延河流域6种草本植物功能性状变异来源分析
2018-10-12曾鸿文温仲明朱朵菊
曾鸿文,温仲明,陶 宇,张 静,朱朵菊
(1.西北农林科技大学 林学院,陕西 杨凌 712100; 2.西北农林科技大学 水土保持研究所,陕西 杨凌 712100; 3.中国科学院 教育部水土保持与生态环境研究中心,陕西 杨凌 712100)
植物在漫长的进化和发育过程中会形成具有特定内部生理和外部形态特征的环境适应策略,这些影响生态系统的结构与功能的形态、生理或物候属性,称为植物功能性状[1-3]。它是植物对环境变化的响应作用机制[4-6],是植物与环境之间的联系桥梁,决定植物能否在这个环境中存活生长[7]。
如当植物生长在干旱贫瘠的环境时,往往通过降低比叶面积、增大叶厚度、增大叶组织密度等方式,减缓代谢速率[8]。Reich等[9]在全球尺度上采集了452个地区的1 280种植物叶片,比较其氮磷含量和经纬度温度间的关系,得出靠近越热带的地区,温度上升,叶片氮磷含量下降,而氮磷比升高;闫建成对内蒙古草原和荒漠区4种一年生植物进行研究,猪毛菜(Salsolacollina)和雾冰藜(Bassiadasyphylla)在干旱胁迫下,调整物候期的长短,减少结果量,加速种子成熟以响应降水量不足,比虎尾草(Chlorisvirgata)和冠芒草(Enneapogonbrachystachyum)表现出更强的耐旱能力[10]。水分和温度是影响植物生长的重要因素,而地形变化会对温度、光照、降水等资源具有再分配作用,微地形会导致小范围气候异质性,进而影响植物性状的表达[11]。
植物功能性状不仅受到环境因素影响,也受到遗传背景因素的限制。在长期的进化过程中,植物物种分化,形成了不同的性状特征与环境适应策略。Givnish[12]研究发现亲缘物种在不同生境下依然表现出相似的物候特征和性状特征,认为植物的适应策略和方式受系统发育背景所限制,而又会在环境胁迫的作用下发生适应和进化。宋彦涛[13]对松嫩草地80种草本植物氮磷含量进行统计,发现豆科植物通过与其共生的根瘤菌固定大气中的氮,从而有较其他科更高的叶氮含量和叶片氮磷比。
在物种进化分化的过程中,同科物种具有相近的亲缘关系,从而拥有相似的性状特性。研究显示,遗传背景因素是植物性状变异主要来源之一,使得植物性状与环境变化间的关系研究存在一定的不确定性[14]。各物种因发育背景的不同,会对环境变化作出不同的应对策略。覃凤飞等对新疆3种牧草种的光合作用与水分利用效率进行比较[15],结果表明,在干旱缺水时,3种牧草种均能通过调节自身耗水来减少对有限水源的过度消耗,但3种物种应对的生态策略不同。因此,分别比较植物遗传背景和环境变化对植物功能性状的影响,有利于深入了解不同植物对环境变化的应对策略和反应机制。
本研究选择黄土丘陵区的延河流域为研究区,选择流域广泛分布的6种乡土草本植物为研究对象,包括阿尔泰狗娃花(Heteropappusaltaicus)、茭蒿(Artemisiagiraldii)、铁杆蒿(Artemisiasacrorum)3种菊科植物,及白羊草(Bothriochloaischcemum)、大针茅(Stipagrandis)、长芒草(Stipabungeana)3种禾本科植物,这些物种为延河流域大多数草本群落的优势种[16],对植被恢复具有重要意义。
本文试图探明不同遗传背景和环境变化对6种植物性状变异的影响,并分析植物功能性状与环境因子的关系,为黄土高原区植被恢复提供一定的理论依据。
1 研究地区与研究方法
1.1 研究区概况
延河是黄河中游河口镇-龙门区间的一级支流(108°45′—110°28′E,36°23′—37°17′N),位于陕西省北部,流域面积为7 687 km2,属大陆性半干旱季风气候带,年均降雨约500 mm,年均温约9 ℃。流域自东南向西北温度、降雨梯度变化明显,植被因气候条件亦呈明显的地带性分布,根据环境差异可依次分为森林带、森林草原带和草原带(表1)[17]。铁杆蒿、阿尔泰狗娃花、白羊草、茭蒿等草本植物在该流域各个植被带广泛分布,是流域草本群落的优势物种。
1.2 野外调查与样品采集
根据环境因子的梯度变化,同时充分考虑地形变化,在3个植被带内选择人类活动干扰、自然植被发育较好的无林地选取样点。此次野外调查共选取146个样点,其中草原带51个,森林草原带61个,森林带34个,样点分布见图1。数据采集时间为2010—2015年7月至8月。在样点设置1 m×1 m的取样样方,分别对样方内的阿尔泰狗娃花、铁杆蒿、茭蒿、长芒草、白羊草、大针茅6个物种进行调查取样,取样时选取生长良好、没有遮阴的植株个体。

表1 延河流域3个植被带的环境因子
对每个样地进行编号,记录样点的经纬度。在每个样方左上角、中部和右下角分别取3株长势良好的目标物种植株,每株分别沿东南西北4个方向采摘10片无病虫害且发育良好的叶片。用精度为0.01 mm的电子游标卡尺避开叶脉分别测量叶片前、中、末端的厚度,取其平均值作为叶片厚度。用铁铲挖取每株目标物种的根部,清除土壤等其他杂质,采集直径小于2 mm的细根。将叶片和细根分别编号后密封,保存至车载冰箱(避免由于叶片和细根的水分蒸发而造成的误差),带回实验室进行分析。用扫描仪将平展的叶片图像扫描至电脑,并用图像分析软件IPP(image pro-plus)计算叶片面积。细根清水洗净,用精度为0.01 mm的电子游标卡尺测量10条健康且直径小于2 mm的细根长度,为细根长;将洗净后的细根完全浸没在盛水的25 ml量筒中,10秒后测出量筒内体积的变化即为细根的体积。将叶片和细根样品编号后分别放入温度为80℃的烘箱内烘干,48 h后,用精度为万分之一的天平称取烘干后的样品的干质量。用粉碎机将烘干后样品粉碎,用H2SO4-HO2法消解,采用凯氏定氮仪法测定叶氮含量(LNC,mg/g)和根氮含量(RNC,mg/g),用钼锑抗比色法测定叶磷含量(LPC,mg/g)和根磷含量(RPC,mg/g)。每个样品的同一性状重复测定3次,取平均值为最终值。最后,根据测得的数据计算各个功能性状的值,比叶面积(SLA,mm2/mg)=叶片面积/叶片干质量;叶组织密度(LTD,mg/mm3)=叶片干质量/叶片体积;比根长(SRL,mm/mg)=总细根长/细根干质量;根组织密度(RTD,mg/mm)=细根干质量/细根体积。
图1 研究区样点分布
叶片是植物进行光合作用,制造养分的重要器官,可塑性强,受环境影响较大[18],其性状与植物对环境中资源的利用率密切相关,能够反映植物在适应环境变化过程中性状的改变与及作出的生存对策。细根作为植物吸收环境水分营养的器官,其性状特征承载一定的环境状况,能反映植物的耐胁迫能力,是植物是否能在新环境生长分布的决定性因素之一。本研究选取十个易于观测且对环境变化敏感、能够反映植物生存策略的叶性状与细根性状,包括:比叶面积(SLA)、叶组织密度(LTD)、比根长(SRL)、根组织密度(RTD)、叶氮含量(LNC)、叶磷含量(LPC)、叶氮磷比(LNP)、根氮含量(RNC)、根磷含量(RPC)、根氮磷比(RNP)。
1.3 环境数据的提取
根据黄土高原数字高程模型提取延河流域的地形因子专题图。收集黄土高原区域各气象观测站近30 a的温度、降雨和蒸发量数据,结合地形变化,采用ANUSPLN空间插值工具插值处理,提取出延河流域各气候因子专题图[19]。提取的7个环境因子包括:(1) 地形因子:坡度(Slope),坡向(Aspect),地形部位(Position),高程(Elevation);(2) 气候因子:年均降雨量(Rainaver),年均温(Temaver)、年均蒸发量(Et)。根据各样地的经纬度数据,从环境因子专题图中提取出相应的环境因子。
1.4 数据处理
对各样点植物功能性状分类用Microsoft Excel整理。在IBM SPSS Statistics 21软件对数据进行正态检验,采用单因素方差分析法(One-way ANOVA)分析3个不同植被带间,不同科种间表现出的性状差异;用Pearson相关系数分别检验各科各个物种性状间的相关性及性状与环境因子的相关性;选取植被区代表气候综合因子,并考虑地形因子(海拔、坡度、坡向、坡位)及遗传背景(用物种区分)的影响,不考虑各因素间的交互作用,将以上6个因素作为固定效应处理,通过方差分析,剖析各因素对各个植物性状的相对影响。以上所有数据分析均在Microsoft Excel和SPSS 21.0中完成。
2 结果与分析
2.1 环境因子对植物功能性状的影响
2.1.1 气候变化对植物功能性状的影响 植物功能性状在不同植被带(代表不同生物气候区)的变化表明,3个植被带间除了LTD外,其他功能性状差异均显著(表2)。其中,SLA和SRL在森林草原带最大,森林带次之,草原带最小。草原带的SLA和SRL均显著低于森林草原带,RTD却显著高于森林草原带。RTD,LPC和RPC在草原带最大,森林带次之,森林草原带最小。草原带的RTD和LPC均显著高于森林草原带,分别是森林草原带的1.795倍和2.144倍。草原带和森林带的RPC显著高于森林草原带,分别是其的2.442倍和1.886倍。LNC,RNC,LNP,RNP在3个植被带间表现出显著差异,且沿草原带,森林草原带至森林带递增。在3个植被带性状的两两对比中,LNC均有显著差异,草原带显著低于森林草原带,森林草原带显著低于森林带。森林带的RNC显著高于草原带和森林草原带。草原带的LNP和RNP显著低于森林草原带和森林带,且森林带和森林草原带的LNP分别是草原带的2.525倍和2.077倍;森林带和森林草原带的RNP分别是草原带的2.177倍和1.703倍。

表2 延河流域6种植物功能性状的空间分布
注:(1)同行不同字母表示差异显著(p<0.05)。
2.1.2 微地形变化对植物功能性状的影响 地形因子对环境资源具有再分配的作用。地形因子与植物功能性状的Pearson相关分析系数表示(表3),坡向与植物功能性状间无显著关系。
坡度与LNP呈显著正相关,与LPC,RPC呈显著负相关;坡位与RPC呈显著正相关,相关度为0.192;海拔与LTD,RTD,LPC呈显著正相关,与SLA,LNP呈显著负相关。
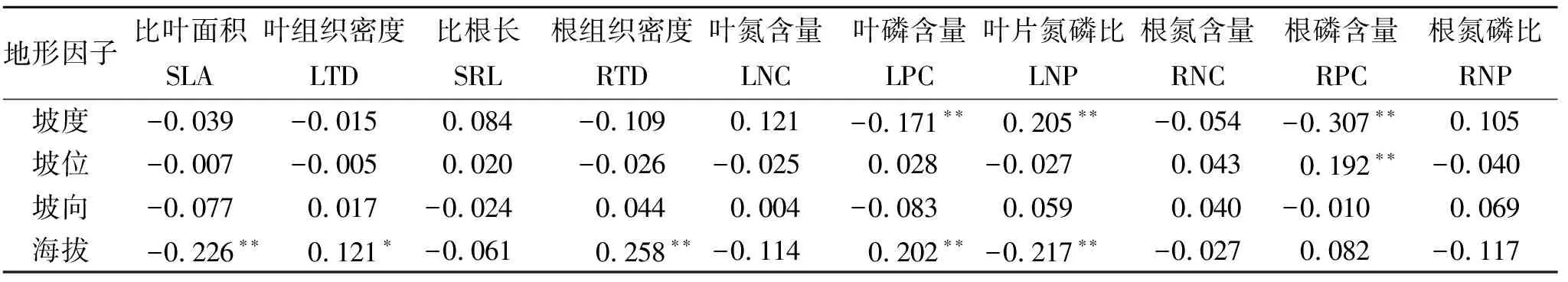
表3 地形因子与植物功能性状的Pearson相关系数
注:*p<0.05;**p<0.01;***p<0.001,下同。
2.2 遗传背景对植物功能性状的影响
植物遗传背景对植物性状的影响是植物经过长时间的自然发展进化而形成的,不容易受到外部环境的动摇[20]。菊科和禾本科间的植物功能性状差异较大(表4)。LTD,RTD,LNP,RPC在菊科和禾本科间差异不显著;菊科的SLA,LNC,LPC,RNC,RNP均显著高于禾本科;菊科的SRL显著低于禾本科。

表4 菊科和禾本科的功能性状
不仅不同科的植物功能性状有所差异,即使是同科内的不同物种功能性状也有较大差异。菊科植物阿尔泰狗娃花、茭蒿、铁杆蒿的比根长、根组织密度、叶氮含量、叶氮磷比、根氮含量差异显著。禾本科植物长芒草、白羊草和大针茅间,比叶面积、比根长、根组织密度、叶氮含量、叶氮磷比、根氮磷比、根氮含量、根氮磷比均有显著差异。
2.3 植物功能性状变异来源
上述结果表明,植物功能性状不仅受到环境因子的影响,且深受遗传背景的限制。为此,本文选取年均降雨量、年均温度和年均蒸发量代表气候因子,坡向、坡度和坡位代表地形因子,植物物种代表遗传背景,分析环境因子(包括气候因子和地形因子)和遗传背景对性状变异的影响(表5)。
结果表明,模型对性状变异解释的比例范围在25.3%~51.4%。其中,植物物种改变解释的比例占0.7%~17.23%,对SLA,SRL,LNC,LNP,RNC均有显著影响,解释比例分别为16.1%,10.7%,17.2%,9.0%,9.0%。年均降雨量的解释比例占0.6%~21.2%,对LTD,LPC,LNP,RPC,RNP均有显著影响,解释比例分别为3.3%,16.0%,9.8%,21.2%,4.9%。年均温度的解释比例占0.5%~12.9%,对SLA,SRL,RTD均有显著影响,解释比例分别为8.0%,5.5%,12.9%。年均蒸发量的解释比例占0.5%~12.4%,对RTD,LNC,LPC,LNP,RPC,RNP均有显著影响,解释比例分别为6.5%,4.5%,12.4%,10.8%,9.0%,5.6%。坡度的解释比例占0.6%~9.0%,对RTD,RPC均有显著影响,解释比例分别为8.9%,9.0%。坡位解释比例占0.6%~6.8%,对LPC,LNP均有显著影响,解释比例分别为5.2%,6.8%。坡向的解释比例占2.2%~7.7 %,对LTD,RTD,LPC,LNP均有显著影响,解释比例分别为7.2%,7.5%,7.6%,7.7%。海拔的解释比例占0.0%~4.6%,仅对SRL有显著影响,解释比例为4.6%。

表5 植物变异来源
注:df表示自由度;F表示方差;ss表示方差解释比例。
3 讨 论
3.1 植物功能性状差异来源
植物性状对环境变化的响应建立在系统发育史的基础上[21]。不同植物对环境的耐受性不同,应对环境资源短缺反应不一,应对环境变化的生存策略也不相同[22],有许多研究都反映出植物性状表达受遗传背景的限制[23]。植物适应环境变化所作出的生存策略并非一朝一夕而成,而是经过漫长的进化演变所得,植物在其极短的生活周期中可能会对为适应环境发生性状的改变,但终其一生,个体所作出的性状变异和这个物种千百年才形成的遗传性状相比是微乎其微的[24],因此植物性状对环境变化的响应是在原有遗传背景下进行的,受到遗传背景的制约。
本文对延河流域的6种植物的SLA,LTD,SRL,RTD,LNC,LPC,LNP,RNC,RPC,RNP共10个功能性状做了研究,表明,遗传背景对除了LTD和RTD外的8个功能性状都有显著影响,其中SRL,LNC,RNC这3个性状无论是在植物科间对比和科内性状比较都表现出显著差异,是性状变异的首要决定因子,与张莉[21]、寇萌[16]等的研究结果吻合。环境因子对植物性状变异也有一定影响,其中气候因子比地形因子的影响更大。
阿尔泰狗娃花、茭蒿、铁杆蒿、白羊草、大针茅、长芒草在延河流域内广泛分布,说明其在遗传背景下已经进行了相应的系统发育以适应环境变化。菊科植物的SLA,LNC,LPC,RNC,RNP均显著高于禾本科,能在水分和营养不足、土壤侵蚀强度高的环境中保持植株体内较高的氮含量,用以保持较高的光合作用效率;禾本科植物SRL显著高于菊科,其发达的根系有利于争夺土壤中的水分和营养。流域内的土壤磷含量较低,磷的可利用性较低,菊科和禾本科的RPC均没有表现出显著差异,但菊科的LPC显著高于禾本科,植物体内的磷主要来源于土壤,说明菊科植物的茎叶对磷元素有较强的积累能力和较高的利用效率。
3.2 植物性状对环境因子的响应
环境通过限制植物的性状范围来筛选物种,所以环境与植物性状的表达有明确的相关性[25-27]。把禾本科和菊科的10个性状分别汇总后与7个环境因素进行回归模型拟合,得出的模型均达不到统计学显著水平。这可能是跨物种分析导致的,植物对生存环境的要求不同,适应环境变化的生存策略也不同而导致的,但这也能看出同一科内的不同物种会对环境变化作出适合其生长繁殖的策略。
以菊科的LTD这一植物性状对主要变异来源年均降雨量的响应为例。菊科植物的LTD中阿尔泰狗娃花和茭蒿与年均降雨量无显著关系,而铁杆蒿则表现出显著正相关,相关性为-0.234(p<0.05),这说明铁杆蒿对干旱缺水比同科内的阿尔泰狗娃花和茭蒿敏感。铁杆蒿的LTD与LPC,RPC表现出显著正相关,相关性分别为0.306(p<0.01),0.636(p<0.05);与SLA,LNC,RNC表现出显著的负相关,相关性分别为-0.309(p<0.01)、-0.358(p<0.01)、-0.230(p<0.05)。在干旱环境下,铁杆蒿会把体内大部分的合成物质用于构建保护组织,以应对干旱胁迫。植物叶片水分不足时,LTD增大,SLA变小,植物体内的LNC和RNC变少,LPC和RPC增多,高组织密度的叶片植物和较低的氮磷比会使生物体周转生长速度减缓,提高抗旱能力。
4 结 论
本文分析的8个植物性状变异来源,依据各因素对性状的解析比例及各因子与性状的Pearson相关系数,得出如下影响排名:物种>年均降雨量>年均蒸发量>年均温度>坡度>坡位>海拔>坡向。
植物性状的表达主要还是受到遗传背景的限制[28],并在环境的影响下在植物的可接受范围波动[29-30]。在禾本科和菊科两者比较中发现植物性状的差异较大,对科内物种间的性状进行比较发现同一科内不同物种间的亦存在性状差异。
在环境变化时,植物通过调节性状的表达以适应环境变化,根据植物自身不同遗传背景和环境耐受性,产生各自的生存策略。
植物在漫长的进化和发育过程中,也会通过自身不同功能性状之间的协同进化,形成具有特定内部生理和外部形态特征的环境适应策略,以逐渐使植物适应环境生活,扩大植物原有的适生区域,这有助于环境生物多样性的提高,促进环境的稳定。
作为植物生长的基质,土壤向植物的根部提供水分养分和无机盐,对植物的生长、繁殖有重要影响,本研究未就土壤因子对植物功能性状的影响进行研究,或许会忽略土壤环境对植物功能性状的限制影响。
