篮子鱼属线粒体基因组序列系统发育信息分析
2018-10-15黄小林杨育凯林黑着
黄小林, 杨育凯, 林黑着, 李 涛, 虞 为, 黄 忠
(1. 中国水产科学研究院 南海水产研究所 深圳试验基地, 深圳 518121; 2. 中国水产科学研究院 南海水产研究所 农业部南海渔业资源开发利用重点实验室, 广州 510300)
篮子鱼属(Siganus)在分类上隶属于鲈形目(Perciformes)刺尾鱼亚目(Acanthuroidei )篮子鱼科(Siganidae)[1],该属鱼类为小型浅海鱼类,栖息于近岸岩礁海域,广泛分布于太平洋、印度洋热带亚热带海域,在中国的南海、东海和台湾海域均有分布。该属鱼类共27种,在中国累积记录有13种[2-3]。篮子鱼肉嫩味美,富含高不饱和脂肪酸,营养价值高,深受消费者喜爱。食性杂食偏植食性,喜刮取附着在礁石或网衣上的藻类为食,具有饲料来源易解决、养殖成本较低、大小均可出售、饲养周期短、网箱套养还能清洁网衣等诸多优点,开发潜力大,已逐渐成为新的海水养殖对象[3]。
目前,国内外对篮子鱼的系统进化研究不多,属内部分种间形态差异不明显,物种不易鉴定,马强等[4]通过检视标本和查阅文献,确定目前在中国海域分布有篮子鱼11种,并依据形态学特征,比较分析了篮子鱼体形、体色、牙齿、头骨、椎骨、耳石等特征方面的种间差异。Kuriiwa等[5]通过线粒体基因Cytb和核基因ITS1对19种篮子鱼进行系统进化分析,在系统发育树中,褐篮子鱼与长鳍篮子鱼合为一支,狐篮子鱼与尖嘴篮子鱼合为一支,且在这两个分支中彼此分布相互嵌套,这与上述两对篮子鱼在形态上较难区分是吻合的。
mtDNA作为重要的核外遗传物质,因其基因组成简单、进化快、严格的母系遗传等优势,已被广大学者用于系统进化研究[6-14]。本文全面分析了篮子鱼线粒体基因组结构和变异位点,检测了进化过程中蛋白质编码基因受到的选择压力,分析了不同基因在系统发育分析中的适用性,并找出合适的分子标记。本研究旨在为篮子鱼系统分类和资源开发提供理论依据。
1 材料与方法
1.1 基因组结构和碱基差异性分析
从NCBI下载6种篮子鱼mtDNA全序列和2条相近属(刺尾鱼属)白胸刺尾鱼(Acanthurus.leucosternon)和纵带刺尾鱼(A.lineatus)[5]的mtDNA全序列(表1)。检索并统计每种篮子鱼mtDNA序列的长度和结构;在Clustal W[15]软件中比对并辅以人工校对,用软件Mega 5.0分析各基因的碱基含量,通过软件DnaSP 5.10分析各基因的变异位点等[16]。
1.2 系统发育分析
以白胸刺尾鱼和纵带刺尾鱼作为外群,用Mega 5.0软件中的邻接法 (NJ)构建6种篮子鱼线粒体基因组全序列、蛋白质编码基因及合并序列的分子系统进化树,采用Bootstrap法(重复1000次)检验各分支的置信度。
1.3 Ka/Ks分析与中性检验
物种进化速率由稳定性(负)选择、定向(正)选择和突变决定[9-13]。为检验篮子鱼mtDNA各基因在进化过程中受到的选择压力,通过软件DnaSP 5.10计算获得每个蛋白编码基因的非同义替换率(Ka)和同义替换率(Ks),并进行Tajima′s D中性检验。
1.4 系统发育信息分析
参照黄小林[11-13]等的方法,用Mega 5.0中的邻接法(K2P Model)构建分子系统树(NJ树)。通过NJ树中各分支Bootstrap的置信度大小来判断各分支的可靠性,从而评估mtDNA中不同基因对篮子鱼属系统进化分析的适用性,计算由单个基因构建的系统树各分支的置信度和与碱基信息量(置信度和/序列长度),将蛋白质编码基因合并序列构建的系统树定为标准树供比对,用于比较分析mtDNA中不同基因在系统进化分析中的适用性。
2 结果
2.1 线粒体基因组结构特征
mtDNA全长16 491~16 505 bp(表1),结构和排列顺序与其他硬骨鱼类一致[18-19](图1),包含蛋白质编码基因13个、rRNA基因2个、tRNA基因22个和D-loop,排列紧凑,无内含子。

表1 篮子鱼线粒体基因组基本信息

图1篮子鱼线粒体基因组结构
除ND4L外,mtDNA碱基组成上均出现明显的反G偏倚,各基因的变异位点分析(表2),其中,12S rRNA和16S rRNA基因最保守,变异位点比例分别为4.01%和6.91%,其次为COX2(12.01%)和ATP8(14.29%),最高的是ND3基因(22.77%)。
2.2Ka/Ks分析及Tajima′s D中性检验
13个蛋白编码基因的Ka/Ks[18]比值(表3、图2),所有蛋白编码基因Ka/Ks均远小于1(0.015~0.62),表明在进化过程中篮子鱼线粒体13个蛋白编码基因受到稳定性(负)选择作用较强;Tajima′s D检验[19]结果与Ka/Ks分析一致(Tajima′sD=0.123 74~0.615 14,P>0.10)。篮子鱼线粒体13个蛋白编码基因中,ATP8的Ka/Ks最大(0.064),表明该基因在进化过程中受到的稳定性(负)选择压力较小,之后是ND2和ND6(0.063和0.062)。反之,Cox1和ND4L基因的Ka/Ks最小(分别是0.015和0.021),表明这两个基因在进化过程中受到的稳定性(负)选择压力较大。
2.3 系统发育分析
mtDNA及蛋白编码基因合并序列的NJ树(图3和图4),全序列与蛋白质编码基因合并序列所构建的NJ树聚类关系一致。作为外类群的白胸刺尾鱼和纵带刺尾鱼位于树的底部。篮子鱼分子系统树结构:尖嘴篮子鱼与狐篮子鱼最先聚成一支,再依次与眼带篮子鱼和点篮子鱼聚类,之后与褐篮子鱼和长鳍篮子鱼聚成的一支聚类。尖嘴篮子鱼与狐篮子鱼的亲缘关系较近,褐篮子鱼和长鳍篮子鱼亲缘关系较近。

表2 篮子鱼mtDNA及不同区域的序列长度、G+C含量(%)、变异位点数(%)置信度和(N)及评价
注:表中P(poor)差;M(middle)中等;G(good)好;VG(very good)很好

表3 篮子鱼属线粒体蛋白质编码序列Ka/Ks分析及Tajima′s D中性检验
2.4 系统发育信息分析
mtDNA全序列、蛋白编码基因合并序列及单基因序列构建的系统发育树的置信度及每个碱基的平均信息量(表2)。mtDNA全序列和蛋白编码基因合并序列的置信度和最高(N=400),高于各单基因序列,但其单碱基信息量(分别为0.03和0.04)低于所有单基因序列。单基因序列中,置信度和最高的是ND4和ND5基因(N=399),其次是COX1、Cytb和12S rRNA基因(N=398),单碱基信息量最大的是ATP8基因(N/L=2.51)。

图2 蛋白质编码基因Ka/Ks分析

图3 线粒体基因组全序列 NJ 树
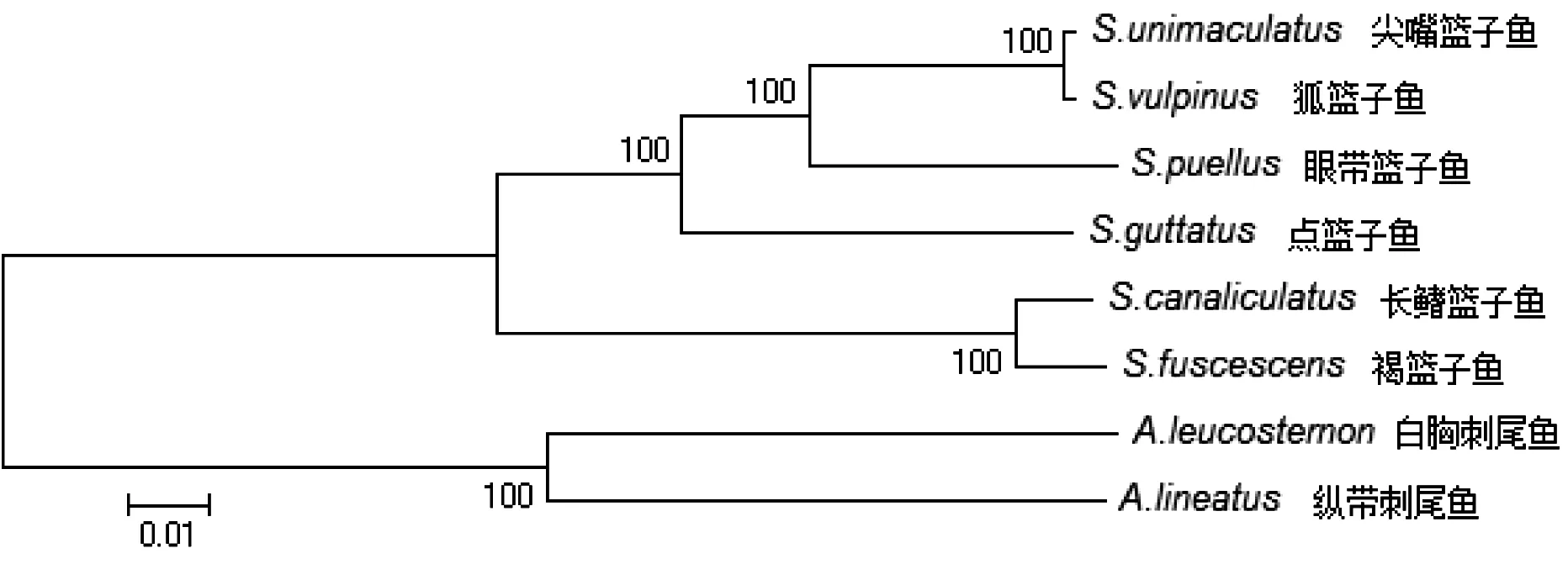
图4 蛋白编码基因合并序列 NJ 树
对比标准树(图4)发现,除ND4L基因系统树(图5)中物种聚类关系存在差异外,其余基因构建的系统树(未列出)与标准树物种聚类关系一致。
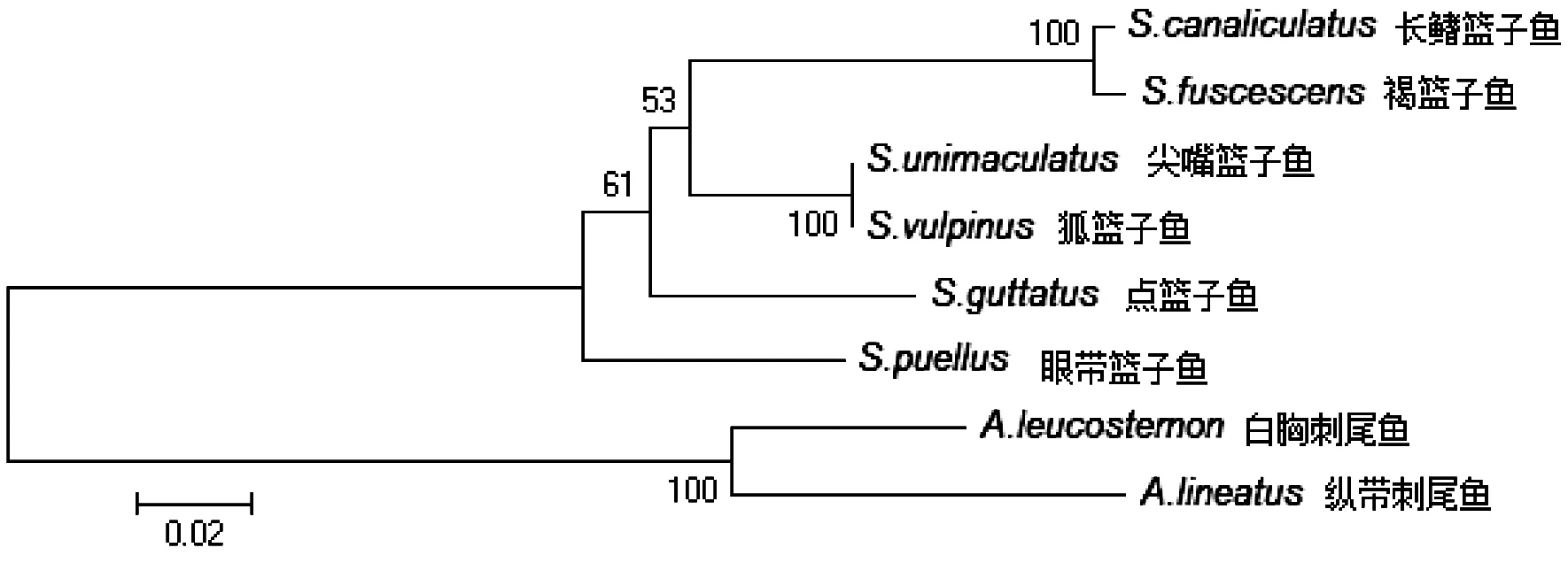
图5 基于ND4L基因的 NJ 树
3 讨论
蛋白编码基因中碱基的非同义突变造成转录的氨基酸不同,导致蛋白质原有功能发生改变,在进化过程中,这部分突变被视为有害突变而被稳定性(负)选择所修复;同义突变不造成氨基酸改变,被认为是中性或近中性的而被随机遗传漂变所固定。因此,体现出来的差异是非同义替换率小于同义替换率(Ka
本文研究的篮子鱼线粒体基因组中,所有蛋白编码基因的Ka/Ks(表3,图2)均远小于1,且Tajima′sD=0.123 74~0.615 14(P>0.10)均不显著大于或小于0,表明篮子鱼线粒体基因组蛋白编码基因进化过程中均受到较强的稳定性(负)选择作用。这与短尾派(Brachyura)[20]、中国明对虾(Fenneropenaeuschinensis)[21]、金线鲃(Sinocyclocheilusgraham)[22]等物种的线粒体基因组研究结论一致。ATP8基因的Ka/Ks最大,这在之前的研究中多有报道,其中海胆纲[26]线粒体基因组时发现ATP8基因Ka/Ks大于1,存在定向(正)选择。而鳀科鱼类[13]线粒体全基因组研究发现ND6基因Ka/Ks(1.139)大于1,存在定向(正)选择,这现象在鱼类mtDNA中极少出现,值得进一步研究,或许与在计算ND6基因Ka和Ks值时未将序列反向有关,因为在mtDNA结构中,8个tRNA和ND6在轻链上编码,其余基因都由重链编码,重链和轻链上的复制方向相反,因此在计算ND6基因的Ka和Ks值时,应先将序列反向。
在NJ树中,褐篮子鱼和长鳍篮子鱼位于系统树根部位置,是本研究6种篮子鱼中最早演化出来的一枝,之后相继分化出点篮子鱼、眼带篮子鱼、狐篮子鱼和尖嘴篮子鱼,这种进化顺序与这几种篮子鱼的体色变化是一致的,从褐篮子鱼和长鳍篮子鱼的棕褐色→点篮子鱼的淡蓝绿色→眼带篮子鱼的黄色至橙黄色→狐篮子鱼和尖嘴篮子鱼的鲜黄色,体色变化越来越艳丽,表明篮子鱼属鱼类体色在进化过程起到了一定的正向选择作用,值得进一步研究。
系统发育信息分析中,ATP8和ND3基因虽然有较高的单碱基信息量,但置信度和较低,不宜选为理想分子标记。综合表2中分析数据,篮子鱼mtDNA各基因用于种群遗传结构和系统发育分析的适用性顺序:COX1、ND4和Cytb最好,其次是ND5、16SrRNA、ND2和ATP6,ATP8、ND4L和12SrRNA较差,其余基因表现中等。mtDNA中不同基因在进化过程中受到的选择压力不同,进化速率也不同,导致各基因所包含的系统发育信息有所差异[18]。黄小林等[11]、张丽丽等[13]、郭昱嵩等[25]、陈姝君等[24]和Zardoya等[23]分别基于石斑鱼属、鳀科、鲈形目、硬骨鱼类和脊椎动物等不同分类阶元评估线粒体蛋白质编码基因在系统发育分析时的适用性。本文在总体上与前人结论一致,但在个别基因上存在差异,作者认为这与研究对象和分类阶元不同有关,存在一定的物种和阶元差异性。通过与同阶元及不同阶元的比较揭示了篮子鱼属mtDNA中不同基因进化的规律,为后续篮子鱼的系统分类研究和分子标记的选取提供参考和依据。
