水分和CO2浓度对冬小麦气孔特征、气体交换参数和生物量的影响
2018-10-10武海霞郭丽丽郝立华王清涛程东娟彭正萍张茜茜李树彬郑云普
武海霞 郭丽丽 郝立华 张 浩 王清涛 程东娟 彭正萍 李 菲 张茜茜 李树彬 徐 明 郑云普,*
水分和CO2浓度对冬小麦气孔特征、气体交换参数和生物量的影响
武海霞1,**郭丽丽1,**郝立华1张 浩2王清涛3程东娟1彭正萍4李 菲1张茜茜1李树彬5徐 明6,7,*郑云普1,*
1河北工程大学水利水电学院, 河北邯郸 056038;2邯郸学院生命科学与技术学院, 河北邯郸 056005;3河北工程大学园林与生态工程学院, 河北邯郸 056038;4河北农业大学资源与环境科学学院, 河北保定 071001;5邯郸市漳滏河灌溉供水管理处, 河北邯郸 056002;6中国科学院地理科学与资源研究所生态系统网络观测与模拟重点实验室, 北京 100101;7中国科学院大学环境与资源学院, 北京 100039
利用可精准控制CO2浓度([CO2])的大型人工气候室, 研究了水分亏缺和[CO2]升高对冬小麦气孔特征、气体交换参数及生物量的影响。结果表明, 水分亏缺导致冬小麦气孔开度减小和气孔空间分布的规则程度降低, 提高[CO2]能够减缓水分亏缺时气孔开度和气孔分布规则程度的下降幅度。与充分灌溉相比, 不同水分亏缺条件下冬小麦的净光合速率、气孔导度和蒸腾速率均显著降低(<0.05), [CO2]仅可缓解轻度亏水对气体交换过程的影响, 该缓解能力随水分亏缺程度的加剧而降低。水分亏缺降低冬小麦生物量, 但[CO2]升高对水分亏缺时生物量产生的影响不显著(>0.05)。水分亏缺条件下, 冬小麦通过调整气孔开度和气孔空间分布格局改变叶片的气体交换效率, [CO2]升高对冬小麦产生的“施肥效应”受土壤水分条件的限制。
CO2浓度倍增; 水分亏缺; 冬小麦; 生物量; 气孔特征; 气体交换参数
冬小麦(L.)为一种广泛分布的C3物种, 也是最重要的粮食作物之一。我国华北平原是冬小麦的主栽区, 其种植面积占总耕地面积的40%[1], 水资源匮乏是限制该地区冬小麦产量的重要因素[2]。C3植物对大气CO2浓度([CO2])变化较敏感, 其生理生化过程、生物量和产量受高[CO2]的影响较大[3]。[CO2]升高可以增加C3作物的净光合反应速率, 降低气孔导度和蒸腾速率, 缓解干旱胁迫, 提高作物水分利用效率[4-5]; 还能抑制作物光呼吸和Rubisco加氧反应, 进而提高净光合反应速率[6]; 但也有研究认为, 充分灌溉条件下提高[CO2]明显增加冬小麦的生物量和产量, 而当水分亏缺时提高[CO2]并没有增加其净光合速率和产量[7]。然而, 以往的大多数试验主要关注[CO2]升高或水分胁迫单一条件对作物生理和生长特性的影响机理, 很少涉及两个因素的共同效应[8]。本研究利用人工气候箱, 从分析单个气孔形态特征和气孔空间分布格局入手, 试图深入剖析[CO2]升高和水分亏缺对冬小麦生物量和产量形成的生理机制, 为农田生态系统管理提供数据支撑和理论依据。
1 材料与方法
1.1 试验材料与处理
在河北工程大学农业水土资源综合管理与调控实验室内进行盆栽试验。盆栽容器为底部打孔的塑料水桶(高27 cm, 桶口面积531 cm2, 桶底面积380 cm2), 且桶内土壤水分饱和后能够由桶底顺畅渗出。采用0~20 cm耕层黄壤土与营养土(2∶1)的混合土壤, 土壤容重为1.58 g cm–3, 田间持水量为52%, 基础肥力为有机质5.26 g kg-1、全氮0.68 g kg-1、全磷0.57 g kg-1、有效磷20 mg kg-1、速效钾58 mg kg-1, 土壤阳离子代换量为15.50 cmol kg-1。选用华北地区普遍种植的冬小麦品种‘石麦15’, 2017年3月22日播种, 3月28日幼苗全部出土, 4月6日幼苗三叶一心时定苗至每盆24株(折合大田密度450万株hm-2), 生育期内无追肥。
采用8个准确控制CO2浓度(±10 µmol mol-1)的大型人工气候箱对冬小麦进行光照培养, 设定人工气候箱的温度为21℃/16℃ (昼/夜), 植株冠层的光照强度为1000 µmol m-2s-1, 光照时间为8:00–20:00, 相对湿度控制为60%~70%。设2个不同[CO2]处理, 即4个人工气候箱内为自然大气[CO2], 约400 µmol mol-1([CO2]); 另4个人工气候箱内[CO2]为800 µmol mol-1([CO2])。
将盆栽塑料桶内盛满混合营养土, 放入水盆中浸泡5 min, 以确保桶内盆栽土壤充分吸收水分, 测定各个塑料桶内土壤的体积含水量(SWC)[8], 使之达到60%左右, 且各桶差异不显著。设4个水分处理, 分别是充分灌溉(SWC 55%~60%)、轻度亏水(SWC 35%~40%)、中度亏水(SWC 25%~30%)和重度亏水(SWC 10%~15%), 每处理5个重复。
冬小麦出土30 d后(4月28日)进行[CO2]和水分处理, 处理90 d后取样测定各项指标。
1.2 印记法气孔取样及测量
随机取每株3片叶, 用无色透明的指甲油分别涂于远轴面和近轴面的中部, 采集气孔印记样品, 用于气孔特征参数的测量。将叶片印迹的载玻片于Leica DM2500显微镜(Germany)下观察, 随机选3个显微视野, 每个视野取4张照片, 共12张气孔显微照片, 再从中选取5张计算气孔密度。利用AutoCAD 2010软件分别测量气孔的长度、宽度、周长、面积以及形状指数[9]。
1.3 气孔的空间分布格局分析
随机选取4个光学显微照片分析水分和[CO2]对气孔分布的影响。假定每个气孔都是叶片表面上分布的单点, 气孔开口的最中间位置为该单点的位置。利用空间分布软件Arc GIS 10.0将显微照片于相同的坐标下进行数字化处理, 得到每个气孔的坐标值; 再运用空间统计分析方法Ripley’s K方程对表征气孔分布状况的点进行空间分析[9]。
1.4 气体交换参数测定
随机取每桶1株冬小麦, 利用Li-6400便携式光合测定系统(LI-COR, USA)测定旗叶的净光合速率(n)、气孔导度(s)和蒸腾速率()。与光合测定系统配套的2 cm × 3 cm标准气室可以独立控制光合光量子通量密度(PPFD)、[CO2]、叶片温度以及气室湿度, 测量时标准气室参数设定为叶室内PPFD 1000 μmol m−2s−1PAR, [CO2] 400 μmol mol-1, 叶片温度25℃, 气室相对湿度50%~60%。利用公式WUE =n/计算叶片尺度上的水分利用效率。
1.5 植株生物量测定
用剪刀将全部冬小麦植株地上部和地下部分开, 分别放入牛皮纸袋中, 置烘箱(80℃)烘烤48 h至恒重后称量。每处理5次重复。
1.6 统计分析
利用SPSS13.0 (Chicago, IL)软件进行单因素方差分析, 再用Duncan’s multiple range test比较不同处理间差异的显著性。
2 结果与分析
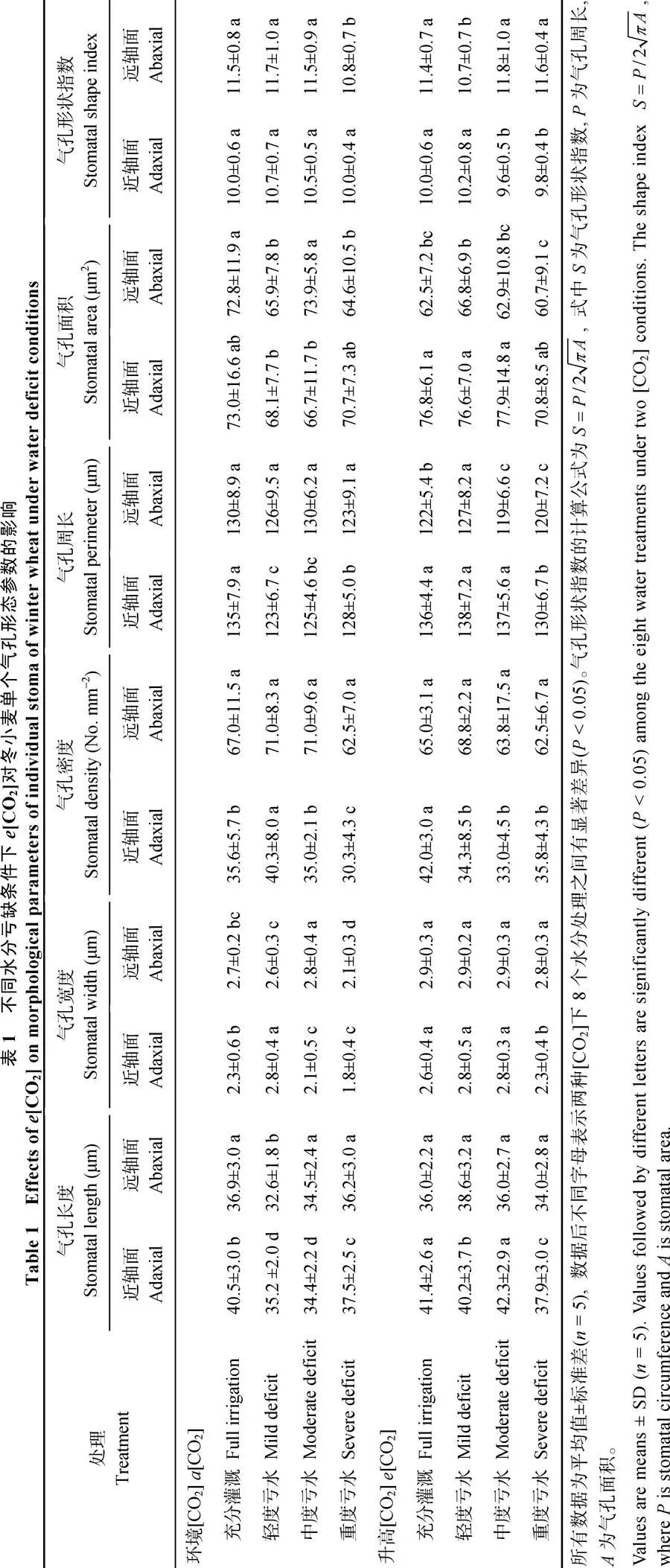
2.1 不同水分条件下e[CO2]对气孔参数的影响
[CO2]条件下近轴面的气孔长度和气孔周长均随土壤水分减少而显著降低, 但气孔密度和气孔宽度在轻度水分亏缺时达到最大值(表1)。水分亏缺导致远轴面的气孔宽度和气孔面积显著降低, 尤其是重度亏缺下的气孔宽度和气孔面积相比充分灌溉分别降低22%和11% (表1)。[CO2]导致水分亏缺时近轴面和远轴面气孔宽度的显著增大(表1)。然而, 轻度亏水条件下[CO2]增加叶片近轴面的气孔周长和气孔面积(表1)。
2.2 不同水分亏缺条件下e[CO2]对气孔空间分布格局的影响
尽管气孔最规则分布的空间尺度在冬小麦叶片近轴面(70 μm)和远轴面(100 μm)存在差异, 但不同亏水条件并没有对其产生明显影响(图1)。不同亏水条件时气孔分布格局由规则分布转变为随机分布的空间尺度在近轴面(120 μm)和远轴面(150 μm)之间也存在明显的差异, 轻度亏水时[CO2]导致叶片近轴面和远轴面气孔在较小空间尺度范围内的分布变得更加规则, 而中度和重度亏水的近轴面和远轴面气孔分布的规则程度在[CO2]条件下明显降低, 尤其中度和重度亏水条件下[CO2]处理的远轴面Lhat ()明显高于[CO2](图1)。

图1 不同水分条件下e[CO2]对冬小麦叶片近轴面(A)和远轴面(B)气孔空间分布格局的影响
深蓝色和褐色包迹线之间为气孔空间分布格局的95%置信区间。Lhat ()表示最小邻域距离, 当Lhat ()值小于95%置信区间时, 气孔在该尺度为规则分布, 且Lhat ()的最小值越小, 则气孔空间分布越规则。
The 95% confidence envelope for complete spatial randomness is between the dark blue and brown dotted lines. Lhat () value is the nearest neighbor distance, and stomata would follow a regular distribution at the scale when the Lhat () value is lower than the 95% boundary. The minimum Lhat () value is smaller, the spatial distribution pattern of stomata is more regular.
2.3 不同亏水条件下e[CO2]对气体交换过程的影响
不同程度水分亏缺显著降低[CO2]环境下的n、s和, 甚至仅轻度亏水就导致其分别降低85%、96%和82%。在[CO2]条件下, 水分亏缺也导致n、s和的下降, 尤其是重度亏水时分别下降99%、98%和94%。无论[CO2]还是[CO2]时, WUE均随水分亏缺的加剧而降低, 重度亏水时相比对照降低35% ([CO2])和83% ([CO2]), 且中度和重度亏水时[CO2]的WUE比[CO2]下降58%和76% (图2)。
2.4 不同亏水条件下e[CO2]对生物量的影响
充分灌溉条件下[CO2]分别提高冬小麦地上生物量、地下生物量和总生物量34%、99%和48%, 但不同水分亏缺时[CO2]对地上、地下和总生物量均没有产生显著影响。然而,[CO2]条件下水分亏缺导致冬小麦的地上、地下和总生物量均显著降低。另外, 同中度胁迫相比, 重度水分胁迫时[CO2]减少冬小麦的根冠比60% (图3)。
3 讨论
本研究中近轴面气孔形态参数随水分亏缺强度的增大而减小, 但远轴面气孔参数的变化并不明显, 表明远轴面气孔对水分亏缺的响应比近轴面更加敏感。[CO2]导致水分亏缺时近轴面的气孔面积增大, 但没有对气孔密度产生明显影响, 表明冬小麦在水分胁迫时主要通过调节叶片气孔开度响应提高的大气CO2浓度, 这可能是由于短期内植物对气孔开口大小的调控比改变气孔数量更高效。另外, 本研究还发现不同水分条件时气孔分布在小尺度范围内呈规则分布, 但气孔在远轴面上的分布比近轴面更加规则, 表明远轴面比近轴面具有更高的气体交换效率[9]。轻度水分亏缺条件下,[CO2]导致气孔在较小空间尺度范围内的分布变得更加规则, 但在中度或重度亏水条件下,[CO2]却使气孔空间分布的规则性降低, 表明土壤水分条件直接决定着植物气孔分布格局对[CO2]的响应状况。
水分亏缺对植物光合作用过程的抑制主要包括气孔限制和非气孔限制两种方式, 且随着亏水程度的增强, 气孔限制还会转变为非气孔限制[10-11]。本研究中, 轻度亏水显著降低n、s和, 表明轻度水分亏缺是由于s的降低而限制n, 但随着水分亏缺程度的逐渐增强, 影响光合作用过程的主要因素由气孔限制转变为非气孔限制, 故研究结果直接支持了前人相关研究所得结论, 即轻度亏水时主要是气孔因素限制光合作用过程, 但重度亏水时n的降低却是由于非气孔因素的限制。以往的研究发现,[CO2]增加n, 降低s和, 从而提高WUE[12-13]。本研究发现,[CO2]提高了轻度水分亏缺时的n, 但却显著降低中度和重度水分胁迫下的n, 表明提高CO2浓度仅可以缓解轻度水分亏缺对冬小麦带来的不利影响, 但随着水分胁迫的增强,[CO2]的“施肥”效应逐渐降低甚至完全消失。另外, 需要指出的是,[CO2]并没有提高冬小麦叶片的WUE, 甚至随着水分亏缺的增强而逐渐降低, 这与前人的研究结果恰恰相反[14]。产生上述现象的原因可能是由于水分亏缺导致冬小麦s和的降低, 而水分亏缺条件下[CO2]并不能提高n, 反而随着水分亏缺的增强,n下降幅度增大, 最终导致WUE的降低。

图2 不同水分条件下e[CO2]对冬小麦叶片气体交换参数的影响
A: 净光合反应速率; B: 气孔导度; C: 蒸腾速率; D: 水分利用效率。所有数据为平均值±标准差(= 5), 误差线上不同字母表示两种[CO2]下8个水分处理之间有显著差异(<0.05)。
A: net photosynthetic rate; B: stomatal conductance; C: transpiration rate; D: water use efficiency. Data are means ± SD (= 5). Different letters above the error bars indicate significantly different (< 0.05) among the eight water treatments under two [CO2] conditions.

(图3)
A: 地上生物量; B: 地下生物量; C: 总生物量; D: 根冠比。所有数据为平均值±标准差(=5)。误差线上不同字母表示两种[CO2]下8个水分处理之间有显著差异(< 0.05)。
A: aboveground biomass; B: belowground biomass; C: total biomass; D: root-to-shoot ratio. Data are means ±SD (= 5). Different letters above the error bars indicate significantly different (< 0.05) among the eight water treatments under two [CO2] conditions.
以往的研究发现作物在不同生育期遭受严重的水分胁迫将导致代谢速率减慢限制生长速度, 最终影响生物量的形成[15-16]。然而, 不同水分条件影响冬小麦生物量和粮食产量的潜在机制极其复杂, 尚未形成一致的结论[17-20]。本研究发现冬小麦的地上生物量和总生物量均随水分亏缺程度的加深呈显著下降趋势, 但地下生物量和根冠比并没有发生明显的变化, 表明水分亏缺条件下冬小麦可能将更多的光合产物分配给根系, 得以增加根系对同化物质的消耗, 确保根系从土壤中吸取更多的水分来维持地上部分生长, 这也可能是应对干旱胁迫形成的适应性策略。本研究结果发现[CO2]明显提高充分灌溉条件下的冬小麦地上生物量、地下生物量和总生物量, 而在水分亏缺条件下[CO2]并未对冬小麦生物量产生影响, 表明土壤水分条件在很大程度上决定着冬小麦的CO2“施肥效应”。本研究还表明,[CO2]对缓解冬小麦水分亏缺的效果不明显, 尽管有研究发现[CO2]能够有效降低干旱对植物生理生化过程及生长的影响程度[17]。这种差异可能主要是不同研究中水分胁迫程度和物种差异造成的。
[1] 张存杰, 王胜, 宋艳玲, 蔡雯悦. 我国北方地区冬小麦干旱灾害风险评估. 干旱气象, 2014, 32: 883–893 Zhang C J, Wang S, Song Y L, Cai W Y. Research of drought risk assessment for winter wheat in northern China., 2014, 32: 883–893 (in Chinese with English abstract)
[2] 唐星林, 曹永慧, 顾连宏, 周本智. 基于FvCB模型的叶片光合生理对环境因子的响应研究进展. 生态学报, 2017, 37: 6633–6645 Tang X L, Cao Y H, Gu L H, Zhou B Z. Advances in photo-physiological responses of leaves to environmental factors based on the FvCB model., 2017, 37: 6633–6645 (in Chinese with English abstract)
[3] Robredo A, Pérez-López U, Hector Sainz de la Maza, González-Moro B, Lacueata M, Mena-Petite A, Muñoz-Rueda A. Elevated CO2alleviates the impact of drought on barley improving water status by lowering stomatal conductance and delaying its effects on photosynthesis., 2007, 59: 252–263
[4] Wall G W, Garcia R L, Kimball B A, Hunaker D J, Pinter Jr. P J, Long S P. Interactive effects of elevated carbon dioxide and drought on wheat., 2006, 98: 354–381
[5] Kang S Z, Zhang F C, Hu X T, Zhang J H. Benefits of CO2enrichment on crop plants are modified by soil water., 2002, 238: 69–77
[6] Li F S, Kang S Z, Zhang J H. Interactive effects of elevated CO2, nitrogen and drought on leaf area, stomatal conductance, and evapotranspiration of wheat., 2004, 67: 221–233
[7] Wu D X, Wang G X, Bai Y F, Liao J X. Effects of elevated CO2concentration on growth, water use, yield and grain quality of wheat under two soil water levels.,, 2004, 104: 493–507
[8] Merewitz E B, Belanger F C, Warnke S E, Huang B R. Identification of quantitative trait loci linked to drought tolerance in a colonial´creeping bent grass hybrid population., 2012, 52: 1891–1901
[9] 郑云普, 徐明, 王建书, 邱帅, 王贺新. 玉米叶片气孔特征及气体交换过程对气候变暖的响应. 作物学报, 2015, 41: 601–612 Zheng Y P, Xu M, Wang J S, Qiu S, Wang H X. Responses of the stomatal traits and gas exchange of maize leaves to climate warming., 2015, 41: 601–612 (in Chinese with English abstract)
[10] Hugh J E. Stomatal and non-stomatal restrictions to carbon assimilation in soybean () lines differing in water use efficiency., 2002, 48: 237–246
[11] 许大全. 光合作用气孔限制分析中的一些问题. 植物生理学通讯, 1997, 33: 241–244 Xu D Q. Some problems in stomatal limitation analysis of photosynthesis., 1997, 33: 241–244 (in Chinese with English abstract)
[12] 孟凡超, 张佳华, 郝翠, 周正明, 李辉, 刘丹, 王凯, 张华. CO2浓度升高和不同灌溉量对东北玉米光合特性及产量的影响. 生态学报, 2015, 35: 2126–2135Meng F C, Zhang J H, Hao C, Zhou Z M, Li H, Liu D, Wang K, Zhang H. Effects of elevated CO2and different irrigation on photosynthetic parameters and yield of maize in northeast China., 2015, 35: 2126–2135 (in Chinese with English abstract)
[13] Van der Kooi C J, Reich M, Löw M, Tausz M. Growth and yield stimulation under elevated CO2and drought: a meta-analysis on crops., 2016, 122: 150–157
[14] 张凯, 王润元, 王鹤龄, 赵鸿, 齐月, 赵福年, 陈斐, 雷俊. CO2浓度升高对半干旱区春小麦生长发育及产量影响的试验研究. 干旱气象, 2017, 35: 306–312 Zhang K, Wang R Y, Wang H L, Zhao H, Qi Y, Zhao F N, Chen F, Lei J. Effects of elevated CO2concentration on growth and yield of spring wheat based on observational experiment in semi-arid area., 2017, 35: 306–312 (in Chinese with English abstract)
[15] 龚吉蕊, 黄永梅, 葛之葳, 段庆伟, 尤鑫, 安然, 张新时. 4种杂交杨对土壤水分变化的生态学响应. 植物生态学报, 2009, 33: 387–396 Gong J R, Huang Y M, Ge Z W, Duan Q W, You X, An R, Zhang X S. Ecological responses to soil water content in four hybridclones., 2009, 33: 387–396 (in Chinese with English abstract)
[16] 徐建文, 梅旭荣, 居辉, 李迎春, 刘勤, 杨建莹. 黄淮海地区冬小麦关键生育期不同灌溉水平对产量影响的模拟. 作物学报, 2014, 40: 1485–1492 Xu J W, Mei X R, Ju H, Li Y C, Liu Q, Yang J X. Simulation of winter wheat yield in response to irrigation level at critical growing stages in the Huang-Huai-Hai Plain., 2014, 40: 1485–1492 (in Chinese with English abstract)
[17] Ma S C, Duan A W, Wang R, Guan Z M, Yang S J, Ma S T, Shao Y. Root-sourced signal and photosynthetic traits dry matter accumulation and remobilization, and yield stability in winter wheat as affected by regulated deficit irrigation., 2015, 148: 123–129
[18] 姚宁, 宋利兵, 刘健, 冯浩, 吴淑芳, 何建强. 不同生长阶段水分胁迫对旱区冬小麦生长发育和产量的影响. 中国农业科学, 2015, 48: 2379–2389 Yao N, Song L B, Liu J, Feng H, Wu S F, He J Q. Effects of water stress at different growth stages on the development and yields of winter wheat in arid region., 2015, 48: 2379–2389 (in Chinese with English abstract)
[19] Ozturk A, Aydin F. Effect of water stress at various growth stages on some quality characteristics of winter wheat., 2004, 190: 93–99
[20] 胡梦芸, 张正斌, 徐萍, 董宝娣, 李魏强, 李景娟. 亏缺灌溉下小麦水分利用效率与光合产物积累运转的相关研究. 作物学报, 2007, 33: 1884–1891 Hu M Y, Zhang Z B, Xu P, Dong B D, Li W Q, Li J J. Relationship of water use efficiency with photo assimilate accumulation and transport in wheat under deficit irrigation., 2007, 33: 1884–1891 (in Chinese with English abstract)
Effects of Water and CO2Concentration on Stomatal Traits, Leaf Gas Exchange, and Biomass of Winter Wheat
WU Hai-Xia1,**, GUO Li-Li1,**, HAO Li-Hua1, ZHANG Hao2, WANG Qing-Tao3, CHENG Dong-Juan1, PENG Zheng-Ping4, LI Fei1, ZHANG Xi-Xi1, LI Shu-Bin5, XU Ming6,7,*, and ZHENG Yun-Pu1,*
1School of Water Conservancy and Hydropower, Hebei University of Engineering, Handan 056038, Hebei, China;2School of Life Science and Engineering, Handan University, Handan 056005, Hebei, China;3School of Landscape and Ecological Engineering, Hebei University of Engineering, Handan 056038, Hebei, China;4School of Resources and Environment Science, Hebei Agricultural University, Baoding 071001, Hebei, China;5Office of Fu-Zhang River Irrigation and Water Supply Management, Handan 056002, Hebei, China;6Key Laboratory of Ecosystem Network Observation and Modeling, Institute of Geographical Sciences and Natural Resources, Chinese Academy of Sciences, Beijing 100101, China;7School of Environment and Resources, University of Chinese Academy of Sciences, Beijing 100039, China
The experiment was conducted in environmental growth chambers with accurately controlled CO2concentrations. Water deficit resulted in the decrease of stomatal openness and more irregular stomatal distribution of winter wheat, and elevated CO2concentration ([CO2]) mitigated the negative impacts on the morphological traits of individual stoma and spatial distribution of stomata. The net photosynthetic rate, stomatal conductance, and transpiration rate of winter wheat significantly decreased (< 0.05) under water deficits than under full irrigation condition, while elevated [CO2] could mitigate the influence of mild water deficit on leaf gas exchange, and this mitigated effect declined with the increase of water deficit degree. In addition, the biomass of winter wheat was substantially decreased with water deficits but barely affected by elevated[CO2] under water deficits. These results suggested that winter wheat changes its leaf gas exchange by adjusting morphological traits of individual stoma and spatial distribution pattern of stomata under water deficit, and the “CO2fertilization effect” on winter wheat may be modified by soil water conditions.
doubling CO2concentration; water deficit; winter wheat; biomass; stomatal traits; gas exchange parameters
2018-02-01;
2018-07-20;
2018-08-03.
10.3724/SP.J.1006.2018.01570
郑云普, E-mail: zhengyunpu_000@sina.com; 徐明, E-mail: mingxu@igsnrr.ac.cn
**同等贡献(Contributed equally to this work)
武海霞, E-mail: whx2307@126.com
本研究由国家重点研发计划项目(2017YFD0300905)资助。
This study was supported by the National Key Research and Development Program of China (2017YFD0300905).
URL: http://kns.cnki.net/kcms/detail/11.1809.S.20180803.1726.002.html
