播系统氮素的供应[1-3],并且可在一定程度上促进豆科牧草固氮效率的提高[4-6],还能促进系统中禾本科牧草对氮的吸收[5, 7],减少土壤无机氮的累积[8-9],从而提高整个混播系统氮素的利用效率[4, 10]。例如,王平等[2]研究表明,混播条件下紫花苜蓿(Medicagosativa)的固氮能力显著高于单播紫花苜蓿,混播提高了紫花苜蓿对大气氮素的固定能力。黑麦(Avenasativa)/红三叶草(Trifoliumpratense)异行混播、黑麦/豌豆(Pisumsativum)异行混播时,牧草氮素累积量和生物量都高于单播[11]。小麦(Triticumaestivum)/蚕豆(Viciafaba)间作显著提高了整个生育期内小麦地上部植株含氮量,显著增加了小麦叶和穗的氮含量,为生物产量的形成和经济产量的提高奠定了基础[12]。因此,豆科牧草根瘤菌固定的氮资源对豆禾混播草地提高土壤氮含量、增加草地氮产量和维持系统氮素平衡起着重要作用[7]。
豆科牧草通过生物固氮作用增加的氮素,一方面被自身生长所消耗,另一方面通过根接触转移或生物体死亡分解后释放到土壤中,再由相邻禾草吸收利用[13-15]。禾草利用豆科牧草固定的氮素途径有3条:即混播牧草可以通过吸收牧草根系的脱落物、根瘤和细小根系的腐烂分解和矿化的物质发生氮转移[13-14];通过泡囊-丛枝菌根(vesicular arbuscular mycorrhiza,VAMF)或丛枝菌根(arbuscular mycorrhiza,AM)菌丝桥的连接作用发生的氮素转移,转移的氮占作物吸收氮含量的0.7%~2.5%[16-17];禾本科通过根系分泌一些小分子的物质,如多糖、氨基酸等发生氮转移[18]。与此同时,豆禾牧草混播后,禾草与豆科牧草间存在着对氮素的竞争权衡,只有当土壤氮含量通过氮循环达到禾草与豆科牧草各自竞争优势平衡的水平时,两者才可以共存[19]。豆禾牧草共存的基础还包括存在空间上或时间上根系分布差异所导致的生态位分离,以及禾草、豆科牧草在养分利用方式上差异所导致的生态位分离(由于豆科牧草和禾本科牧草在利用土壤无机氮方面存在不同的竞争差异,而引起种植豆科牧草后的土壤无机氮比种植禾本科后的土壤无机氮变高的现象被称为“节约氮”),这些均能有效地降低豆禾牧草对土壤养分的竞争[20]。因此,豆科牧草生物固氮能力,豆禾牧草间转移氮素的数量和时间,以及豆禾牧草养分竞争状态等直接影响着混播中氮素效益,也是氮素能否被高效利用的关键。
现有的研究表明,并不是所有豆禾牧草混播体系均具有较高氮素利用效率,由于水分胁迫[21]、豆科牧草养分竞争劣势[2]和群体结构[22]等因素的影响,导致豆科牧草转向对土壤无机氮的吸收,氮素利用效率并没有比单播/单作高[23]。特别是混播群体结构,不仅影响了地上光资源的竞争结局,还影响了根系生长[24],从而导致地下养分竞争格局发生变化。祁军等[25]、于辉等[26]和杨峰等[27]研究结果表明,豆禾混播牧草群体结构还影响了冠层光合有效辐射、地上种间竞争格局和根系形态,从而导致混播群体生物产量与经济产量的变化。如何筛选出较优的混播组合与混播群体结构,权衡地上光资源竞争与地下根系养分竞争,则需要利用氮素利用效率测度豆禾牧草混播复合系统高效生产机制。因而,需要通过深入探讨混播种类、混播群体结构与混播系统氮素利用效率之间的关系,明确豆禾牧草混播产生氮素补偿效应及其他生态位分离的机理,推动豆禾混播系统高效生产潜能的充分发挥。因此,本研究以无芒雀麦(Bromusinnermis)、猫尾草(Phleumpratense)、鸭茅(Dactylisglomerata)、红豆草(Onobrychisviciaefolia)、红三叶和紫花苜蓿6种豆禾牧草在混2组合(红豆草和无芒雀麦)、混4组合(无芒雀麦、红豆草、猫尾草和红三叶)和混6组合(6种牧草混合)3种混播种类的豆禾混播草地为研究对象,将种群空间距离(行距)、种群空间作用方式(同行/异行/异行阻隔)作为豆禾牧草群体结构的变化因素,从豆科牧草固氮量、转氮量及其氮素利用效率对草地氮产量、生物产量的贡献角度出发,利用盆栽试验分析和比较豆禾混播牧草在不同混播种类与混播群体结构中的氮素利用效率变化,重点解析混播群体结构与豆科牧草生物固氮能力和氮素利用效率的关系,明确通过合理混播种类与混播群体结构提高混播系统氮素利用效率的基本机制,为阐明豆禾牧草混播复合系统高效生产机制做出贡献。
1 材料与方法
1.1 试验区概况
试验地点位于新疆伊犁哈萨克自治州昭苏县的昭苏马场(E 81°03′-81°05′,N 42°38′-43°15′),该区域气候类型属于温带山区半湿润易旱冷凉类型。根据国家气象数据网的数据(站点号51437),2013-2015年,研究区年均温4.44 ℃,年最高温均值为31.38 ℃,年最低温均值为-22.98 ℃,年均降水量482.73 mm,生长季(5-10月)降水量均值为358.20 mm,占年降水量的74.20%,全年日照时长2499.53 h。
1.2 盆栽试验设计
本试验选用的材料为无芒雀麦、猫尾草、鸭茅、红豆草、红三叶和紫花苜蓿6种豆禾牧草,试验前从昭苏马场山地草甸取回0~20 cm的土壤,去除根系、凋落物和大石块,混匀,作为盆栽试验基质。昭苏马场山地草甸土壤为黑钙土,土壤有机质含量为13.63%~14.89%,全氮、全磷、全钾量分别为9.53 g·kg-1,2.16 g·kg-1,12.20 g·kg-1,碱解氮、有效磷、有效钾含量分别为459.20 mg·kg-1,144.43 mg·kg-1,473.58 mg·kg-1[28]。将花盆置于室外自然环境,把混匀的土壤放入花盆中,低于花盆边缘2 cm左右,并控制每个花盆土壤具有相同的容重。
试验设3种混播种类组,分别为:合混2(无芒雀麦+红豆草)、混4(无芒雀麦+红豆草+猫尾草+红三叶)和混6(无芒雀麦+鸭茅+红豆草+猫尾草+红三叶+紫花苜蓿);4种群落结构处理,分别为同行混播+15 cm行距,异行混播+15 cm行距,异行阻隔混播+15 cm行距(中间用木板阻隔)和异行混播+30 cm行距,豆禾混播比例均为5∶5。单播无芒雀麦、猫尾草、鸭茅、红豆草、红三叶和紫花苜蓿作为对照,行距均为15 cm。每个花盆均只播种2行,所有处理均为3次重复;15 cm行距花盆大小为70 cm(长)×30 cm(宽)×30 cm(高),30 cm行距花盆大小为70 cm(长)×45 cm(宽)×30 cm(高),播种时距花盆边缘均为7.5 cm。混播与单播密度相同,混播比例按种子占单播重量的实际用价来计算,具体设计及播量见表1。
在上述花盆中施入同位素氮肥硫酸铵[(15NH4)2SO4],牧草生长旺期(2016年6月10日至6月17日)分2次(1周之内)均匀施入,每个花盆总施量为10 kg·hm-2纯N,将肥料与细沙以1∶10混匀后施入,轻灌以促进肥料均匀下渗。
1.3 测定项目与方法
1.3.1牧草产量及氮产量的测定 牧草产量:2016年8月15日在牧草结实期齐地面刈割,按混播牧草种类分开,及时置于65 ℃烘箱烘至恒重后记录各组成物种干重。
牧草N产量的测定:将烘干样品带回室内,另取样品粉碎后过40目筛(0.425 mm),采用H2SO4-H2O2消煮,利用ZDDN-Ⅱ型凯式定氮仪(中国)测定牧草样品的粗蛋白(crude protein, CP)含量。并根据牧草产量换算成CP产量,再折算成牧草N产量,具体计算方法参见杨胜[29]的方法。

表1 豆禾混播草地混播比例与播量Table 1 Mixed sowing ration and sowing quantity in legume-grass mixture
1.3.2豆科牧草固氮量、转氮量及其氮素利用效率的测定15N自然丰度法的理论基础:在大气中,15N作为一种较稳定的同位素,其丰度是稳定而均匀的,原子丰度为0.3663%,被认为是15N标准同位素丰度[30]。通常以样品中的15N与标准自然丰度(大气15N丰度)的千分差(δ15N)表示其相对丰度,大气的δ15N为零,如下所示。
式中:δ15N为样品的15N与大气15N的千分差;atom%15N (sample)为样品15N原子丰度;atom%15N (standard)为大气标准15N原子丰度(0.3663%)。
牧草样品氮丰度测定:2016年8月15日在牧草结实期收获全株,按豆禾牧草分开,将全株样品置于65 ℃烘箱中烘至恒重,初步剪碎后,再经球磨仪粉碎后过120目筛(0.124 mm)制成待测样品。采用Delta VAdvantage气体稳定同位素质谱仪(Thermo Finnigan, German)测定牧草样品氮丰度。
1)应用生物固氮率(percentage of nitrogen derived from air, PN)、生物固氮量(amount of nitrogen derived from air, WN)和转氮率(percentage of nitrogen derived from legume, PNt)、转氮量(amount of nitrogen derived from legume, WNt)测度混播草地的氮素利用效率,其计算公式为:
式中:A%EMl为混播(包括同行与异行混播)中豆科牧草体内15N丰度;A%EG为单播禾草体内15N丰度;A%EMg为混播(包括同行与异行混播)中禾草体内15N丰度;YN1为混播豆科牧草总氮素产量;YNg为混播禾草总氮素产量[2, 30]。
2)生物固氮对草地氮产量的贡献率(contribution of biological nitrogen fixation, CBNF)计算公式为:
生物固氮贡献率(CBNF)=(WN/草地总氮产量)×100%
1.3.3牧草营养竞争比率及生态位分离 营养竞争比率(competition ratio, CRpm)是度量植物养分吸收能力强弱的一种指标[31]。本试验用豆科牧草相对禾本科牧草对养分的竞争比率来衡量养分竞争能力:
CRpm=(Uil/Usl×Fl)/(Uig/Usg×Fg)
式中:Uig和Uil分别为混播禾草和豆科牧草的养分吸收量;Usg和Usl分别为单播禾草和豆科牧草的养分吸收量;Fg和Fl分别为混播中禾草和豆科牧草所占比例。当CRpm>1时,表明豆科牧草比禾草营养竞争能力强;当CRpm<1时,表明豆科牧草比禾草营养竞争能力弱。
相对产量(relative yield, RY)和相对产量总和(relative yield total, RYT)可以反映混播牧草间的竞争格局与竞争结果[13, 21],可作为养分竞争的重要参照目标。
禾草相对产量(RYG)=Ygl/(pYg)
豆科牧草相对产量(RYL)=Yll/(qYl)
式中:Ygl是在混播条件下禾草的单位面积产量,Yg是单播条件下禾草的单位面积产量;Yll是混播条件下豆科牧草的单位面积产量,Yl是单播条件下豆科牧草的单位面积产量;p为禾草在混播中所占比例;q为豆科牧草在混播中所占比例;RYG=1表明禾草的种内竞争与豆科牧草的种间竞争相似;RYG>1表明禾草的种内竞争大于其种间竞争;RYG<1表明种间竞争要大于种内竞争[13]。

式中:Yij为种i与种j混播时种i的草产量;Yii为种i单播时的草产量;Yji为种j同种i混播时种j的草产量;Yjj为种j单播时的草产量。RYT值表明混播牧草间的相互关系及对同一环境资源的利用情况;RYT>1时,混播牧草占有不同的生态位,利用不同的资源,有一定共生关系;RYT=1时,混播牧草种间利用共同的资源;RYT<1时,表示混播牧草间相互拮抗[32]。
1.4 数据处理与分析
采用Excel 2010、SPSS 20.0对数据进行统计分析,统计完成后用Origin 8.0制图。采用q检验法(Student-Newman-Keuls,SNK)和最小差异显著法 (Least—significant difference,LSD)进行不同混播处理间牧草产量、牧草N产量、生物固氮率、生物固氮量、转氮率、转氮量和营养竞争比率的差异显著性检验(P<0.05),采用最小二乘法对牧草产量与生物固氮率、生物固氮量、转氮率、转氮量和生物固氮对草地氮产量的贡献率进行回归分析。
2 结果与分析
2.1 不同混播处理牧草产量和N产量的比较
牧草产量和N产量直接反映了不同混播组合氮素利用效率的最终结果。如图1所示,从混播种类来看,混播处理牧草产量显著高于单播处理(P<0.05);2种豆禾牧草混播(混2)牧草产量显著高于6种牧草混播(混6)(P<0.05),混6牧草产量显著高于4种豆禾牧草混播(混4)。从混播群体结构来看,豆禾牧草异行混播+行距30 cm(Int30)牧草产量显著高于其他群体结构(P<0.05);豆禾牧草同行混播+行距15 cm(Mix15)与豆禾牧草异行混播+行距15 cm(Int15)牧草产量、豆禾牧草异行混播+行距15 cm+根系阻隔(Int45)无显著差异(P>0.05),但显著高于单播(P<0.05);Int15牧草产量显著高于Int45(P<0.05)。从各处理来看,混2+豆禾牧草异行混播+行距30 cm(M2-30)和混6+豆禾牧草异行混播+行距30 cm(M6-30)牧草产量最高,其次为混2+豆禾牧草异行混播+行距15 cm(M2-15)和混2+豆禾牧草同行混播+行距15 cm(T2-15),且上述4种处理的牧草产量显著高于混4、混6(M6-30除外)和单播各处理的牧草产量(P<0.05)。
由图2所示,从混播种类来看,牧草N产量与牧草产量变化规律类似,但混2牧草N产量与混6无显著差异,混6牧草N产量也与混4无显著差异(P>0.05)。从混播群体结构来看,牧草N产量与牧草产量变化规律也类似。从各处理来看,M6-30、M2-30、M2-15和T2-15牧草N产量也较高,其他处理的牧草N产量变化规律与牧草产量相似。
双因素(混播种类、混播群体结构)方差分析表明(表2和表3),混播种类与混播种类×混播群体结构的交互作用对牧草产量的影响极显著(P<0.01),混播群体结构对牧草产量的影响显著(P<0.05);混播群体结构与混播种类×混播群体结构的交互作用对N产量的影响极显著(P<0.01),混播种类对N产量的影响显著(P<0.05)。上述结果表明,随着混播种类和混播群体结构的改变,牧草产量和N产量发生显著变化,混2、混6+异行混播30 cm行距的处理具有较好的生产性能。
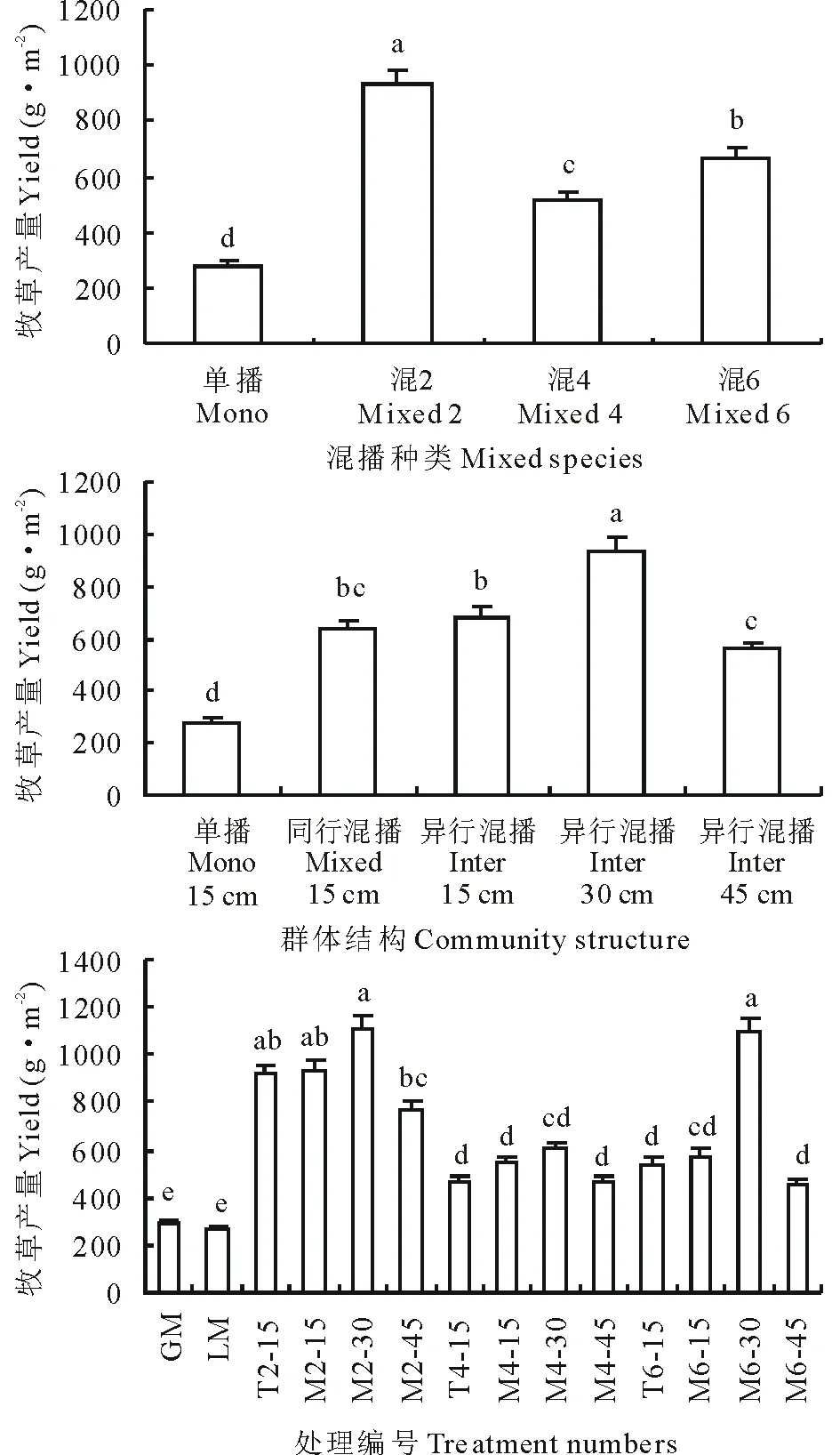
图1 不同混播种类、群体结构和处理间牧草产量的比较Fig.1 Effects of species diversity, community structures and treatments on forage yield

图2 不同混播种类、群体结构和处理间牧草N产量的比较Fig.2 Effects of species diversity, community structures and treatments on forage N yield
不同小写字母表示不同处理间的差异显著(P<0.05),下同。Different letters within the same figure indicates significant difference (P<0.05), the same below.
2.2 不同混播处理固氮量、氮素利用效率及贡献率的比较
禾草与豆科牧草混播后,可得到豆科牧草直接或间接转移过来的氮素。由图3所示,从混播种类来看,混2固氮量(WN)显著高于混4和混6(P<0.05);混2和混6转氮量(WNt)显著高于混4(P<0.05)。从混播群体结构来看,Int30和Mix15的WN显著高于其他群体结构(P<0.05);Int30的WNt显著高于Mix15和Int15(P<0.05),且三者均显著高于Int45(P<0.05)。从各处理来看,M2-30、T2-15和混4+豆禾牧草同行混播+行距15 cm(T4-15) WN显著高于其他处理(P<0.05);M2-30、M6-30和混6+豆禾牧草异行混播+行距15 cm(M6-15)具有较高的WNt。
由图4所示,从混播种类来看,固氮率(PN)和贡献率(CBNF)在各混播种类处理间无显著差异(P>0.05);混2和混6的转氮率(PNt)显著高于混4(P<0.05)。从混播群体结构来看,PN在各混播群体结构处理间无显著差异(P>0.05); Int30的PNt显著高于Mix15和Int15(P<0.05), Mix15和Int15的PNt显著高于Int45(P<0.05),Int45的PNt值为负值;Int30和Int15的CBNF显著高于Mix15和Int45(P<0.05)。从各处理来看,M2-15的PN显著小于M2-30、T4-15、混4+豆禾牧草异行混播+行距30 cm(M4-30)、M6-15和M6-30(P<0.05),与其他处理无显著差异(P>0.05);M2-30、T2-15、M6-15和M6-30的PNt显著高于其他处理(P<0.05),M6-30的PNt值最高,可达37.13%,豆禾牧草异行混播+行距15 cm+根系阻隔的各处理PNt值为负值,显著小于其他处理(P<0.05);M2-15、M4-30、M6-30的CBNF较高,分别为53.26%、51.14%和54.23%,与T2-15、混4+豆禾牧草异行混播+行距15 cm+根系阻隔(M4-45)和M6-15无显著差异(P>0.05),但显著高于其他处理(P<0.05)。

图3 不同混播种类、群体结构和处理的固氮量Fig.3 Effects of species diversity, community structures and treatments on nitrogen fixation amount

图4 不同混播种类、群体结构和处理氮素利用效率Fig.4 Effects of species diversity, community structures and treatments on nitrogen use efficiency
不同小写字母(a, b, c…)表示不同处理间固氮量的差异显著;不同大写字母(A, B, C…)表示不同处理间转氮量的差异显著;P<0.05。Different letters (a, b, c…) within the same figure indicates significant difference in the amount of nitrogen derived from air (WN), different capital letters (A, B, C…) within the same figure indicates significant difference in the amount of nitrogen derived from legume (WNt),P<0.05. 固氮量:Amount of nitrogen fixation,转氮量:Amount of nitrogen derived from legume. 不同小写字母(a, b, c…)表示不同处理间固氮率的差异显著;不同大写字母(A, B, C…)表示不同处理间转氮率的差异显著;不同字母(a′, b′, c′…)表示不同处理间贡献率的差异显著;P<0.05。Different letters (a, b, c…) within the same figure indicates significant difference for the percentage of nitrogen derived (PN), different capital letters (A, B, C…) within the same figure indicates significant difference for the percentage of nitrogen derived from legume (PNt), different letters (a′, b′, c′…) within the same figure indicates significant difference for the contribution,P<0.05. 固氮率:Percentage of nitrogen derived,转氮率:Amount of nitrogen derived from legume, 贡献率:Contribution.
双因素(混播种类、混播群体结构)方差分析表明(表3),混播种类、混播群体结构与混播种类×混播群体结构的交互作用对的WN的影响均不显著(P>0.05);仅混播群体结构对WNt的影响显著(P<0.05);混播种类、混播群体结构与混播种类×混播群体结构的交互作用对CBNF的影响也均不显著(P>0.05)。上述结果表明,随着混播群体结构的改变,混播群体的WNt发生了显著变化,异行混播30 cm行距的处理具有较高的固氮能力。
2.3 不同混播处理氮素营养竞争与生态位分离的比较
由表2可知,从混播种类来看,混4的豆科牧草吸氮量显著高于混6(P<0.05),但与豆科牧草单播(LM)无显著差异(P>0.05);混4的禾草吸氮量也较高,显著高于禾草单播(GM) (P<0.05),但与混2、混6无显著差异(P>0.05);混2的营养竞争比率(CRpm)较高,但与混4、混6也无显著差异(P>0.05)。从混播群体结构来看,Mix15的豆科牧草吸氮量显著高于Int15和Int30(P<0.05), 与Int45无显著差异(P>0.05); Int30的禾草吸氮量显著高于Mix15和Int45(P<0.05),与Int15无显著差异(P>0.05);Mix15和Int45的CRpm显著高于Int15和Int30(P<0.05)。从各处理来看,M4-45具有较高的豆科牧草吸氮量,与T2-15和T4-15无显著差异(P>0.05),但显著高于其他处理(P<0.05);M4-30、M4-15具有较高的禾草吸氮量,与M2-30无显著差异(P>0.05),但显著高于其他处理(P<0.05);M4-45还具有较高的CRpm,与T2-15、T4-15、M2-45和T6-15无显著差异(P>0.05),但显著高于其他处理(P<0.05)。且多数处理中禾草的吸氮量大于豆科牧草。双因素(混播种类、混播群体结构)方差分析表明(表3),混播种类对CRpm的影响不显著(P>0.05),混播群体结构对CRpm的影响极显著(P<0.01),混播种类×混播群体结构的交互作用对CRpm的影响显著(P<0.05)。
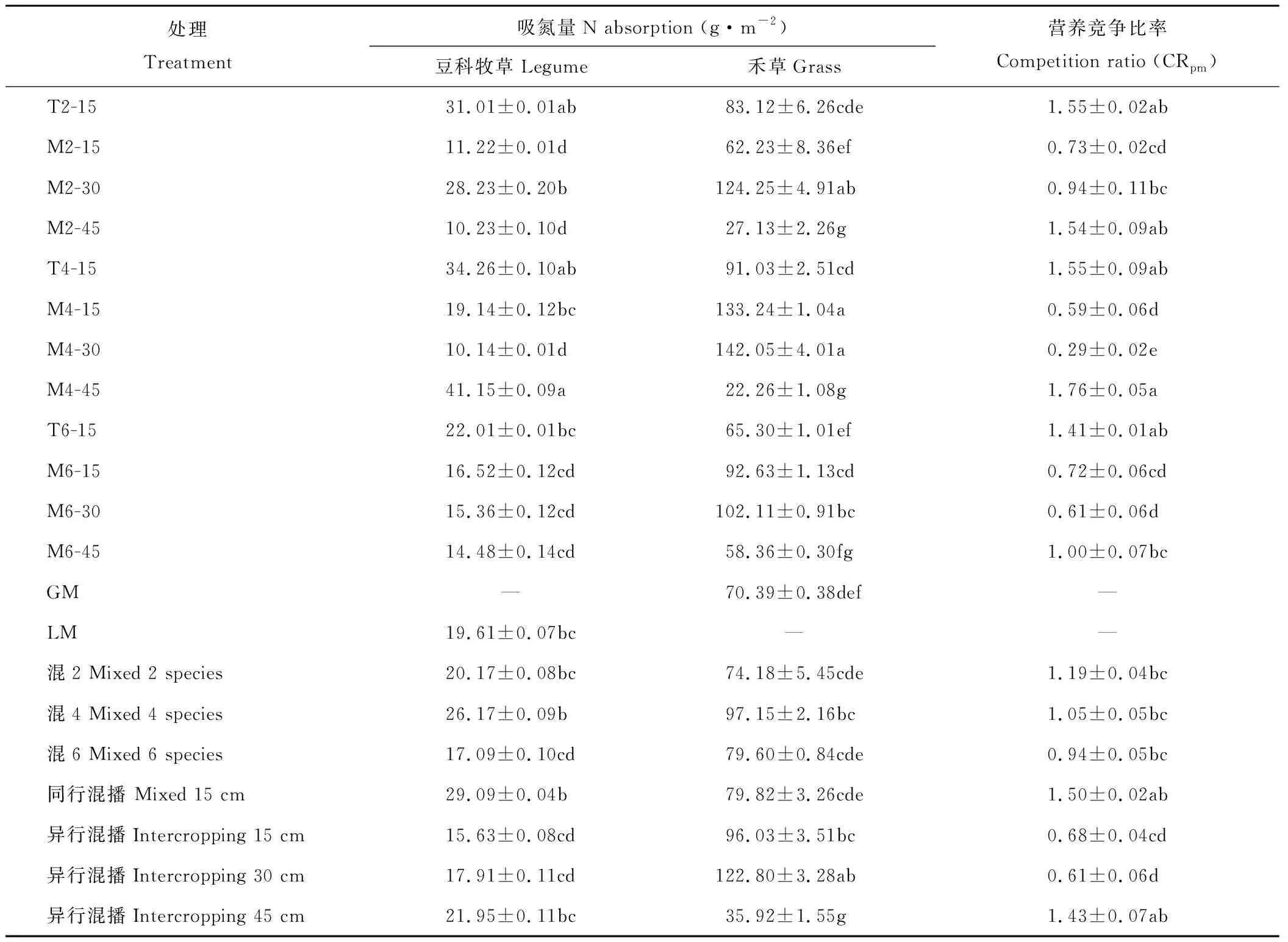
表2 不同混播种类、群体结构和处理间吸氮量及营养竞争比率(CRpm)Table 2 Effects of species diversity, community structures and treatments on nitrogen absorption amounts and competition ratios
注:同列不同小写字母表示不同处理间的差异显著,P<0.05。
Note:Different letters within the same column indicates significant difference,P<0.05.
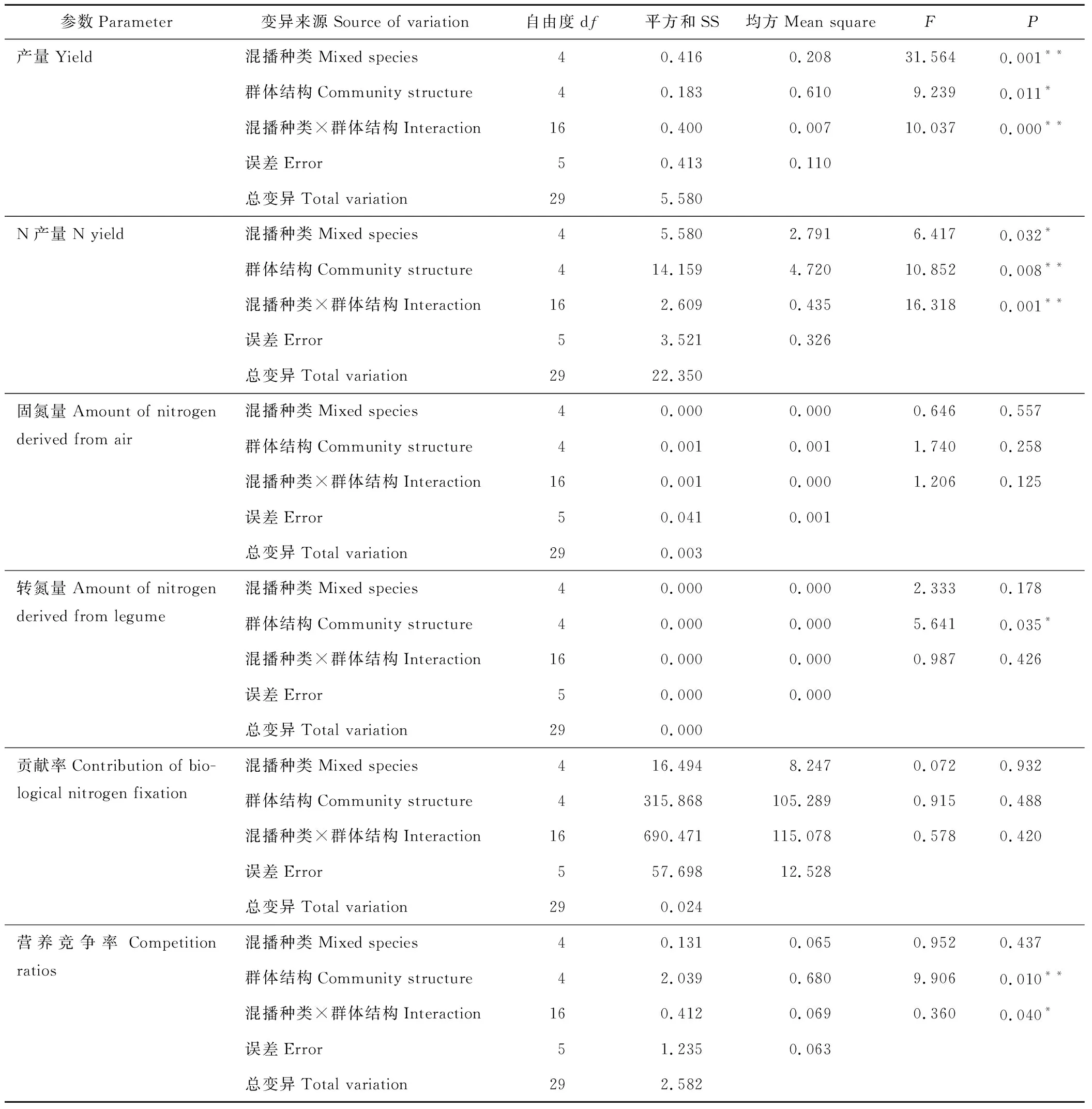
表 3 双因素方差分析Table 3 The two-way ANOVA
注:**表示在0.01水平(双侧)上极显著相关;*表示在0.05水平(双侧)上显著相关。
Note:** show significant correlation at 0.01 level (double side); * show significant correlation at 0.05 level (double side).
如图5所示,从混播种类来看,混4、混6豆科牧草相对产量(RYL)、相对产量总和(RYT)显著高于混2 (P<0.05);各混播种类间禾草相对产量(RYG)无显著差异(P>0.05)。从混播群体结构来看,Int30具有较高的RYL,显著高于Int15和Mix15(P<0.05),但与Int45无显著差异(P>0.05);Int30还具有较高的RYG,显著高于Int15和Int45(P<0.05),但与Mix15无显著差异(P>0.05);Int30的RYT显著高于其他混播群体结构(P<0.05)。从各处理来看,M4-30具有较高的RYL、RYG和RYT,其中,M4-30的RYL与M6-15、M6-30和M6-45无显著差异(P>0.05),但显著高于其他处理(P<0.05);M4-30的RYG与T2-15、M2-30无显著差异(P>0.05),但显著高于其他处理(P<0.05);M4-30的RYT显著高于其他处理(P<0.05)。上述结果表明,随着混播群体结构的改变,混播群体的营养竞争率和竞争格局发生了显著变化,异行混播30 cm行距的处理具有较低的豆科牧草氮素营养竞争率以及豆禾牧草生态位分离,具有较高的资源利用效率。
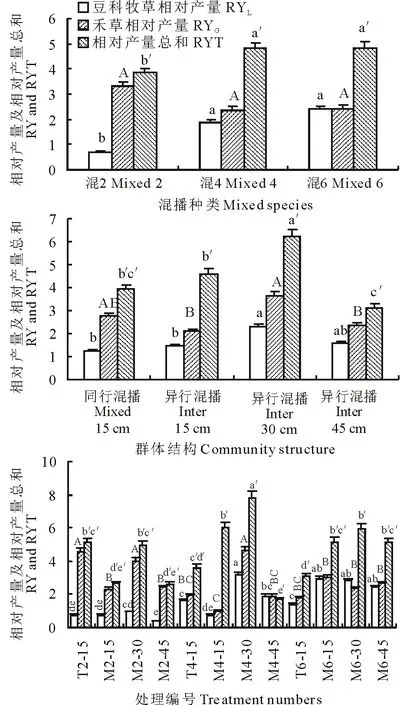
图5 不同混播种类、群体结构和处理间相对产量及相对产量总和的比较 Fig.5 Effects of species diversity, community structures and treatments on relative yield and relative yield total 不同小写字母(a, b, c…)表示不同处理间豆科牧草相对产量的差异显著;不同大写字母(A, B, C…)表示不同处理间禾草相对产量的差异显著;不同字母(a′, b′, c′…)表示不同处理间相对产量总和的差异显著;P<0.05。Different letters (a, b, c…) within the same figure indicates relative yield of legume significant difference, different capital letters (A, B, C…) within the same figure indicates relative yield of grass significant difference, different letters (a′, b′, c′…) within the same figure indicates relative yield total significant difference, P<0.05. RY: Relative yield; RYT: Relative yield total. 下同 The same below.豆科牧草相对产量:Relative yield of legume,禾草相对产量:Relative yield of grass,相对产量总和:Relative yield total.
2.4 牧草产量和相对产量总和与固氮量、固氮效率和营养竞争等参数的关系
如图6所示,对牧草产量和RYT与固氮能力、固氮效率和营养竞争等参数之间进行拟合发现(n=12):牧草产量与WNt呈极显著正相关关系(P<0.01),与WN、PN、PNt呈显著正相关关系(P<0.05),与CBNF和CRpm呈显著负相关关系(P<0.05);牧草产量与WN、WNt、PNt和CRpm间的关系可用二项式函数模型表示,与PN和CBNF间的关系可用三项式函数模型表示。RYT与CRpm呈极显著负相关关系(P<0.01),与WN、PN和CBNF呈显著正相关关系(P<0.05),与WNt、PNt呈显著负相关关系(P<0.05);RYT与PN间的关系可用一元线性函数模型表示,与WNt、PNt和CBNF间的关系可用二项式函数模型表示,与WN间的关系可用三项式函数模型表示,与CRpm间的关系可用指数函数模型表示。上述结果表明,牧草产量与转氮量关联性较强,而与其他固氮效率与固氮量参数关联性较弱;相对产量总和与营养竞争比率关联性较强,而与固氮效率与固氮量参数关联性较弱。
3 讨论
3.1 混播种类对氮素利用效率和生产性能的影响
在间混作群体中,植物种之间的竞争和互补提高了产量和养分利用效率,主要原因包括间混作植物根系间作用对养分吸收的促进、作物产量形成的竞争-恢复、养分高效利用以及植物对非高效作物养分的有效化等[33]。大部分研究是通过豆科牧草与禾草/农作物(豆科牧草+禾草或豆科牧草+农作物)2种植物种的组合来研究氮素利用效率、生物固氮量和生产性能改善,如Snoeck等[31]利用15N自然丰度法研究了在咖啡种植园中套种豆科植物,咖啡对豆科植物所提供的氮素的利用状况,结果表明咖啡30%的氮素来源于豆科植物;朱树秀等[34]对老芒麦(Elymussibiricus)与紫花苜蓿混播盆栽进行了氮素转移的研究,结果表明,盆栽的当年老芒麦从与之混播的苜蓿固定的氮中获得植物生长所需的22.9%~33.4%的氮,次年获得56%的氮。而另一方面,有关植物多样性对牧草产量或生物量影响的研究则表明,随着混播种类的增加,牧草产量呈增加的趋势[35],且混播种类多的草地长期利用的稳定性更好[28]。这是否意味着混播种类的增加有利于生物固氮量的增加,氮素利用效率的增加,从而使豆禾牧草养分竞争减弱,生态位分离,产生互补作用,生产性能提升?本研究结果表明,与单播牧草相比,各混播种类组合牧草产量、N产量均显著增加,但混播种类的增加未使牧草产量、N产量进一步增加(图1和图2)。与此同时,随着混播种类的增加,固氮量呈减少趋势,转氮量、转氮率和贡献率均呈先下降后上升的变化规律(图3和图4);但豆科牧草相对产量呈增加趋势,禾草相对产量和氮素营养竞争率呈减少趋势,相对产量总和呈增加趋势(表2,图5)。由此可见,混播种类的增加为豆禾牧草间资源的高效利用提供了更多机会,混播系统使氮素的固定、传递及吸收效率提高,对氮素的养分竞争也随之下降,为豆禾牧草的生态位分离提供了重要途径。但混播种类或者说植物物种多样性对混播系统生态功能影响是复杂的,不能简单归结为种类数量增加,混播系统生态功能随之提高[36],其中植物种的功能属性可能更能决定系统的功能属性[37]。因此,在豆禾牧草混播系统中利用混播种类提升系统功能可能并不能达到满意效果,筛选高效养分利用的物种混播组合更为关键。
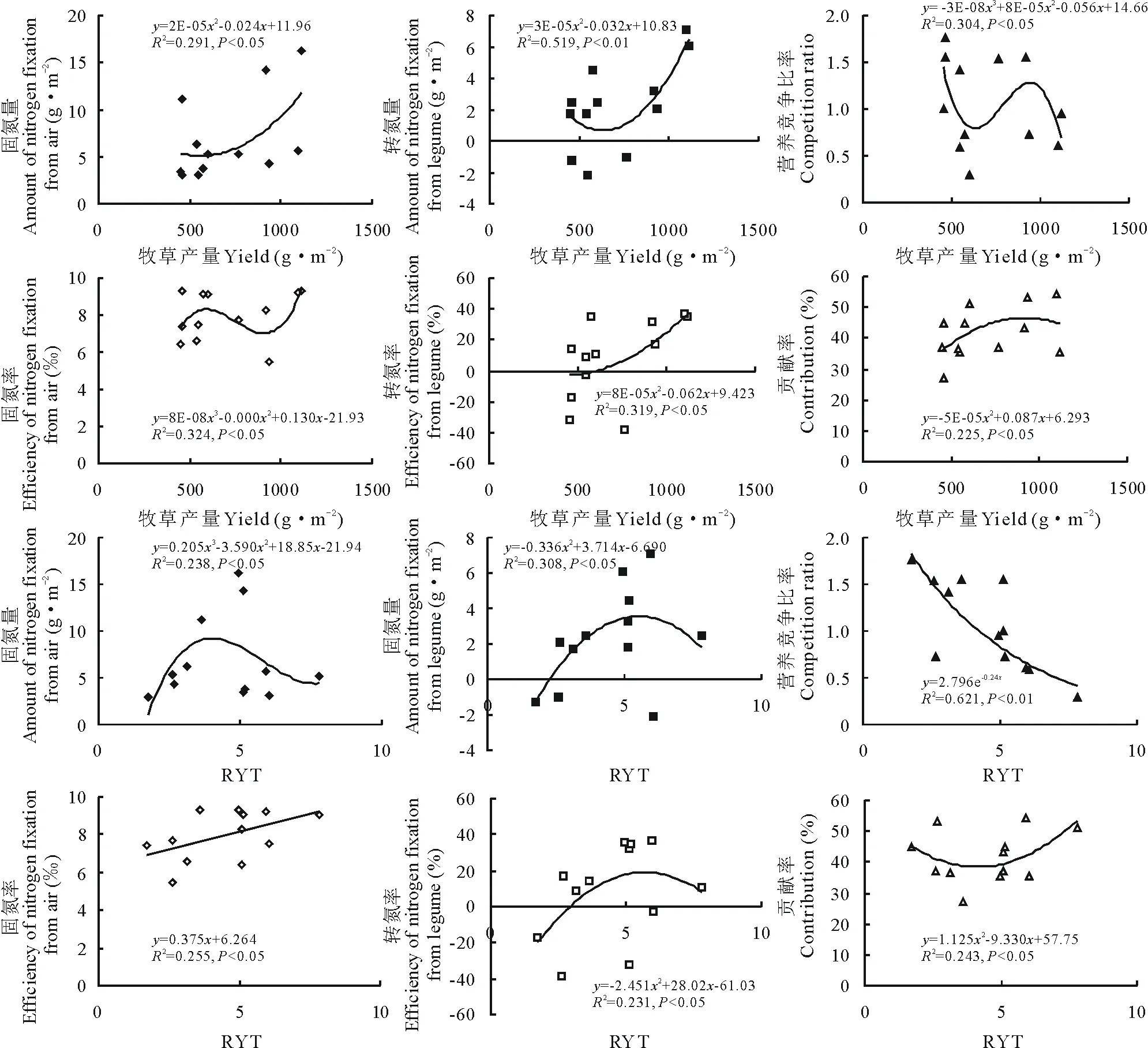
图6 牧草产量和相对产量总和与固氮量、转氮量、固氮率、转氮率、氮产量贡献率及营养竞争比率的关系Fig.6 The relationship between forage yield and WN, WNt, PN, PNt, CBNF and CRpm, and the relationship between RYT and WN, WNt, PN, PNt, CBNF and CRpm
3.2 群体结构对氮素利用效率和生产性能的影响
群体空间结构的变化(行距、间套作、同行、异行)直接改变了植物群体光能的截获、地上地下种间竞争过程以及产量潜力的发挥[38],还可影响土壤氮素的利用[39]以及混播物种间氮素利用共存关系[40]。Zhao等[40]认为间作群体的空间结构显著影响了豆科作物的生物固氮能力、间作系统的氮素利用效率以及“氮阻遏”效应。但也有研究认为不同间作群体的空间结构之间,氮素利用的互补作用差异不显著[5]。本研究结果表明,豆禾牧草异行混播+行距30 cm的群体空间结构具有较高的牧草产量、N产量、转氮量、转氮率、贡献率以及相对产量总和,但氮素营养竞争率较低(图1~图5,表2)。由此可见,优化豆禾牧草群体结构可以有效改善豆科牧草向禾草转移氮素的效率,促进二者之间在氮素利用上形成共存,显著提高混播系统生态功能。与此同时,牧草产量(或N产量)与转氮量呈极显著正相关(P<0.01),而相对产量总和与氮素营养竞争比率呈极显著负相关(P<0.01)(图6),这也进一步证明了豆科牧草转移氮素效率的提高有助于牧草产量的增加,而豆科牧草氮素营养竞争能力的下降促进了豆科与禾本科牧草之间氮素营养的共存,为混播系统生态功能的提升提供了重要途径。
4 结论
混播种类及其组合、混播群体的空间结构(行距、同行、异行、异行阻隔)改变了豆禾混播草地土壤养分利用与竞争环境,使得混播系统的氮素吸收、转移与利用效率发生变化,生态功能也随之改变。与单播相比,各混播组合显著提高了其资源利用效率和生态功能;但混播种类的增加并未显著增加混播群体生态功能,仅仅使混播群体发生共存的概率增加。从同行混播到异行混播以及行距增加,使得地下根系对氮素营养的竞争强度减弱,由豆科牧草固定及转移的氮素呈增加趋势,混播群体氮素利用的互补作用增强,种间相容性增加。但根系阻隔下,由豆科牧草固定及转移氮素的途径被阻断,混播群体互补作用被严重削弱。上述结果较好地反映了混播群体结构的优化可以使豆禾混播牧草根系氮素固定、转移、利用途径和效率得到提高,为混播系统生态功能的提升提供了重要基础。对混播种类与群体空间结构进行综合考虑,混2(无芒雀麦+红豆草)和混6(无芒雀麦+猫尾草+鸭茅+红豆草+紫花苜蓿+红三叶)+异行混播+行距30 cm的组合具有较高的氮素固定、转移效率,生产性能也较好。
