5-氮杂-2′-脱氧胞苷对小鼠海马神经元细胞系HT22细胞DNA甲基转移酶表达和细胞周期的影响❋
2018-10-11田小莉闫少春时静华
田小莉 杨 洁 杨 静 闫少春 时静华# 邵 国
(1 包头医学院中心实验室生物医学研究中心, 神经科学研究所, 包头 014010; 2 内蒙古自治区低氧转化医学重点实验室, 包头 014010; 3 首都医科大学宣武医院低氧适应转化医学北京重点实验室, 北京 100053)
表观遗传如DNA甲基化在哺乳动物生长发育过程中起着重要的作用。DNA甲基转移酶(DNMTs)是调控表观遗传DNA甲基化基因表达的关键酶,它催化甲基使其从S-腺苷-甲硫氨酸共价转移到CpG岛胞嘧啶残基上[1]。哺乳动物体中,有3类DNA甲基化转移酶:DNMT1、DNMT3A和DNMT3B,在基因组甲基化过程中起着不同的作用。DNMT1是维持基因组中甲基化状态所必需的。DNMT3A和DNMT3B负责在胚胎发育期间或有丝分裂后的神经元中将非甲基化的DNA甲基化[2-3]。5-氮杂-2′-脱氧胞苷(5-Aza-CdR)是DNMTs抑制剂,是一种胞嘧啶类似物,其通过与酶的半胱氨酸残基共价结合阻断DNMTs功能[4]。很多癌细胞系的研究发现5-Aza-CdR的修饰可以作为一种有效的药物来干扰癌症发展[5-7]。Xiong等[8]报道,5-Aza-CdR 通过改变表观遗传状态诱导人结直肠癌细胞的细胞周期停滞和细胞死亡。本研究主要是观察5-Aza-CdR 对HT22细胞周期、凋亡及DNMTs表达水平的影响。
1 材料和方法
1.1 实验细胞
HT22细胞系是小鼠海马神经元细胞系,来自于HT4。本文所用细胞由中国医学科学院北京协和医学院癌症研究所和肿瘤分子生物学国家实验室提供。HT22细胞中加入含有胎牛血清(FBS)、青霉素和链霉素的DMEM高糖培养基(Sigma-Aldrich,USA),置于37℃、5% CO2潮湿的细胞培养箱中培养[9],待细胞长至80%~90%且细胞状态良好,传代并开始实验。
1.2 5-Aza-CdR浓度筛选
为确定5-Aza-CdR的有效浓度,在HT22细胞中加入5、10、15、20mol/L和40μmol/L不同浓度的5-Aza-CdR,使用IncuCyte FLR在显微镜(4×)下对每孔细胞进行实时成像,观测细胞的增殖。预实验显示,20μmol/L 浓度的5-Aza-CdR对细胞增殖抑制作用最大,5μmol/L浓度的5-Aza-CdR对细胞增殖抑制作用最小。因此,我们选择将HT22细胞在5μmol/L(最低)和20μmol/L(最高)浓度的5-Aza-CdR中孵育24h。对照组细胞中加入PBS孵育24h。
1.3 IncuCyte成像分析
96孔细胞培养皿中,每孔加入100μl含有10% FBS的DMEM高葡萄糖培养基,每孔接种4×103个细胞。待细胞生长24h后加入不同浓度的5-Aza-CdR,加药后实时观察细胞68h。使用IncuCyte Confluence 1.5版软件(ESSEN Bioscience, Ann Arbor, MI, USA)分析数据。IncuCyte实验重复3次。
1.4 Real-time PCR分析
使用TRIzol试剂从HT22细胞中提取总RNA,根据反转录试剂盒(cDNA Synthesis Kit(Invitrogen Company, USA))说明书将总RNA逆转录成cDNA,在ABI 7900PCR仪上进行Real-time PCR,使用2△△CT方法,以β-actin做为内参[10]。每个样品做3个复孔,重复实验6次。表1是由Sangon生物技术公司(中国上海)合成的Real-time PCR引物。
1.5 免疫印迹检测
使用(RIPA)细胞裂解液(碧云天,中国上海)提取蛋白,用BCA蛋白测定试剂盒(Pierce Biotechnology, Rockford, USA)测定蛋白浓度,然后用5%的SDS-PAGE浓缩胶和10% SDS-PAGE分离胶电泳分离等量的蛋白样品,转移至PVDF膜上,5%脱脂奶粉封闭1h,鼠源DNMT1、DNMT3A、DNMT3B多克隆抗体(Novus Biologicals, Littleton, CO, USA)、鼠源β-actin 单克隆抗体4℃孵育14h(Sigma, St.Louis, MO, USA),洗膜3次,孵育山羊抗鼠二抗(1∶1000),室温2h。加ECL(碧云天,中国上海)发光液,放入蛋白凝胶图像分析系统中扫描,使用Image J(Scion Corporation, Torrance, CA, USA)图像分析系统来量化目的条带的光密度。蛋白印迹数据以β-actin做内参。
1.6 细胞周期检测
HT22细胞弃掉培养基,PBS洗涤2次,胰蛋白酶消化20~30s后,将细胞收集于离心管中,1500r/min 4℃离心8min,预冷的PBS洗细胞2次,缓慢加入预冷的70%乙醇,4℃固定过夜[11]。弃掉固定液,PBS冲洗细胞,加入适量PI(碘化丙啶)染色,避光4℃反应30min。流式细胞仪检测,对G1期、S期细胞进行计数。
1.7 细胞凋亡检测
如上方法收集细胞悬液,加入400μl 1×Binding Buffer悬浮细胞,按试剂盒说明书加入Annexin V-FITC和碘化丙啶(PI)进行染色,轻轻混匀,避光4℃反应30min。流式细胞仪检测。
1.8 DNMTs活性测定
收集的HT22细胞,1500r/min离心5min,弃去上清液。将收集的HT22细胞用EpiQuik核提取试剂盒(Epigentek, Brooklyn, NY, USA)提取核蛋白,用EpiQuik DNMT活性/抑制测定Ultra试剂盒(Epigentek Brooklyn, NY, USA)检测总DNMTs,DNMT1和DNMT3A的活性,对样品进行DNA甲基转移酶活性分析。
1.9 统计学处理

2 结果
2.1 5-Aza-CdR对HT22细胞形态的影响
不同浓度5-Aza-CdR对HT22细胞形态的影响如图2所示。光学显微镜下观察HT22细胞显示,对照组(图1A)中HT22细胞数量较多,细胞大且亮,折射率好,细胞膜边界清晰。与对照组相比,5-Aza-CdR处理组中HT22细胞数量明显减少,细胞膜边界粗糙且附着颗粒,出现胞质空泡化(图1B、C)。
2.2 5-Aza-CdR对HT22细胞周期和细胞凋亡的影响
结果表明,加入不同浓度5-Aza-CdR的HT22细胞,HT22细胞在S期的数目显著增加,但在G1期细胞数目减少(图2A),表明细胞在S期阻滞。此外,5-Aza-CdR抑制早期凋亡,但促进晚期凋亡(图2B、C)。

图1 光学显微镜下HT22细胞形态,×40。A: 对照组;B: 5μmol/L 5-Aza-CdR加药组;C: 20μmol/L 5-Aza-CdR加药组;箭头: 细胞质空泡.
图2 5-Aza-CdR对细胞周期和凋亡的影响。A: 细胞周期;a: S期;b: G1期。B: 流式细胞仪检测结果图;a: 对照组;b: 5μmol/L 5-Aza-CdR group; c: 20μmol/L 5-Aza-CdR group。C: 细胞凋亡;a: 早期凋亡;b: 晚期凋亡。*P<0.05.
Fig 1 Light microscopic examination of morphology of HT22 cells,×40. A: Control group; B: 5μmol/L 5-Aza-CdR group ; C: 20μmol/L 5-Aza-CdR group. Arrows depicted vacuolization.
Fig 2 Effects of 5-Aza-CdR on cell cycle progression. A: Cell cycle; a: S phase; b: G1 phase. B: Flow cytometric analysis results; a: Control group; b: 5μmol/L 5-Aza-CdR group; c: 20μmol/L 5-Aza-CdR group. C: Apoptosis; a: Early apoptosis; b: Late apoptosis.*P<0.05.
2.3 5-Aza-CdR对DNMT1、DNMT3A、DNMT3B mRNA和蛋白表达的影响
Real-time PCR结果显示,与对照组相比,DNMT1和DNMT3A mRNA表达水平显著降低,但DNMT3B mRNA表达水平没有明显变化(图3A)。DNMT1和3A蛋白表达水平显著降低,与mRNA表达水平相一致(图3B),表明5-Aza-CdR对DNMT1和DNMT3A 的mRNA和蛋白表达起抑制作用。
2.4 DNMTs活性检测
结果显示,在2种5-Aza-CdR浓度下,DNMT1和DNMT3B活性显著降低(图4),然而,总DNMTs活性未发生明显变化。

图3 5-Aza-CdR对DNMT1、DNMT3A和DNMT3B mRNA(A)和蛋白(B)表达水平的影响
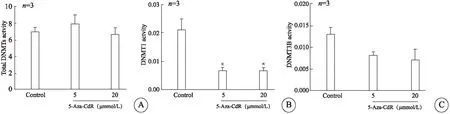
图4 5-Aza-CdR调节DNMTs活性的检测结果
3 讨论
癌症是严重威胁人类健康和生命且发病率仅次于心血管疾病的重大疾病,受到人们广泛关注。研究显示,癌症的发生发展与基因组异常甲基化有关,如在肺癌发生发展过程中出现整个基因组低甲基化,使原癌基因活跃、基因组易变等,致使原癌基因过度表达及染色体稳定性降低,从而导致肺癌发生;某些基因启动子区CpG岛发生高甲基化[12-13],使抑癌基因、凋亡基因、细胞周期调节基因等转录受到抑制,表达减少或沉默,相应的功能减弱或丧失,从而引起癌症发生。
DNA甲基化是一种调控基因表达的机制,且甲基化和去甲基化有一定的可逆性。5-Aza-CdR作为一种甲基转移酶抑制剂,曾被报道其影响细胞增殖、周期及凋亡,如Chen等[14]报道了5-Aza-CdR处理人食管鳞状细胞癌(ESCC)TE-1细胞系,显示KLF4因子上调诱导细胞凋亡和细胞周期停滞在S期;Fang等[15]指出,2μmol/L的5-Aza-CdR处理Daudi细胞系24h后,细胞在S期被阻滞;本研究用5μmol/L或20μmol/L 5-Aza-CdR处理HT22细胞24h后,显示HT22细胞在S期阻滞,说明5-Aza-CdR可以阻滞细胞周期,使细胞不再继续分裂增殖。Christman[16]提出,5-Aza-CdR并入DNA骨架,导致DNMTs丧失活性;Pan等[17]报道了在胰腺癌细胞系(PANC-1)中,5-Aza-CdR 联合大黄素通过减少DNMT1和DNMT3A的表达,来增强抑癌基因p16、RASSF1A和ppENK 5-Aza-CdR 的去甲基化;Zhang等[18]报道,10μmol/L的 5-AzaCdR对NG108-15胶质母细胞瘤和神经母细胞瘤杂交细胞系产生完全细胞增殖停滞且降低DNMT1、DNMT3A和DNMT3B的表达;本研究结果表明5-Aza-CdR处理降低DNMT1和DNMT3A的表达以及DNMT1和DNMT3B活性,说明5-Aza-CdR可以有效降低DNMTs的表达及活性。
5-Aza-CdR可抑制细胞中DNA甲基转移酶活性及其表达,影响细胞周期及凋亡,可以为癌症治疗提供一种有效途径,且随着研究的不断深入,DNA甲基化与癌症之间关系的研究将为癌症早期诊断、评估癌症风险、制定治疗方案以及预后判断等方面提供新的启示。
