PPARγ基因干扰体外对兔骨髓间充质干细胞成脂基因的影响
2018-10-08禹宝庆黄建明许大峰
白 祥 禹宝庆 黄建明 许大峰
(上海市浦东医院骨科,上海 201300)
骨髓基质系统是由基质细胞及细胞外基质组成的网络组织。骨髓基质细胞中存在具有高度增殖及多向分化能力的多潜能骨髓间充质干细胞(BMSCs)。酒精作用于细胞时,BMSCs分化为成骨细胞明显减少,分化为脂肪细胞大为增加。脂肪细胞数量随着酒精的作用时间延长及浓度递增而增多,细胞内三酰甘油含量明显增高,说明酒精能够诱导BMSCs分化为脂肪细胞〔1〕。而且其诱导脂肪细胞分化具有一定的剂量依赖性。酒精促进了BMSCs成脂基因表达,在某个环节诱导BMSCs中多潜能基质干细胞分化为脂肪细胞〔2〕。当血液循环中的脂肪物质增多,聚集成脂肪球,并可通过多种途径最终导致骨股头坏死(ANFH)。本研究通过正常兔股骨骨髓培养细胞证实过氧化物酶体增殖物激活受体(PPAR)γ抑制剂T0070907是否能抑制酒精诱导骨髓基质细胞分化为脂肪细胞,从而阐明其分子机制。
1 材料与方法
1.1材料 实验动物:新西兰大白兔,雌雄各半,SPF级,4~6周龄(上海交通大学动物实验基地)。siRNA腺病毒:对兔PPARγ基因的siRNA腺病毒(pAdeno-U6-siPPARγ),空载siRNA腺病毒(郑州大学第一附属医院惠赠)。试剂:高糖DMEM培养基、胎牛血清(美国Gibco公司),RNA提取试剂盒(Qiagen公司),三酰甘油试剂盒(温州津玛生物科技有限公司);Trizol试剂盒购自Invitrigen公司。鼠抗兔CD90-FITC,CD73-FITC,CD105-FITC,CD13-FITC,CD45-FITC,CD34-FITC(Santa Cruz)。仪器:流式细胞仪(BD,FACSCalibur),离心机(1-15K,Sigma),分光光度计(DU730,Beckman),CO2培养箱(Thermo Forma 370)、显微镜(奥林巴斯CX23)、超净工作台(苏信YJ-840/YJ-1340)、移液器(Eppendorff)等。
1.2BMSCs细胞的分离、培养和鉴定 将6周龄新西兰大白兔处死后,清洗干净,在无菌操作下取大白兔四肢长骨,用咬骨钳打开长骨髓腔,用无血清高糖DMEM培养基冲洗髓腔,从而获得骨髓,3 000 r/min离心3 min,去除悬液,加入DMEM培养液4 ml后反复吹打,使细胞成为单个细胞悬液,并以6×106/cm2接种在培养瓶中,加入完全DMEM培养液,温度37℃,5%CO2孵育箱中培养,培养4 d后,去除悬浮细胞,此后隔日换液一次,获取贴壁生长的细胞即为BMSCs。用倒置显微镜观察细胞的形态和生长情况,将原代细胞传代后,取二代BMSCs细胞分别接种于6孔培养板中,以每孔1×104接种,设为实验0 d。取约1×106个第5代细胞悬于100 μl磷酸盐缓冲液(PBS)中,分别加入各抗体室温反应30 min,PBS洗涤去除未结合抗体后重悬于PBS中,上流式细胞仪检测。
1.3实验分组 将实验分为5组,每组3个培养孔。对照组:BMSCs未经任何处理;模型组:BMSCs给予0.09 mol/L乙醇处理;空siRNA组:MSCs给予0.09 mol/L乙醇+空siRNA;PPARγ组:BMSCs给予0.09 mol/L乙醇+PPARγ siRNA;T0070907组:BMSCs给予0.09 mol/L乙醇+ T0070907。
1.4RT-PCR检测 BMSCs在乙醇诱导培养7 d后,运用RT-PCR方法检测PPARγ、PETALA2(AP2)、脂蛋白脂肪酶(LPL)、脂肪酸转运蛋白(FATP)1 和脂肪酸转运酶(FAT)基因表达水平及ANFH模型治疗6 w后,股骨头组织PPARγ表达水平。采用总RNA提取试剂Trizol提取标本组织中的总RNA;取1 μl总RNA、AMV逆转录酶0.5 μl进行反转录;取模板cDNA 2.5 μl,聚合酶Ex Taq HS 0.1 μl,引物合成见表1,上游引物0.1 μl,下游引物0.1 μl。反应条件为:94℃预变性2 min,94℃变性40 s,50~65℃退火40 s,72℃延伸1 min,共进行35个循环,72℃再延伸5 min,1个循环。所得PCR产物-20℃冰箱保存。每组均以β-actin作为内参照,同样条件下分管进行扩增;配制2%琼脂糖凝胶;取PCR产物6 μl,2%琼脂糖凝胶电泳120 V,100 mA,30 min。电泳结束后,取出凝胶置于EB液中约5 min,取出凝胶。RT-PCR结果判定:电泳条带清晰并符合预期扩增片段大小,判定为PCR阳性条带。应用Quantity One software (Bio-Rad Inc)分析电泳结果并进行灰度扫描。以目的基因与内参照(β-actin)的灰度值比来表示样本中目的基因mRNA产物的相对表达量。

表1 各个基因引物序列
1.5Western印迹检测 BMSCs在乙醇诱导培养7 d后,提取细胞蛋白,测定浓度,十二烷基硫酸钠-聚丙烯酰胺(SDS-PAGE)凝胶电泳,分离胶浓度为12%,样品进入分离胶前80 V电泳,然后用120 V电泳至样品前沿距凝胶下端1 cm左右。用甲醇浸泡适当大小的聚偏氟乙烯(PVDF)膜5 min,再用转移缓冲液(pH8.3,25 mmol/L Tris-HCl,192 mmol/L甘氨酸,20%甲醇)浸泡10 min,然后将PAGE凝胶上的蛋白质通过电转移印迹到PVDF膜上,100 V转移70 min。用5%牛血清白蛋白/PBS液封闭PVDF膜4℃过夜;加1∶2 000稀释的相应一抗,室温3 min,0.05%吐温20 PBS液洗膜10 min/次,共3次,然后用1∶8 000稀释的山羊抗小鼠IgG二抗室温3 h,0.05%吐温20 PBS液洗膜10 min/次,共3次。电化学发光(ECL)试剂显色:等比例混合ECL试剂盒中的A液、B液,与膜反应1 min,凝胶成像系统显影,照相。结果用Image J version 1.44 software对胶片扫描并进行平均密度值(ADV)测定,与β-actin的比值作为各组产物相对吸光度值。
1.6统计学分析 采用SPSS20.0软件进行t检验、方差分析、q检验及χ2检验。
2 结 果
2.1BMSCs的流式细胞技术分析 BMSCs表面抗原CD90(89.03%)、CD105(90.90%)、CD73(95.03%)均呈阳性表达;CD13(0.83%)、CD34(1.09%)、CD45(1.29%)基本无表达。见图1。
2.2各组脂肪细胞计数、三酰甘油含量、PPARγ基因和蛋白表达比较 模型组和空siRNA组脂肪细胞计数和三酰甘油含量明显高于对照组(P<0.01),PPARγ干扰后或加入T0070907后,PPARγ组和T0070907组的脂肪细胞计数和三酰甘油含量明显降低(P<0.01),与对照组差异无统计学意义(P>0.05)。模型组和空siRNA组PPARγ基因和蛋白表达水平明显高于对照组(P<0.01),用PPARγ干扰后或加入T0070907后,PPARγ组和T0070907组PPARγ基因或蛋白表达水平明显降低(P<0.01),而与对照组差异无统计学意义(P>0.05)。见表2。

图1 各组PPARγ基因和蛋白表达量比较
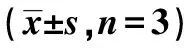
表2 各组脂肪细胞计数、三酰甘油含量、PPARγ基因和蛋白表达比较
与对照组比较:1)P<0.01;与模型组和空siRNA组比较:2)P<0.01;下表同
2.3各组成脂基因转录水平比较 模型组和空siRNA组Ap2、LPL、FATP1和FAT的基因表达水平明显高于对照组(P<0.01),用PPARγ干扰后或加入T0070907后,PPARγ组和T0070907组成脂基因表达水平明显降低(P<0.01),与对照差异无统计学意义(P>0.05)。见表3。
表3 各组成脂基因转录水平比较
3 讨 论
早在130年前德国病理学家Cohnheim证实骨髓中存在非造血实质细胞的干细胞存在,后来实验证实BMSCs是具有高度自我复制能力和多向分化潜能的细胞,不仅可以分化为软骨细胞、成骨细胞、脂肪细胞、肌肉细胞和造血细胞,同样也能分化为神经细胞如神经元、星形胶质细胞等〔3,4〕。对于BMSCs特征的鉴定还没找到理想的标志物,只能根据细胞贴壁后期表面的蛋白标记如CD44、CD13、CD105、CD59、CD166和HLA-ABC表达阳性;CD34、CD80、CD45、CD117和HLA-DR阴性来进行判断;但这些表面抗原并非BMSCs特有,不仅具有间皮细胞的表面抗原特征,同时也具有上皮和内皮细胞表面抗原的特征,因此根据BMSCs培养分化表型、生长规律和生物特性逆推,仍然是一种比较可靠的方法〔3,5〕。本文BMSCs表面抗原表达与文献报道结果类似〔6,7〕。
RNAi具有高效和高特异性等优点,目前在实验研究中广泛运用到基因功能和疾病的治疗。临床和基础实验均认为ANFH疾病是由于骨髓内脂肪积聚和脂肪代谢异常引起的病理生理变化,体外实验证实乙醇可上调BMSCs内PPARγ mRNA表达,并诱导转化为脂肪细胞,最终导致大量的脂肪细胞增殖,股骨头内的骨细胞因脂质沉积,脂肪变性而死亡〔8,9〕。因此从现有的研究可以发现PPARγ可能是引起ANFH疾病的重要靶基因,阻断PPARγ基因的表达,从而阻止乙醇诱导BMSCs的脂肪分化,防止乙醇性ANFH形成具有重要的应用前景〔10,11〕。本研究结果说明siRNA和T0070907治疗取得同样的疗效,而且siRNA和T0070907均能抑制BMSCs的PPARγ基因或蛋白表达,从而阻止向脂肪细胞转化。
Ap2是成熟脂肪细胞形成的重要标志物,位于PPARγ下游,主要对于脂肪的储存和脂肪的分解代谢的靶基因,有研究表明〔11〕肥胖大鼠脂肪组织中的PPARγ和Ap2 mRNA表达明显增强,认为机体的脂肪合成能力明显提高。本研究说明干扰PPARγ基因的表达或T0070907治疗,能够明显降低下游Ap2基因水平的表达。LPL是水解乳糜颗粒和极低密度脂蛋白中三酰甘油的关键酶,通过酶的催化活性调节脂蛋白的代谢,从而达到调节血脂的作用。研究表明LPL是PPARγ下游的重要脂肪调节酶,当机体的PPARγ活性降低时,其机体的LPL的分泌量及酶的活性明显降低〔12〕。FAT最先从小鼠脂肪组织中分离得到,具有转运脂肪酸的作用,被称为脂肪转运酶,对骨骼肌细胞膜上跨膜调节脂肪酸的吸收具有重要作用〔13〕。FATP1有PPARγ靶基因的结合位点,在脂肪细胞中,PPARγ上调FATP1的表达,FAT和FATP1这些调节脂肪酸跨膜转运的蛋白因子,可以相互作用和相互促进,通过协作传递游离的脂肪酸的作用〔14〕。本研究结果说明机体通过干扰PPARγ基因的表达,或对PPARγ表达进行抑制,从而达到降低机体成脂基因的表达,达到治疗的效果。
