牧压梯度下高寒草甸土壤容重及持水能力的变化特征
2018-10-12祝景彬贺慧丹李红琴杨永胜未亚西李英年
祝景彬,贺慧丹,李红琴,杨永胜,未亚西,罗 谨,李英年
(1.中国科学院 西北高原生物研究所,西宁 810001; 2.中国科学院大学,北京 100049; 3.中国科学院 高原生物适应与进化重点试验室,西宁 810001)
土壤持水能力反映出水资源在时空上的再分配状况[1],很大程度上受到土壤机械组成、有机质和植物地下生物量的直接或间接影响[2-4]。在生态系统中,水源涵养既是重要的服务功能,也具有不可替代的调节作用[5]。此外,土壤水分运动及含水量高低既是生态系统物质循环和能量流动的基础,又是控制各种营养物质矿化、稳定和转移的必要条件[6-7],在植物生长过程和繁殖策略等功能中具有不可替代的作用[8]。而土壤持水能力是评价陆地生态系统水源涵养的主要指标之一[9]。
青藏高原被誉为“中华水塔”,湖泊、冰川众多,是众多大江大河的发源地。同时,高寒草地植被土壤根系发达,具有较高的持水和滞水能力,对中国东部区域的水资源具有重要的调节和补给作用。近几十年来,由于全球气候变暖和人类活动等因素的影响,导致了高寒草甸大面积退化,青藏高原高寒草地土壤侵蚀趋于严重、水土流失加剧,致使高寒草甸植被的土壤持水能力下降。已有研究表明,退化的高寒草地除植被状况发生明显变化外[10],其土壤理化性质也产生显著的改变[11-12],进而影响到其水源涵养功能[13]。同时研究表明,土壤水分含量将影响到土壤碳的储存,土壤水分的减少可提高其通透性,导致土壤微生物活性和土壤呼吸速率增强,有机质分解速率加快[14]。还有研究表明,围栏封育或轻度放牧有利于提高退化草地的生产力[15],增加植被覆盖度及枯落物,枯草层厚度的增加不仅截留外来水分补偿能力,也减缓了土壤水蒸发能力[16],还可影响到植物地上地下生物量对土壤有机质的补给[17],进而会增加土壤有机质及营养成分含量[18]。而土壤有机质能改变土壤结构、土壤容重、紧实度等,终究影响到土壤含水量和持水能力[19-21]。然而,对高寒草甸的研究多集中在退化原因、演替过程、土壤养分以及恢复措施等支持功能方面,较少涉及其水循环和土壤水固持等水调节功能方面,更少有对放牧和封育草地的水分利用及水涵养方面的研究。为此,对牧压梯度下的高寒草甸土壤水分特征、持水能力的研究,将对于揭示高寒草甸水源涵养、保护寒区生态环境具有重要意义。
1 材料与方法
1.1 试验地概况
试验在青海海北高寒草甸生态系统国家野外科学观测研究站(海北站)进行。海北站地处青藏高原东北隅祁连山北支冷龙岭东段南麓坡地(37°29′—37°45′N,101°12′—101°23′E,海拔3 200~3 600 m)。该区位于亚洲大陆腹地,具有明显的高原大陆性气候,东南季风微弱。冬半年受西伯利亚冷高压控制,气候寒冷、干燥。夏半年受微弱的东南季风影响,气候凉爽、湿润。高海拔条件制约,气温极低,无明显四季之分,仅冷暖季之别,干湿季分明。年平均气温-1.7℃,降水量约560 mm,降水主要集中于植物生长季的5—9月,约占年降水量的80%,年平均日照时数2 462.7 h[22]。植被建群种为矮嵩草(Kobresiahumilis),主要优势种为垂穗披碱草(Elymusnutans)、异针茅(Stipaaliena)、麻花艽(Gentianastraminea)、甘肃棘豆(Oxytropiskansuensis)和紫羊茅(Festucarubra)等[23]。牧草生长低矮,群落结构简单,植被生长缓慢等[24]。土壤为草毡寒冻雏形土,具有发育年轻、土层浅薄、粗骨性强、有机质含量丰富等特征[25]。
1.2 研究材料及分析方法
1.2.1 试验设计 冬季放牧强度试验平台于2011年8月建立,参照以往研究放牧强度的经验[10],牧压梯度设置为封育对照(禁牧,CK)、轻度放牧(LG,4.5只羊/hm2)、中度放牧(MG,7.5只羊/hm2)和重度放牧(HG,15只羊/hm2)4种管理方式。试验地用围栏围封,试验羊为当地藏系绵羊。放牧时间为9月20日到次年5月31日。
1.2.2 要素监测与分析 监测与样品收集:2013年9月初开展了植被生物量、土壤样品的监测与收集。监测时以每个试验区中央点为中心,设计40 m×40 m的观测区(试验地远大于40 m×40 m),这样以中央点及40 m×40 m的角点附近为观测样点,即每个试验区共5个点(重复)。9月初依4个点正北方旋转270°后外延2~3 m处进行了样方为50 cm×50 cm的生物量、种类组成等监测。先用卷尺测定了植物群落的平均高度(5个重复的平均),用目测法记录总盖度,然后用卷尺测定分种高度,用目测测定分种盖度,而后收集枯落物装袋,用手刮的方式收集地表由枯落物、牛羊粪多年形成且并未分解留存于地表的半腐殖质量装袋,用剪刀齐地面分种剪下绿色植物个体装袋;再用内径8 cm的根钻分0—10 cm,10—20 cm和20—40 cm层次收集土柱(每个样方3个重复),以测定地下根系生物量;最后在该样方区挖掘剖面,分0—10 cm,10—20 cm和20—40 cm层次收集适量土壤样品装袋,并收集环刀土壤样品,环刀(容积为100 cm3)土壤样品现场测量重量后封盖,备测容重和持水量等参数。
土壤容重:用环刀烘干重计算得到(g/cm3)。
土壤有机质:所取土壤样品带回室内自然阴干,利用重铬酸钾容量法—外加热法测定土壤有机质的含量。
土壤饱和含水量[26]:将装有湿土的环刀(环刀质量为m)揭去上下底盖,仅留垫有滤纸带孔底盖,放入平底盆或其他容器内,注入并保持盆中水层高度至环刀上沿为止,使其吸水达12 h(质地粘重的土壤浸泡时间可稍长),此时环刀土壤中所有孔隙都充满了水,盖上上、下底盖,水平取出,用干毛巾擦掉环刀外沾的水,立即称量质量(m1)即可算出土壤饱和持水量(%,mm)。
土壤毛管持水量:将上述称量质量(m1)后的环刀,仅留垫滤纸的带网眼的底盖,放置在铺有干砂的平底盘中2 h,此时环刀中土壤的非毛管水分已全部流出,但毛细管中仍充满水分。盖上底盖后立即称量质量(m2),即可计算出毛管持水量(%,mm)。
土壤田间持水量:将上述称量质量(m2)后的环刀,如前一样继续放置在铺有干沙的平底盘中,保持一定时间(沙土1昼夜,壤土2~3昼夜,黏土4~5昼夜),此时环刀中土壤的水分为毛管悬着水。盖上上下底盖,立即称量质量(m3)后计算得到。
1.2.3 数据处理 数据处理前采用Kolmogorov-Smirnov法进行正态性检验,当数据不满足正态分布则采用标准化后进行比较。对所测定的数据利用Excel和SPSS 16.0软件对数据进行统计分析,单因子方差分析(ANOVA)和新复极差法(SSR)用于牧压梯度下各指标间的比较和差异显著性检验(α=0.05)。
2 结果与分析
2.1 牧压梯度下高寒草甸土壤容重的变化特征
图1表明,0—10 cm土壤剖面上LG与CK,HG处理下土壤容重具有显著性差异,10—20 cm剖面上CK与LG存在显著性差异,20—40 cm剖面上各试验组之间均不具有显著差异,由此说明短期放牧对于土壤容重的影响主要体现在表层,对于深层土壤的影响并不显著。同时也可看出,在不同层次的土壤容重都表现为CK最大,这可能与样地的选择有关,具体原因有待进一步分析;但是,不同牧压梯度下同一层次的土壤容重都表现为HG最大,LG,MG的容重相对较小,说明适度放牧使土壤容重减小,而放牧超过一定强度后使土壤容重增加。此外,土壤容重随着土层的加深逐渐增大,二者呈现明显的正相关关系。
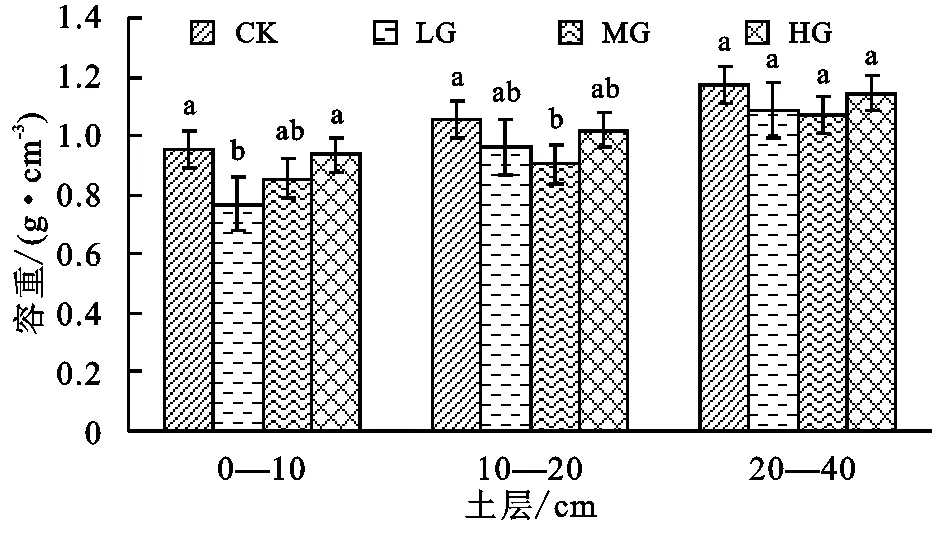
图1 牧压梯度下高寒草甸土壤容重的变化特征
2.2 牧压梯度下高寒草甸土壤持水能力的变化特征
表1可见,CK与LG的饱和持水量在0—10 cm具有显著差异,CK与MG的饱和持水量在10—20 cm具有显著差异,不同土层的各试验组毛管持水量之间不具有显著差异,而只有CK与LG的田间持水量在0—10 cm土层具有显著差异,表明短期放牧对于土壤持水能力的影响主要体现在表层,对于深层土壤持水能力的影响并不明显。此外,不同土层的饱和持水量、毛管持水量和田间持水量整体上表现为LG,MG相对较大,而CK,HG相对较小,所以适度放牧有利于提高土壤的持水能力,而禁牧和重度放牧都不利于土壤持水能力的提高。另外,也可看出土壤的持水能力随着深度的增加逐渐下降,二者呈明显的负相关关系。
2.3 牧压梯度下高寒草甸土壤持水能力的影响因素分析
图2A可见,不同牧压梯度下的地下生物量都表现为随着深度的增加而减小,CK,LG,MG,HG在0—10 cm的地下生物量分别占其在0—40 cm地下生物量的85.12%,78.06%,84.73%,79.91%。此外,在0—10 cm的地下生物量表现为HG>CK>LG>MG,在其他层次均表现为HG>LG>CK>MG。图2B可见,不同牧压梯度下的土壤有机质含量都表现为随着土层深度的增加而减小。此外,在不同土层的有机质含量都表现为LG>CK>MG>HG,说明在生长季末期(9月初)适度放牧会增加土壤有机质含量,而过度放牧则使土壤中的有机质含量减少。
表2 可见,饱和持水量、毛管持水量和田间持水量三者之间表现出极显著正相关关系(p<0.01),三者对于不同牧压梯度表现出一致的响应;土壤容重与持水能力表现出极显著负相关关系;地下生物量、有机质和土壤持水能力的相关性虽然不显著(p<0.1),但仍能说明地下生物量、土壤有机质同土壤持水能力呈正相关关系。

表1 牧压梯度下高寒草甸土壤持水能力的变化特征

图2 牧压梯高寒草甸植被地下生物量、土壤有机质的变化特征

表2 牧压梯度下高寒草甸土壤持水能力与各因素的相关性
3 讨 论
土地利用方式对土壤的结构功能和理化性质有直接的影响,研究表明合理的土地利用方式有利于土壤的改善,不合理的土地利用方式会导致土壤质量的下降[27]。土壤持水能力是土壤重要的物理性质,也是评价土壤水分调节功能和水源涵养的重要指标。
土壤容重是指单位体积原状土壤的重量,对于土壤的物理结构和化学性质都有重要影响,而且对植被根系的生长和营养物质的转移也有重要影响[28]。有些研究认为,封育可以使土壤容重减小[29],草地退化会使土壤容重显著增加[30],而本研究中封育土壤容重相对较大,这是由于不同区域的降水、温度、地形等一系列因素的差异,使之对于处理方式具有不同的响应。土壤容重的改变是一个相对缓慢的过程,由于放牧年限较短,所以放牧对于土壤容重的影响主要体现在表层,对于深层土壤容重的影响仍需一段时间才能有所体现。整体来讲,适度放牧可以使土壤容重减小,过度放牧会使土壤容重增加,这是因为牲畜的适度踩踏促进了枯落物和表层土壤的融合,加之牲畜排泄的粪便,一定程度上加快了地上生物量向土壤的归还,使土壤表层的有机质含量增加,有利于表层土壤的改善;但是,当放牧超过一定强度后,由于牲畜的过分踩踏,会使得土壤变得硬实,最终使土壤容重增加。
放牧对于群落地上生物量的影响是即时发生的,而对于地下生物量的影响有一定的滞后效应,植被受到环境的干扰胁迫后,生物量地上地下生物量的分配会发生改变[31]。重牧条件下的植被由于受到牲畜相对严重的啃食和踩踏,其光合作用制造的有机物更倾向于往地下分配,加之受高海拔条件的制约,研究区冬半年漫长而寒冷,致使根系在低温冻结期不易矿化,即使在夏半年,也因地温不高而分解较弱,大部分死根长期保留在近地表层,因而HG的地下生物量相对较高。土壤有机质含量的动态变化主要取决于植物凋落物的归还量和枯枝落叶的分解速率,以及土壤生物的呼吸速率、表层土壤的流失量,渗水作用下在土体中的淋溶、迁移、淀积及其在土壤小动物作用下与矿质土体振动、混合的过程。适度放牧对于植被的踩踏和啃食有利于地上地下生物量向土壤的归还,使得生长季末期(9月初)土壤有机质含量增多;而过度放牧由于啃食严重,使生物量往土壤的归还量减少,最终使得生长季末期(9月初)土壤有机质减少。
土壤容重和孔隙度是土壤物理性质的重要参数,直接影响土壤水源涵养能力的高低[32]。毛管持水量能反映土壤毛管孔隙度的大小[27],由表1可知适度放牧可以增加土壤的毛管孔隙度,过度放牧和完全禁牧都不利于提高土壤毛管孔隙度,并且由表2可知毛管孔隙度与土壤持水能力呈极显著正相关。土壤容重较大时,土质相对硬实,毛管孔隙度较小,土壤持水能力较低;土壤容重较小时,土质相对比较疏松,具有较大的毛管孔隙度,土壤持水能力较强,容重与土壤持水能力呈极显著负相关,这与吴启华[33]的研究结果一致。地下生物量和土壤有机质有利于改善土壤物理性质,使土壤变得疏松多孔,容重降低,使土壤持水能力提高,因此地下生物量、有机质与土壤持水能力呈正相关。
土地利用方式对土壤的容重、孔隙度、地下生物量、有机质含量等一系列要素有明显的影响,而这些要素直接影响土壤的持水能力,所以不同放牧梯度通过对于土壤这些要素的不同影响而改变了土壤的水源涵养能力。总体来说,适度放牧有利于改善土壤,降低容重,增加毛管孔隙度,使土壤有机质含量增加,最终使得土壤的持水能力提高。这一研究结果表明,在高寒草甸地区过度放牧会使土壤水源涵养能力下降,而完全禁牧也不利于土壤水源涵养能力的提高,只有适度放牧才有利于提高水源涵养能力,才有利于生态环境的保护。但是,本研究只是探讨了牧压梯度下高寒草甸土壤持水能力的变化特征,而土壤的实际贮水量不仅受到土壤持水能力的影响,同时也受到植被生长状况、地表覆盖物等因素影响,牧压对土壤中实际贮水量和水分有效利用率的影响仍待进一步研究。
4 结 论
(1) 短期放牧对土壤容重的影响主要体现在表层,对于深层土壤的影响并不显著;土壤容重随着土层深度的加深逐渐增大;适度放牧会使土壤容重减小,而放牧强度超过一定强度后会使土壤容重增加。
(2) 牧压梯度下高寒草甸在不同土层的饱和持水量、毛管持水量和田间持水量整体上表现为LG,MG相对较大,而CK,HG相对较小,所以适度放牧有利于提高土壤的持水能力。短期放牧对于土壤持水能力的影响主要体现在表层,且土壤的持水能力随着深度的增加逐渐下降。
(3) 土壤的持水能力受多种因素共同作用,主要受土壤容重、有机质、地下生物量的影响,与土壤容重呈极显著的负相关(p<0.01),与土壤有机质、地下生物量呈弱的正相关关系。
