不同地理来源MGIII组大豆品种生育期结构分析及E基因型鉴定
2018-10-10宋雯雯吴存祥武婷婷胡水秀韩天富
江 红 孙 石 宋雯雯 吴存祥 武婷婷 胡水秀韩天富,*
不同地理来源MGIII组大豆品种生育期结构分析及基因型鉴定
江 红1,2,**孙 石2,**宋雯雯2,**吴存祥2武婷婷2胡水秀1,*韩天富2,*
1江西农业大学农学院/ 江西省教育部作物生理生态与遗传育种重点实验室, 江西南昌 330000;2中国农业科学院作物科学研究所 / 农业部北京大豆生物学重点实验室, 北京 100081
于2014¾2015年对60份不同地理来源、生育期组为MGIII的大豆品种进行生育期结构分析和基因型鉴定表明, 不同地理来源的MGIII大豆品种生育期相近, 但生育期结构差异较大。来自中国北方和美国的MGIII组春大豆品种营养生长期(V期)较短(开花较早), 生殖生长期(R期)较长, R期与V期的比值(R/V)较高; 黄淮海品种和南方MGIII组品种V期较长(开花较晚), R期较短, R期与V期的比值(R/V)较低。北方春大豆MGIII组品种的开花期受播期影响较其他地区同生育期组品种更为明显。中国MGIII组大豆品种存在6种基因型, 其中和分布区域广, 覆盖播季类型多, 而在8个美国MGIII组品种中只鉴定出1种基因型(), 表明中国大豆品种在生育期结构性状上存在更为丰富的遗传变异。通过比较不同播期下MGIII大豆品种基因在生育期性状上的平均效应值发现, 含显性位点越多的材料, 其V期越长, R期越短, R/V值越小。反之亦然。不同基因对开花、成熟期的增强效果不尽相同, 且春播时各显性基因的效应值均比夏播时大。不同地理来源MGIII组大豆品种农艺性状存在明显差异, 且与生育期结构存在显著相关性。中国北方春大豆品种底荚高度与R/V值负相关, 但单株荚数与R/V值正相关; 黄淮海大豆品种的分枝数、单株荚数、百粒重与R/V间无显著相关性; 南方大豆品种分枝数与V期呈显著负相关。试验结果可为大豆品种生育期结构的改良及适应不同环境的品种选育提供依据。
大豆; MGIII; 地理来源; 开花期;基因; 农艺性状
生育期性状包括全生育期、营养生长期(V期)、生殖生长期(R期)[1]和生育期结构(R/V)[2-3], 是大豆最重要的生态性状。我国大豆生产区域广大, 地理环境和耕作制度复杂, 各地大豆品种在生育期性状上形成了丰富的多样性[4-5]。宋雯雯等[6]在全国36个试验点对共计840个大豆品种进行生育期组精细划分, 证明我国大豆品种生育期组范围为MG000-IX。Jia等[7]在中国东北高纬度冷凉地区及俄罗斯远东大豆品种中发现早于MG000组的材料。宋雯雯等[6]进一步证明, MGIII是覆盖大豆品种数量最多的生育期组。部分学者发现, 同一生育期组品种在生育期结构上表现出明显差异[2-3,8]。大豆的生育期性状由包括~和在内的多个主效基因[9-25]及微效多基因控制[26]。除和外, 显性基因都对开花和成熟起到不同程度的延迟作用[13,16], 且在不同的生育期阶段存在加性效应[12], 其表达效应还受光周期、温度等环境因素[27-28]及播期的影响[19]。随着[20]、[21]、[22]、[23]、[16]、[24-25]等相继被克隆, 对不同大豆品种的基因型鉴定更加容易, 已发现丰富的位点等位变异[29-33]。其中的等位变异有等, 这些变异均会引起大豆光周期敏感性不同程度的弱化[20,30,34];的等位变异包括可造成功能缺失的和功能尚不清楚的及[30-31];的等位变异有、引起功能缺失的, 以及引起光周期敏感度变化的和s[22,32];的等位变异有及在高纬度大豆品种中发现的和[23,33]。近年研究发现,~等位变异组合对大豆品种的生育期组归属有决定性影响[35],基因的构成、基因之间及基因与环境因子(如光周期、温度)的互作也与生育期结构有关[3,36-37]。Jiang等[35]在北美生育期组标准品种中发现同一种~基因型对应于1个以上生育期组的现象; Jia等[7]在中国东北高纬度冷凉地区及俄罗斯远东地区MG0000-0组早熟大豆品种中发现1个生育期组对应多种~基因型的情况。然而, 对于中晚熟大豆品种基因型多态性尚未进行深入的研究。本试验收集到我国覆盖纬度范围最大、播季类型最多、最能体现我国生态类型和种植制度多样性的MGIII大豆代表性品种[5-6,38-39], 在北京地区种植, 对其生育期性状特点和基因型进行分析, 并比较不同来源MGIII组品种的农艺性状表现, 以期深入了解不同生态类型品种的生育期特点及与基因型和环境的关系, 为大豆生育期性状分子设计育种提供理论依据。
1 材料与方法
1.1 供试材料
60个供试材料分别来自美国和我国不同大豆生态区(表1), 这些品种经过多年多点生育期组鉴定试验, 已明确生育期组归属为MGIII[6]。
1.2 种植方法
田间试验于2014年和2015年进行, 春播均在中国农业科学院作物科学研究所院部试验农场进行, 播期分别为2014年4月30日和2015年5月4日; 夏播在该所昌平试验基地进行, 播期分别为2014年5月20日和2015年6月17日。春播时每品种1行, 行长2 m, 重复3次, 每行40粒, 定苗20株。夏播时每品种3行, 行长6 m, 重复3次, 每行120粒, 定苗60株。2年试验结果趋势一致, 但2014年大部分品种鼓粒至成熟期病虫害较重, 致使农艺性状数据不够完整, 故在结果分析中以2015年数据为主。
1.3 记载项目及标准
根据Fehr和Carviness[1]提出的大豆生育时期分期标准分株挂牌调查大豆的出苗期(VE)、初花期(R1)、生理成熟期(R7)和完熟期(R8)。昌平夏播收获时, 选取30份供试材料(9个北方品种, 15个黄淮海品种, 6个南方品种), 于每行随机取10株, 按照《大豆种质资源描述规范和数据标准》, 对株高、底荚高度、主茎节数、分枝数、总荚数、秕荚数及百粒重等产量相关性状进行考种。
1.4 DNA提取
用CTAB法提取大豆幼叶中的DNA, 1%的琼脂糖凝胶电泳检测DNA质量, 并用Naco 2000超微量分光光度计测定DNA的浓度后稀释成工作液25 ng μL–1。
1.5 E1、E2、E3和E4基因型的鉴定
根据Xu等[31]报道的方法, 用基因特异性引物扩增待鉴定品种的DNA, 再经琼脂糖凝胶电泳检测和测序分析, 对、、和进行基因型鉴定, 引物见表1。

表1 用于鉴定E1、E2、E3和E4基因型的引物
1.6 数据统计分析
利用IBM SPSS数据处理系统(IBM SPSS Statistics 19.0)对2015年供试品种的生育期性状及农艺性状分区域进行多重比较分析, 对生育期性状和农艺性状进行Pearson相关性分析。在分析基因显性位点的平均效应时, 选取背景基因型相同且仅1个基因在显、隐性位点上有差别的基因型计算二者表型差值, 再以该基因在不同背景基因型中表型差值的平均数作为该基因位点的效应值[27]。
2 结果与分析
2.1 不同地理来源MGIII组大豆品种生育期性状的比较
从表2可以看出, 不同地理来源的MGIII组大豆品种成熟期相近, 但V期、R期及R/V值存在明显差异且受播季影响。在春播条件下, 中国北方和北美春大豆MGIII组品种V期相近, 中国黄淮海和中国南方品种V期相近且均比中国北方及北美品种长, 而在R期上则情况相反; 中国北方和北美春大豆品种R/V值相近, 均值皆为3.1; 中国黄淮海和中国南方品种R/V值相近, 分别为2.1和1.9, 小于中国北方地区。在夏播条件下, 中国北方品种V期较短, R期较长, R/V值为2.3; 而中国黄淮海和中国南方品种的R/V值分别为1.6和1.4。夏播供试品种总体上比春播时V期延长, R期缩短, 全生育期缩短, R/V值变小, 属于“前延后促”[40]。
2.2 不同播季类型MGIII组大豆品种生育期性状的差异
从表3可以看出, 不同播季类型MGIII组的大豆品种成熟期相近, 但V期、R期及R/V值差异较大, 且受播期影响。在春播条件下, 北方春大豆(Sp) V期较短, R期较长, R/V平均值在各类品种中最大; 黄淮海夏大豆(Su)和南方春大豆(Sp**) V期相近且较北方春大豆(Sp)长, R期则相近且较北方春大豆(Sp)短, R/V值分别为2.1和1.9, 均比北方春大豆(Sp)小。在夏播条件下, 北方春大豆(Sp)V期最短, R期最长, R/V值最大; 黄淮海夏大豆(Su)和南方春大豆(Sp**)的V期相近且较北方春大豆(Sp)长, R期则较北方春大豆(Sp)短, R/V值分别为1.6和1.4, 均比北方春大豆(Sp)小。与春播相比, 夏播时存在“前延后促”现象[40], 在各播期类型品种中, 北方春播类型(Sp)“前延”幅度最大、“后促”幅度最小。
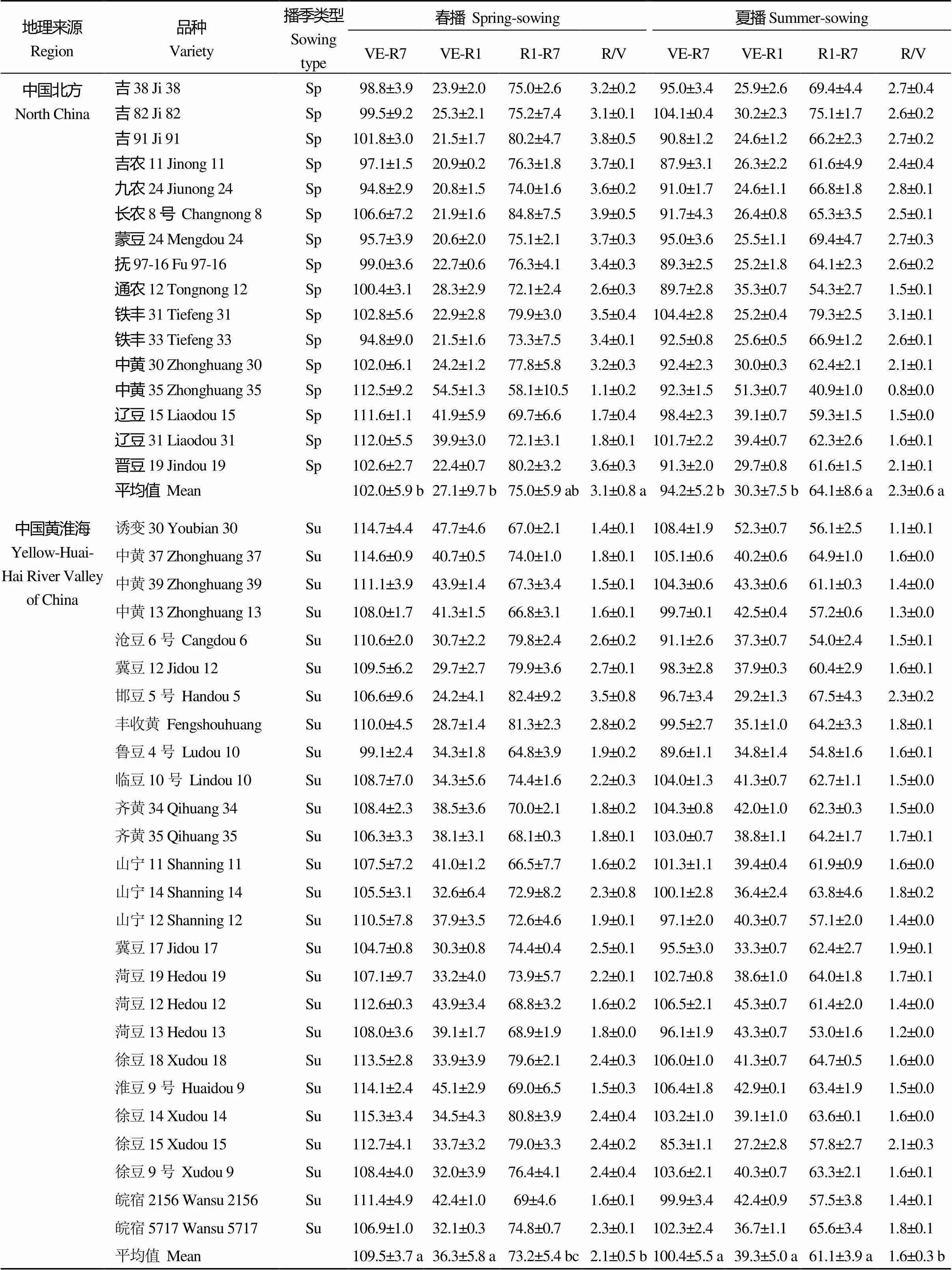
表2 不同地理来源MGIII大豆品种在北京春播和夏播条件下的生育期性状表现
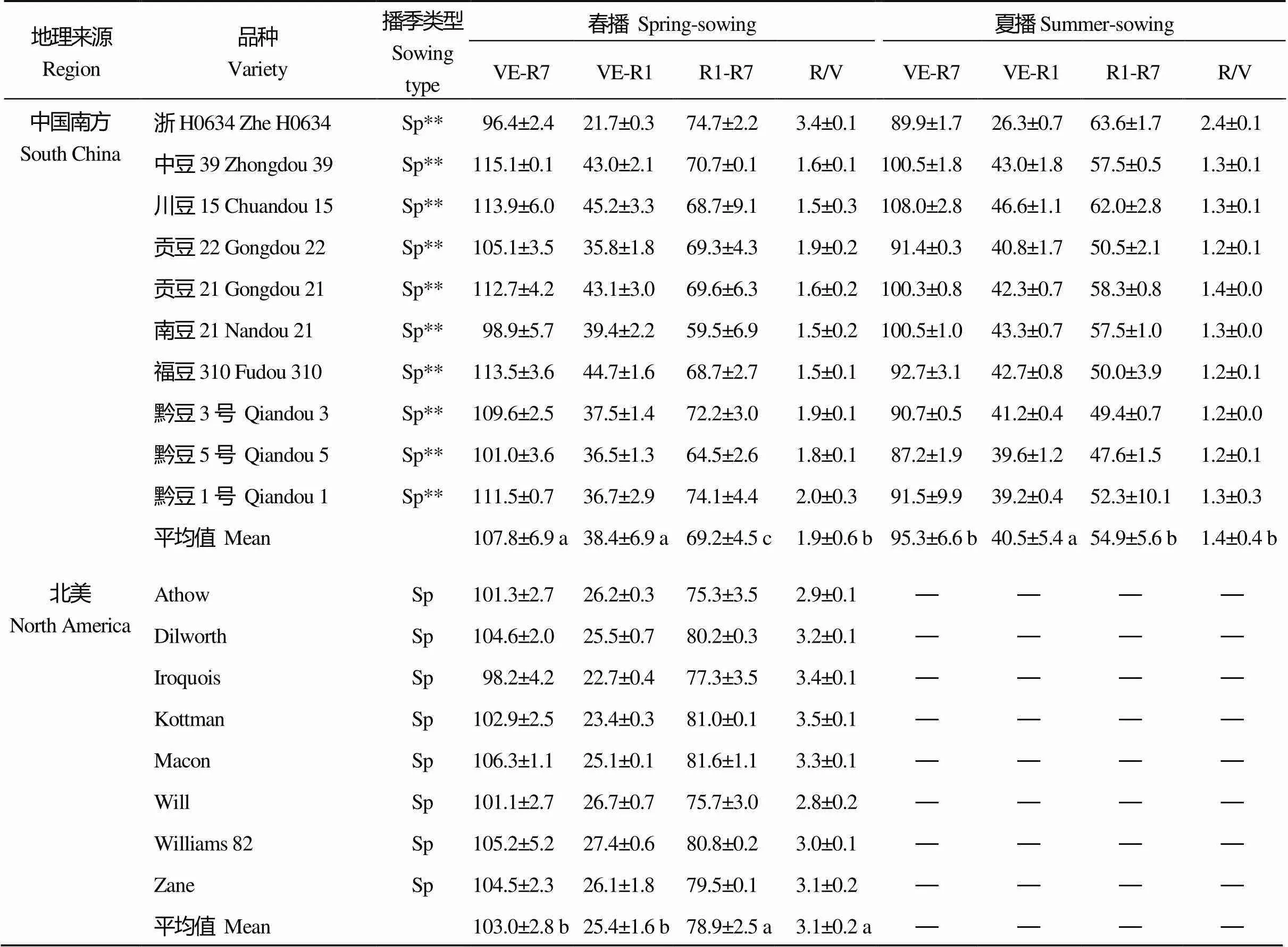
(续表2)
VE: 出苗期; R1: 初花期; R7: 生理成熟期; R/V: (R1-R7)/(VE-R1); Sp: 北方春播类型; Sp** 南方春播类型; Su: 黄淮海夏播类型。同一列内不同字母表示地区平均数间有显著差异(< 0.05)。
VE: emergence; R1: beginning bloom; R7: physiological maturity; R/V: (R1–R7)/(VE–R1); Sp: North spring-sowing soybean; Sp**: South spring-sowing soybean; Su: Yellow-Huai-Hai River Valley summer-sowing soybean. In the same column, the regional means followed by different lowercase letter are significantly different at< 0.05.
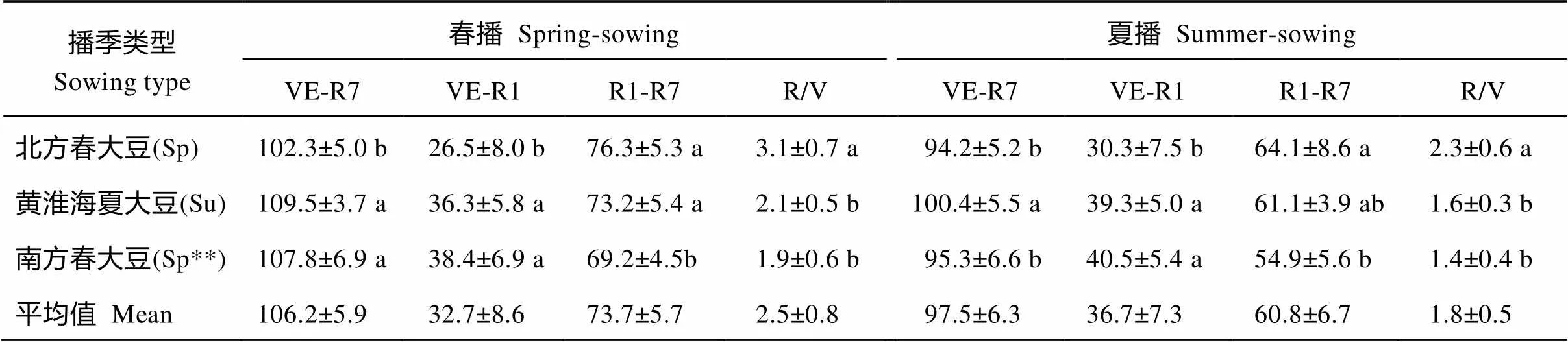
表3 不同播季类型MGIII大豆在北京春播和夏播条件下的生育期结构性状表现
缩写同表2。同一列内标以不同字母表示地区平均数间有显著差异(< 0.05)。
Abbreviations are the same as those given in Table 2. In the same column, the means followed by different lowercase letter are significantly different at< 0.05.
2.3 不同地理来源MGIII组大豆品种的E基因型构成
从表4可看出, 不同来源的中国MGIII组大豆品种的生育期主效基因(~)组成存在一定差异, 共有9种组合, 其中和编码同种氨基酸, 表现的基因功能一致, 故用表示这2种等位变异, 最终将基因型分为6种组合(9-3 = 6)。其中,和占的比例最大, 分别为78.8%和9.6%, 在我国分布区域广, 覆盖播季类型多;、、、所占的比例较低, 只分别在1~2个品种中被检测出。而在美国春大豆品种中只鉴定出, 与中国东北部分品种一致, 表现出该地区大豆品种生育期基因型的单一性。
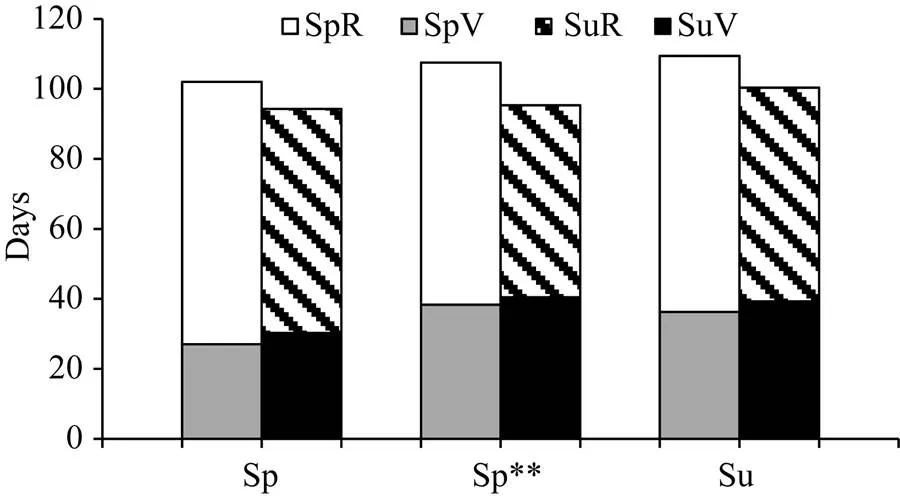
图1 春播和夏播时不同播季类型大豆品种在北京的生育期性状
SpV: 春播时营养生长期; SpR: 春播时生殖生长期; SuV: 夏播时营养生长期; SuR: 夏播时生殖生长期; Sp: 北方春播类型; Sp** 南方春播类型; Su: 黄淮海夏播类型。
SpV: vegetative growth period of spring-sowing soybean; SpR: reproductive growth period of spring-sowing soybean; SuV: vegetative growth period of summer-sowing soybean; SuR: reproductive growth period of summer-sowing soybean; Sp: North spring-sowing soybean; Sp**: South spring-sowing soybean; Su: Yellow-Huai-Hai River Valley summer-sowing soybean.
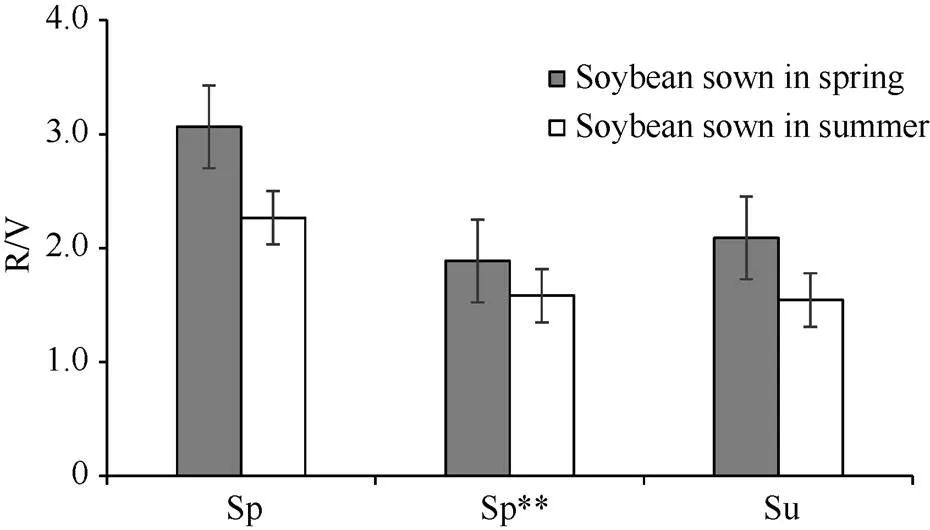
图2 春播和夏播时不同生态类型大豆品种的R/V值
Sp: 北方春播类型; Sp** 南方春播类型; Su: 黄淮海夏播类型。
Sp: North spring-sowing soybean; Sp**: South spring-sowing soybean; Su: Yellow-Huai-Hai River Valley summer-sowing soybean.
2.4 MGIII组大豆品种生育期结构与E基因型的关系
通过比较分析不同生育期主效基因型在春播和夏播条件下的全生育期、V期和R期发现, 大豆各生育期主效基因对大豆发育的不同阶段效应大小不一, 且存在明显的加性效应。总体而言, 含显性位点越多的材料, 其V期越长、R期越短、R/V值越小, 不同基因的增效作用大小不尽相同。含的大豆品种R/V值较大(3.3~3.5), 明显高于完全显性的大豆品种(R/V值在1.8~2.4之间)。此外, 基因与环境因子之间也存在明显的互作, 同一基因型大豆品种在夏播时的V期比在春播时长, R期则比春播时短, R/V值减小。不同基因型大豆品种受播季影响的程度也存在差异, 如在夏播条件下V期比春播延长了2 d, R期缩短了15 d, R/V值减小0.5, 而携带的大豆品种在夏播时V期比春播延长4 d, R期缩短11 d, R/V值减小1.0 (表5)。
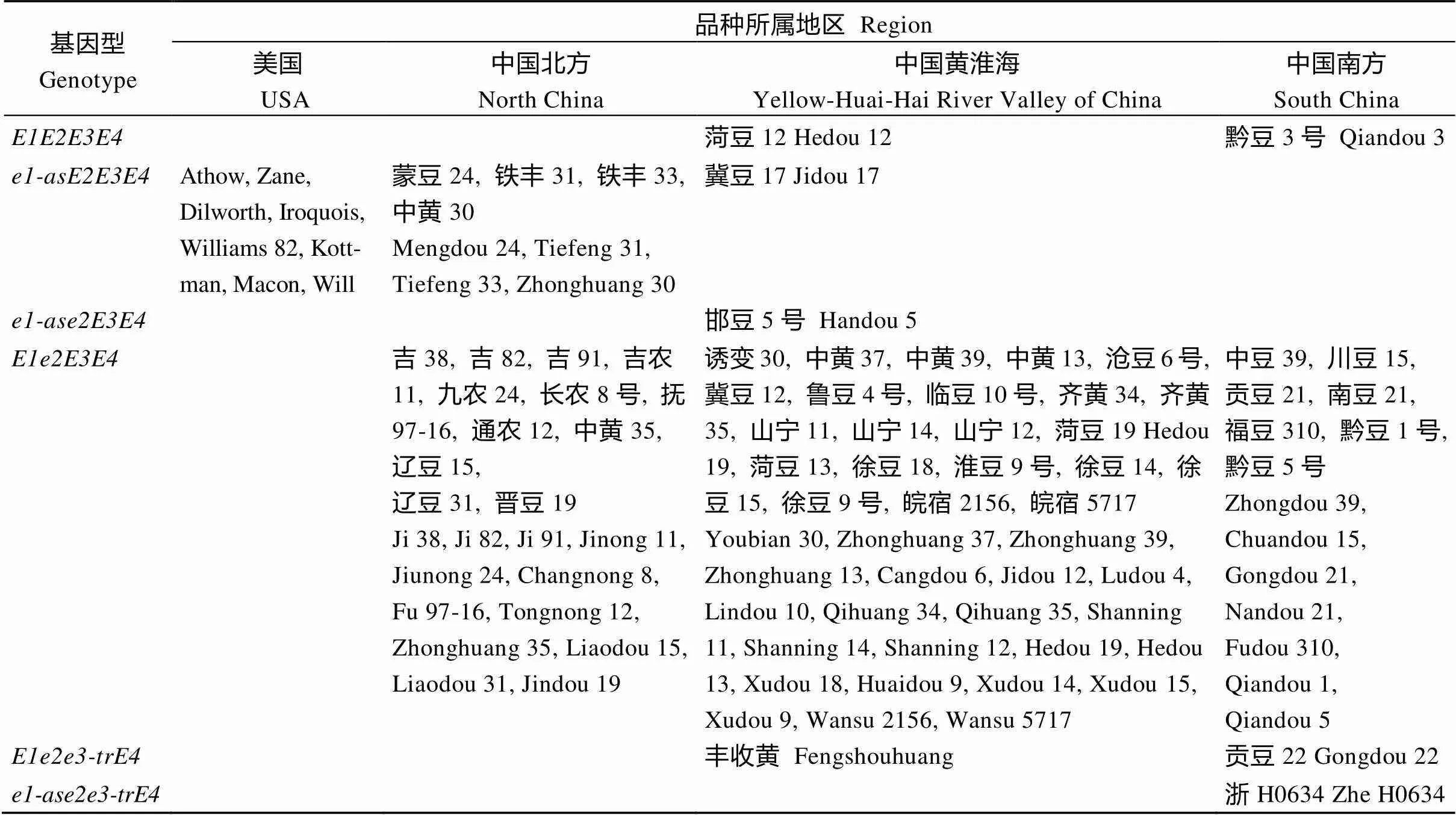
表4 MGIII大豆品种的E基因型分类和区域分布情况
同一大区内品种顺序按原产地纬度由北向南排列。
The varieties in the same area are arranged from north to south according to the latitude of origin
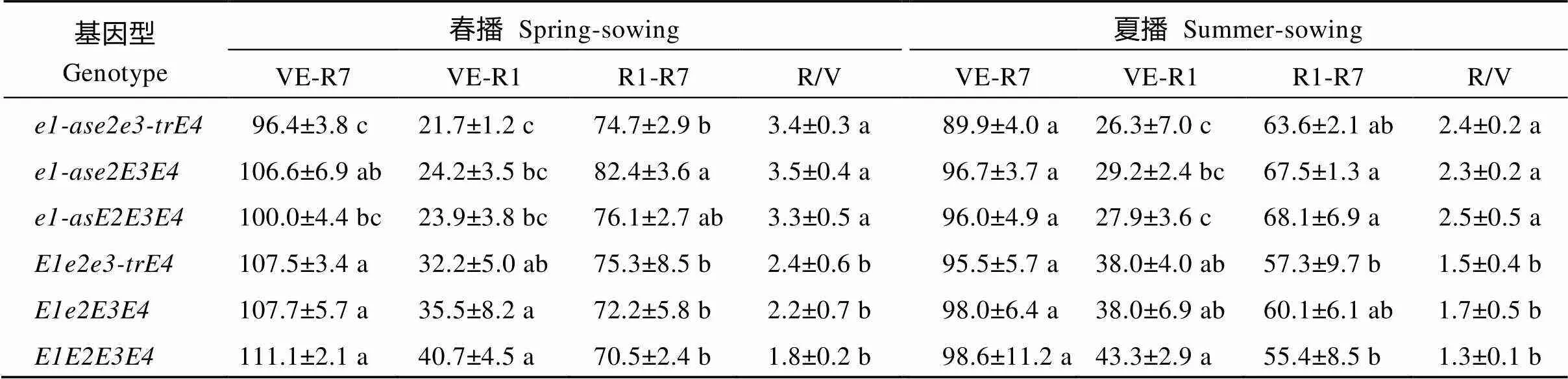
表5 不同E基因型的MGIII大豆品种在北京春播和夏播条件下的生育期性状表现
缩写同表2。同一列内标以不同字母表示地区平均数间有显著差异(< 0.05)。
Abbreviations are same as those given in Table 2. In the same column, the means followed by different lowercase letter are significantly different at< 0.05.
2.5 E基因等位变异对大豆生育期性状的影响及环境效应
从表6可以看出, 不同基因的表达效应在不同生育期阶段存在差异, 且受背景基因型影响, 如在不同背景基因型中的显性基因相对于隐性基因的平均效应值在V期是12.8, 在R期则是-5.0, 对R/V值的效应是-1.3; 当背景基因型为时,对V期的效应值是13.8, 对R期是-5.6, 对R/V值的效应是-1.5; 当背景基因型为时,对V期的效应值是10.5, 对R期是0.7, 对R/V值的效应值是-1.0。同时还可以看出,基因的表达效应与播期有关。在春播条件下,、能够延长V期、缩短R期、降低R/V值, 对生育期性状贡献值较大;能够延迟开花和成熟, 降低R/V值, 对生育期性状的贡献相对较小。在夏播条件下、、在延长V期和降低R/V值上的平均效应比春播时小,在缩短R期上的平均效应比春播时大,在夏播时的效应比春播时小,在延长R期上的平均效应比春播时大。

表6 不同播期下E基因对MGIII大豆品种生育期结构相关性状的效应
缩写同表2。Abbreviations are the same as those given in Table 2.
2.6 不同地理来源MGIII大豆品种生育期结构与农艺性状的关系
本研究选用的材料均属MGIII组, 生育期相近, 便于分析生育期结构与农艺性状的关系。从图3可以看出, 地理来源和生育期结构不同的MGIII组品种在农艺性状上存在明显差异。北方春大豆品种营养生长期较短, 生殖生长期较长, R/V值较大, 分枝数、总荚数较少, 百粒重偏小, 底荚较低; 南方品种营养生长期较长, 生殖生长期较短, 株高偏矮, 分枝多, 底荚较高, 百粒重较小; 黄淮海品种R/V值介于北方和黄淮海品种之间, 植株较高, 主茎节数、总荚数较多, 百粒重较大。
相关分析表明, MGIII组大豆品种的农艺性状与生育期结构之间存在一定的相关性。在夏播条件下, 北方春大豆品种的底荚高度与R/V值极呈显著负相关(<0.01), 单株荚数与R/V值呈极显著正相关(<0.01); 黄淮海大豆品种的底荚高度、分枝数、单株荚数、百粒重与R/V间无显著相关性; 南方大豆品种分枝数与V期呈显著负相关(<0.05)。从全国平均水平总体来看, 大豆品种底荚高度、分枝数与R/V值呈极显著负相关(<0.01)(表7)。

图3 夏播条件下不同来源MGIII大豆品种农艺性状的比较
PH: 株高; BPH: 底荚高度; NM: 主茎节数; BM: 分枝数; PP: 单株荚数; SW: 百粒重。
PH: plant height; BPH: bottom pods height; NM: node number on the main stem; BM: branch number; PP: pod number per plant;
SW: 100-seed weight.
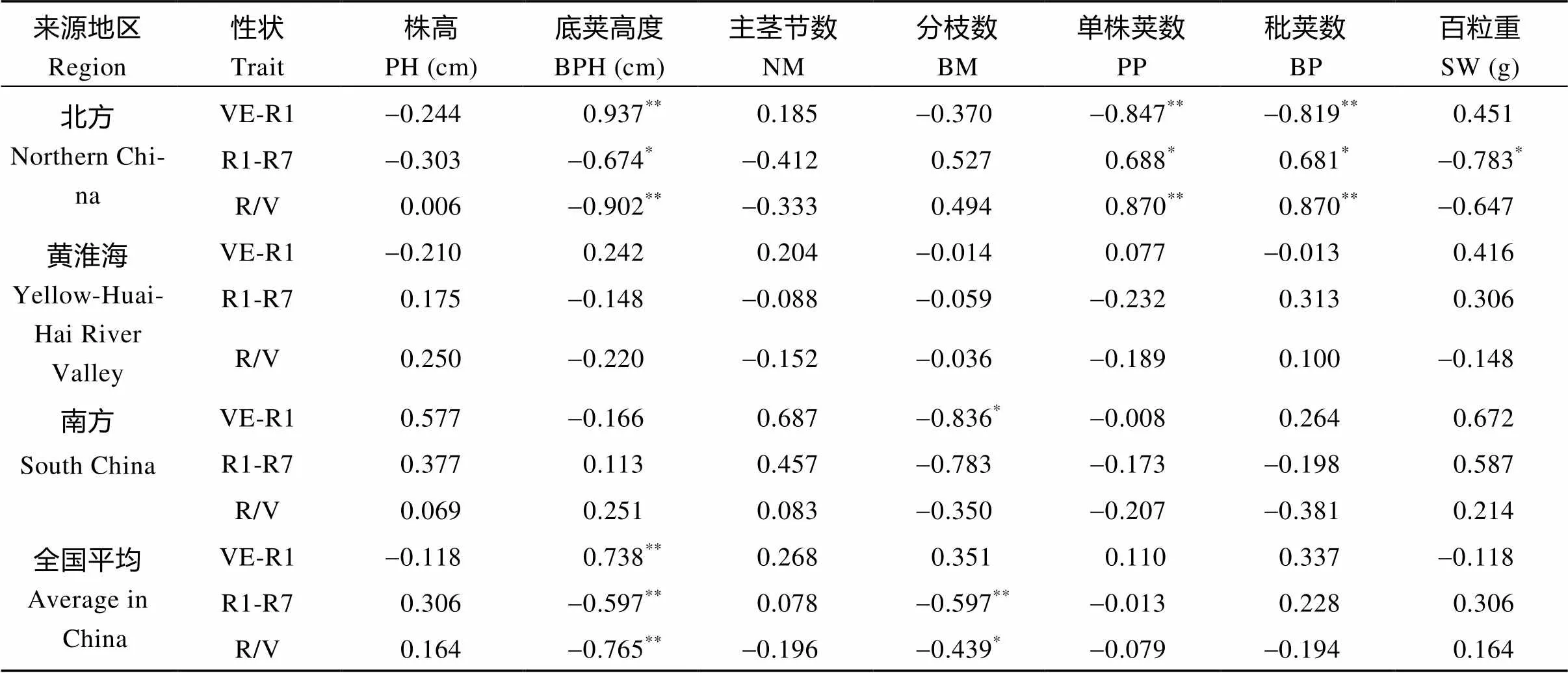
表7 不同地理来源MGIII组大豆品种生育期结构与其他农艺性状间的相关系数
缩写同图3。*和**分别表示在0.05和0.01水平上显著。
Abbreviations are the same as those given in Fig. 3.*and**represent significant correlations at the 0.05 and 0.01 probability levels, respectively.
3 讨论
3.1 不同来源MGIII组大豆品种生育期结构相关性状的多样性
MGIII组大豆品种分布广泛, 播季类型众多, 它们通过生育期结构的变化, 适应所在地区的自然环境和栽培制度。中国北方春大豆生长在长日、冷凉条件下, 相对较低的温度弱化了长日照对开花的抑制作用[41], 使这些品种在夏至后很快开花, 以保证大豆有足够长的结荚、鼓粒时间, 并在霜前正常成熟。这类品种开花较早, 生殖生长期较长, R/V较高。中国南方春大豆MGIII品种多分布在长江流域, 在春播条件下, 南方春大豆前期生长通常处于温度较低、日照较短的条件下, 低温弱化了短日对大豆发育的促进作用, 导致营养生长期相对较长, 有利于形成较大的营养体, 积累较多的干物质, 其较短的生殖生长期有利于大豆快速鼓粒、成熟, 缓解高温高湿等不利气象条件对种子发育的影响。这类品种开花较晚, R/V较小。美国大豆的祖先亲本主要从我国东北地区引进, 即北美MGIII组大豆品种的选育是以中国东北大豆品种为基础的[42-43], 其生育期性状与中国东北品种相似。以上皆表明, 不同地理来源的大豆品种, 尽管生育期组相同, 但在生育期结构上存在丰富的遗传变异。R/V值的合理调整, 可使大豆的生殖生长阶段处于较好的气象条件下, 规避干旱等逆境的影响。通过选用生育期结构合适的品种或调整播期等措施实现“花雨相遇”是大豆充分利用自然降水的重要措施。在相近生育期组品种分布地区进行穿梭育种则可望选育出广适应品种。中国MGIII组品种分布在北方、黄淮海和南方地区, 可在该组品种中率先开展跨区穿梭育种。
3.2 MGIII组大豆品种生育期基因型的复杂性
本研究发现, 中国MGIII大豆品种存在6种基因型, 而在北美标准品种中只鉴定出一种基因型, 且与中国北方春大豆部分品种一致。吴存祥[44]利用与大豆成熟期有关的SSR标记, 分析中国国家区域试验对照品种与北美大豆熟期组标准品种的基因型, 发现中国大豆品种在生育期性状上存在更为丰富的遗传变异。Jiang等[35]研究报道, 北美标准品种生育期组与基因型拟合程度较好, 能较真实地反映基因型的特征, 而中国品种基因型与表型的拟合程度相对较差。正如我们所知, 北美地区大豆生育期组分布区域和纬度是基本平行的[5], 而我国MGIII大豆品种覆盖纬度范围大, 表明生育期性状的遗传变异受地理纬度影响。Liu等[39]研究发现, MGIII大豆品种根据“开花期占全生育期的比重”可被划分出2个亚组, 亚组的划分与品种的复种制度有关, 春播类型前期短后期长, 夏秋播类型前期长后期短。我国各地种植制度多样, 大豆生育期基因型丰富。而北美大豆生产大多采用一年一熟制, 大豆品种类型单一, 生育期结构及基因型相对简单。
3.3 生育期主效基因在大豆生育期结构调整和适应性改良中的应用
Li等[38]研究证明基因组合是决定大豆生育期组归属的主要遗传因素。本研究进一步证明, 不同基因对大豆的开花期和成熟期均有影响, 且基因间存在加性效应, 这与前人研究结果一致[12,27]。可延长大豆的营养生长期, 缩短生殖生长期, 降低R/V值。的突变型可缩短营养生长期, 延长生殖生长期, 提高R/V值, 可作为大豆生育期结构调整的重要基因资源。可延迟开花和成熟期, 降低R/V值, 对大豆生育期性状的贡献值仅次于。延迟开花和成熟期, 但对生育期结构的影响较小。延迟开花和成熟期, 在开花后受光周期影响较大。Saindon等[45]研究发现的效应大于, 且对有上位作用。此外,~同一基因型的大豆品种在生育期性状上表现存在一定差异, 其中,类型的MGIII组品种数目最多, 品种间开花期变幅较大, 表明除、、和外, 还有其他重要基因参与对大豆生育期性状和光周期反应的调控[27]。在不断挖掘大豆生育期相关基因的基础上, 可针对目标地区的环境条件和耕作制度开展品种生育期结构设计育种, 并根据品种基因型预测其生育期组归属和在特定条件下的开花期、成熟期。当前, 在以北美标准品种为对照、根据生育期相对长度对大豆品种进行生育期组划分时, 可以结合生育期基因型, 进一步校验大豆品种熟期组归属, 为大豆品种跨区适应性评价和相互引种提供依据。
对MGIII大豆品种生育期性状与产量性状的相关性分析发现, R/V值与农艺性状的关系在不同地区和播期类型品种中有一定区别, 且以底荚高度、分枝数与R/V值负相关程度最大, 而合适的底荚高度和分枝数对于提高品种的抗逆性和满足人们利用需求有重要意义, 以上研究结果可以为大豆生育期相关基因的有效利用和生育期结构的定向和定量改良提供依据, 为进一步研究R/V与大豆抗逆性的关系以及选育广适应抗逆品种提供帮助。
4 结论
MGIII组大豆品种在生育期结构和生育期基因型方面存在丰富的遗传多样性, 这些变异是复杂多样的地理环境、耕作制度及产量选择共同作用的结果。中国北方和北美春大豆品种开花较早, R/V较高; 黄淮海品种和南方品种开花期相近且较晚, R/V较低。在中国大豆品种中鉴定出6种基因型, 而供试的美国大豆品种只有一种基因型。不同基因在不同背景基因型、生育期阶段和播期下的效应值不尽相同。含显性位点越多的材料, 其营养生长期越长、生殖生长期越短、R/V值越小, 含位点的大豆品种R/V值比较大。R/V值与产量性状存在显著的相关性, 从总体上看, 大豆品种的R/V值与底荚高度、分枝数呈显著负相关。
[1] Fehr W R, Caviness C E. Stages of Soybean Development. In: Special Report 80. Ames: Cooperative Extension Service, Agriculture and Home Economic Experiment Station, Iowa State University, 1977. pp 1–11
[2] 孙志强, 田佩占, 王继安. 东北大豆品种生育期结构的初步分析. 大豆科学, 1990, 9: 198–205 Sun Z Q, Tian P Z, Wang J A. Preliminary study on the growth period structure of soybean varieties in the northeast of China., 1990, 9: 198–205 (in Chinese with English abstract)
[3] 韩天富, 盖钧镒, 陈风云, 邱家驯. 生育期结构不同的大豆品种的光周期反应和农艺性状. 作物学报, 1998, 24: 550–557 Han T F, Gai J Y, Chen F Y, Qiu J X. Photoperiod response and agronomic characters of soybean varieties with different growth period structures., 1998, 24: 550–557 (in Chinese with English abstract)
[4] 王金陵, 祝其昌. 大豆生育期遗传的初步研究. 作物学报, 1963, 2: 333–335 Wang J L, Zhu Q C. The preliminary research on the growth period of soybean genetic., 1963, 2: 333–335 (in Chinese)
[5] 吴存祥, 李继存, 沙爱华, 曾海燕, 孙石, 杨光明, 周新安, 常汝镇, 年海, 韩天富. 国家大豆品种区域试验对照品种的生育期组归属. 作物学报, 2012, 38: 1977–1987 Wu C X, Li J C, Sha A H, Zeng H Y, Sun S, Yang G M, Zhou X A, Chang R Z, Nian H, Han T F. Maturity group classification of check varieties in national soybean uniform trials of China.2012, 38: 1977–1987 (in Chinese with English abstract)
[6] 宋雯雯. 中国大豆品种生育期组的精细划分与应用.中国科学院大学博士学位论文, 北京, 2016 Song W W. Digitized Classification and Application of Soybean Variety Maturity Groups in China. PhD Dissertation of University of Chinese Academy of Sciences, Beijing, China, 2016 (in Chinese with English abstract)
[7] Jia H C, Jiang B J, Wu C X, Lu W C, Hou W S, Sun S, Yan H R, Han T F. Maturity group classification and maturity locus genotyping of early-maturing soybean varieties from high-latitude cold regions., 2014, 9: e94139
[8] Wang Y, Cheng L R, Leng J T, Wu C X, Shao G H, Hou W S, Han T F. Genetic analysis and Quantitative trait locus identification of the reproductive to vegetative growth period ratio in soybean ((L.) Merr.)., 2015, 201: 275–284
[9] Bernard R L. Two major genes for time of flowering and maturity in soybeans., 1971, 11: 242–244
[10] Buzzell R I. Inheritance of a soybean flowering response to fluorescent daylength conditions., 1971, 13: 703–707
[11] Buzzell R I, Voldeng H D. Inheritance of insensitivity to long day length., 1980, 7: 26–29
[12] McBlain B A, Bernard R L, Cremeens C R, Korczak J F. A procedure to identify genes affecting maturity using soybean isoline testers., 1987, 27: 1127–1132
[13] Bonato E R, Vello N A., a dominant gene conditioning early flowering and maturity in soybeans., 1999, 22: 229–232
[14] Cober E R, Voldeng H D. A new soybean maturity and photoperiod sensitivity locus linked toand., 2001, 41: 698–701
[15] Cober E R, Molnar S J, Charette M, Voldeng H D. A new locus for early maturity in soybean., 2010, 50: 524–527
[16] Kong F J, Nan H Y, Cao D, Li Y, Wu F F, Wang J L, Lu S J, Yuan X H, Cober E R, Abe J, Liu B H. A new dominant geneconditions early flowering and maturity in soybean., 2014, 54: 2529–2535
[17] Samanfar B, Molnar S J, Charette M, Schoenrock A, Dehne F, Golshani A, Belzile F, Cober E R. Mapping and identification of a potential candidate gene for a novel maturity locus,, in soybean., 2017, 130: 377–390
[18] Ray J D, Hinson K, Mankono J E B. Genetic control of a long-juvenile trait in soybean., 1995, 35: 1001–1006
[19] 常汝镇, 李星华. 夏播条件下大豆成熟期基因作用的研究. 中国油料, 1993, (3): 15–17 Chang R Z, Li X H. Study on effect of maturity genes in soybeans under summer sowing condition., 1993, (3): 15–17 (in Chinese with English abstract)
[20] Xia Z J, Watanabe S, Yamada T, Tsubokura Y, Nakashima H, Zhai H, Anai T, Sato S, Yamazaki T, Lyu S X, Wu H Y, Tabata S, Harada K. Positional cloning and characterization reveal the molecular basis for soybean maturity locusthat regulates photoperiodic flowering.2012, 109: 2155–2164
[21] Watanabe S, Xia Z J, Hideshima R, Tsubokura Y, Sato S, Yamanaka N, Takahashi R, Anai T, Tabata S, Kitamura K. A map-based cloning strategy employing a residual heterozygous line reveals that thegene is involved in soybean maturity and flowering., 2011, 188: 395–407
[22] Watanabe S, Hideshima R, Xia Z J, Tsubokura Y, Sato S, Nakamoto Y, Yamanaka N, Takahashi R, Ishimoto M, Anai T. Map-based cloning of the gene associated with the soybean maturity locus., 2009, 182: 1251–1262
[23] Tsubokura Y, Matsumura H, Xu M L, Liu B H, Nakashima H, Anai T, Kong F J, Yuan X H, Kanamori H, Katayose Y. Genetic variation in soybean at the maturity locusis involved in adaptation to long days at high latitudes., 2013, 3: 117–134
[24] Yue Y L, Liu N X, Jiang B J, Li M, Wang H, Jiang Z, Pan H T, Xia Q J, Ma Q B, Han T F, Nian H. A single nucleotide deletion inencodingconfers long juvenility and is associated with adaption of tropic soybean., 2017, 10: 656–658
[25] Lu S J, Zhao X H, Hu Y L, Liu S L, Nan H Y, Li X M, Fang C, Cao D, Shi X Y, Kong L P, Su T, Zhang F G, Li S C, Wang Z, Yuan X H, Cober E R, Weller J L, Liu B H, Hou X L, Tian Z X, Kong F J. Natural variation at the soybeanlocus improves adaptation to the tropics and enhances yield.,2017, 49: 773–779
[26] Cheng L R, Wang Y, Zhang C B, Wu C X, Xu J L, Zhu H Y, Leng J T, Bai Y N, Guan R X, Hou W S, Zhang L J, Han T F. Genetic analysis and QTL detection of reproductive period and post-flowering photoperiod responses in soybean., 2011, 123 : 421–429
[27] 王英, 吴存祥, 张学明, 王云鹏, 韩天富. 不同光周期条件下大豆生育期主基因的效应. 作物学报, 2008, 34: 1160–1168 Wang Y, Wu C X, Zhang X M, Wang Y P, Han T F. Effects of soybean major maturity genes under different photoperiods., 2008, 34: 1160–1168 (in Chinese with English abstract)
[28] Mao T T, Li J Y, Wen Z X, Wu T T, Wu C X, Sun S, Jiang B J, Hou W S, Li W B, Song Q J, Wang D C, Han T F. Association mapping of loci controlling genetic and environmental interaction of soybean flowering time under various photo-thermal conditions., 2017, 18: 415–423
[29] 夏正俊. 大豆光周期反应与生育期基因研究进展. 作物学报. 2013, 39: 571–579 Xia Z J. Research progresses on photoperiodic flowering and maturity genes in soybean (Merr.)., 2013, 39: 571–579 (in Chinese with English abstract)
[30] Tsubokura Y, Watanabe S, Xia Z J, Kanamori H, Yamagata H, Kaga A, Katayose Y, Abe J, Ishimoto M, Harada K. Natural variation in the genes responsible for maturity loci,,andin soybean,2014, 113: 429–441
[31] Xu M L, Xu Z H, Liu B H, Kong F J, Tsubokura Y, Watanabe S, Xia Z J, Harada K, Kanazawa A, Yamada T. Genetic variation in four maturity genes affects photoperiod insensitivity and PHYA-regulated post-flowering responses of soybean., 2013, 13: 91
[32] Harada K, Watanabe S, Xia Z J, Tsubokura Y, Yamanaka N, Anai T. Positional cloning of the responsible genes for maturity Loci,andin soybean. In: Krezhovaed D eds. Soybean-Genetics and Novel Techniques for Yield Enhancement. Shanghai: InTech, 2011. pp 51–76
[33] Liu B, Kanazawa A, Matsumura H, Takahashi R, Harada K, Abe J. Genetic redundancy in soybean photoresponses associated with duplication ofgene., 2008, 180: 996–1007
[34] Hong Z, Lu S X, Wang Y Q, Chen X, Ren H X, Yang J Y, Cheng W, Zong C M, Gu H P, Qiu H M, Wu H Y, Zhang X Z, Cui T T, Xia Z J. Allelic variations at four major maturitygenes and transcriptional abundance of thegene are associated with flowering time and maturity of soybean cultivars., 2014, 9: e97636
[35] Jiang B J, Nan H Y, Gao Y F, Tang L L, Yue Y L, Lu S J, Ma L M, Cao D, Sun S, Wang J L, Wu C X, Yuan X H, Hou W S, Kong F J, Han T F, Liu B H. Allelic combinations of soybean maturity loci,,andresult in diversity of maturity and adaptation to different latitudes., 2014, 9: e106042
[36] Mansur L M, Orf J H, Chase K, Jarvik T, Cregan P B, Lark K G. Genetic mapping of agronomic traits using recombinant inbred lines of soybean., 1996, 36: 1327–1336
[37] Tian Z X, Wang X B, Lee R, Li Y H, Specht J E, Nelson R L, McClean P E, Qiu L J, Ma J X. Artificial selection for determinate growth habit in soybean.2010, 107: 8563–8568
[38] Li J C, Wang X B, Song W W, Huang X Y, Zhou J, Zeng H Y, Sun S, Jia H C, Li W B, Zhou X A, Li S Z, Chen P Y, Wu C X, Yong G, Han T F, Qiu L J. Genetic variation of maturity groups and fourgenes in the Chinese soybean mini core collection., 2017, 12: e0172106
[39] Liu X Q, Wu J A, Ren H X, Qi Y X, Li C Y, Cao J Q, Zhang X Y, Zhang Z P, Cai Z Y, Gai J Y. Genetic variation of world soybean maturity date and geographic distribution of maturity groups., 2017, 67: 221–232
[40] 刘汉中. 大豆引种中发育“延”、“促”效应之再论证. 中国农业大学学报, 1988, 7(1): 89–96 Liu H Z. A further discussion on “enhancing” and “delaying” effects on soybean development by daylength and temperature during variety introduction., 1988, 7(1): 89–96 (in Chinese with English abstract)
[41] 费志宏, 吴存祥, 孙洪波, 侯文胜, 张宝石, 韩天富. 以光周期处理与分期播种试验综合鉴定大豆品种的光温反应. 作物学报, 2009, 35: 1525–1531 Fei Z H, Wu C X, Sun H B, Hou W S, Zhang B S, Han T F. Identification of photothermal responses in soybean by integrating photoperiod treatments with planting-date experiments., 2009, 35: 1525–1531 (in Chinese with English abstract)
[42] 邱丽娟, 常汝镇, 袁翠平, 关荣霞, 刘章雄, 李英慧. 国外大豆种质资源的基因挖掘利用现状与展望. 植物遗传资源学报, 2006, 7(1): 1–6 Qiu L J, Chang R Z, Yuan C P, Guan R X, Liu Z X, Li Y H. Prospect and present statue of gene discovery and utilization for introduced soybean germplasm.2006, 7(1): 1–6 (in Chinese with English abstract)
[43] 汪越胜, 马宏惠. 美国的大豆熟期划分及其影响. 安徽农学通报, 2000, 6(4): 28–29 Wang Y S, Ma H H. The division and influence of soybean maturity in the United States.20006(4): 28–29 (in Chinese)
[44] 吴存祥. 中国不同生态类型大豆品种的生态适应性研究. 华南农业大学博士学位论文, 广东广州, 2009 Wu C X. Studies on the Ecological Adaptability of Different Soybean Ecotypes in China. PhD Dissertation of South China Agricultural University, Guangzhou, Guangdong, China, 2009 (in Chinese with English abstract)
[45] Saindon G, Voldeng H D, Beversdorf W D, Buzzell R I. Genetic control of long daylength response in soybean., 1989, 29: 1436–1439
Characterization of Growth Period Structure and Identification ofGenes of MGIII Soybean Varieties from Different Geographic Regions
JIANG Hong1,2,**, SUN Shi2,**, SONG Wen-Wen2,**, WU Cun-Xiang2, WU Ting-Ting2, HU Shui-Xiu1,*, and HAN Tian-Fu2,*
1College of Agriculture, Jiangxi Agricultural University, Nanchang 330000, Jiangxi, China;2MOA Key Laboratory of Soybean Biology (Beijing) / Institute of Crop Sciences, Chinese Academy of Agricultural Sciences, Beijing 100081, China
In 2014 and 2015, 60 Maturity Group (MG) III soybean varieties from different geographic regions were chosen to characterize the maturity-related traits and to identify the alleles of the maturity genes (alleles) in these varieties. The MGIII soybean varieties shared the similar growth period but owned different growth period structure. The spring-sowing MGIII soybean varieties from Northern China and the US bloomed earlier (shorter VP) and exhibited lower R/V values than those from Yellow-Huai-Hai River Valley (YHH)and South China. Six combinations ofalleles were identified in the tested Chinese varieties, among themandwere found to distribute in larger areas of different regions and cover more ecotypes than other four combinations. In contrast, the type ofwas the onlyallele combination in the eight US standard varietiesindicating that the Chinese MGIII varieties were more abundant in genetic variations for the maturity-related traits. By comparing the average effect of eachallele on the maturity-related traits in MGIII soybean varieties under different sowing dates, it was found that the varieties with more dominantalleles showed longer VP, shorter RP and smaller R/V value. The effects of differentalleles on flowering time and maturity were different, and the effect of the dominantalleles on spring- sowing soybeans was stronger than that on the summer-sowing ones. The agronomic traits for MGIII varieties from different regions were significantly different and that correlated with the growth period structure. The height of bottom pods of MGIII soybean varieties from Northern China was negatively correlated with R/V, while the number of pods per plant was positively correlated with R/V. Branch number, pod number per plant and 100-seed weight of YHH MGIII varieties were not correlated with R/V. The branch number of southern soybean varieties was significant negative correlated with the VP. These results provide a basis for the improvement of the growth period structure of soybean varieties and adaptability of soybeans to multiple environments.
soybean; MGIII; geographical origin;gene; agronomic traits
2018-02-14;
2018-06-12;
2018-07-02.
10.3724/SP.J.1006.2018.01448
韩天富, E-mail: hantianfu@caas.cn; 胡水秀, E-mail: 45461707@qq.com
**同等贡献(Contributed equally to this work)
江红, E-mail: jianghongnuli@163.com; 孙石, E-mail: sunshi@caas.cn
本研究由国家重点研发计划项目(2017YFD0101400), 国家现代农业产业技术体系建设专项(CARS-04)和中国农业科学院农业科技创新工程资助。
This study was supported by the National Key R&D Program of China (2017YFD0101400), the China Agriculture Research System (CARS-04), and the Agricultural Science and Technology Innovation Program of the Chinese Academy of Agricultural Sciences.
URL: http://kns.cnki.net/kcms/detail/11.1809.S.20180630.1337.004.html
