刺参神经肽S受体基因的克隆及表达
2018-09-19宋伊敏杨静文闫允君平洪领王天明
宋伊敏,杨静文,闫允君,平洪领,2,王天明
(1.浙江海洋大学海洋科学与技术学院,国家海洋设施养殖工程与技术研究中心,浙江舟山 316022;2.浙江海洋大学海洋与渔业研究所,浙江省海洋水产研究所,浙江舟山 316021)
神经肽S(neuropeptide S,NPS)是于2002年发现的一种活性肽,利用反向药理学证明它是GPR154(G-protein coupled receptor 154,其功能与哮喘、过敏、焦虑及炎症反应相关)的内源性配体[1],后经XU Yanling,et al[2]对其进一步研究将其命名为神经肽S。NPS作为一类肽类神经递质,由20个氨基酸组成,通过激活其受体(NPSreceptor,NPSR)介导下游信号转导[3]。NPSR是一种G蛋白偶联受体(GPCR),主要结合Gs和Gq蛋白;NPS与NPSR结合后,可引起细胞内Ca2+瞬时增加,也可使cAMP水平提高[2],此外,还能激活MAPK信号通路从而激活细胞外调节蛋白激酶(extracellular regulated protein,ERK1/2)[4-5],介导细胞内下游信号的传递。NPSR与其他多肽受体有中等程度的同源性,尤其是血管加压素(vasopressin/oxytocintype,VP/OT-type)受体,但也仅仅有 21%~23%的氨基酸残基相同[6]。XU Yanling,et al[2]发现,NPS 前体肽mRNA在大鼠的脑、甲状腺、唾液腺、乳腺、脊髓、眼、舌、心、肺、小肠、睾丸、食管等多种组织中均有表达,在脑、甲状腺、唾液腺和乳腺表达水平最高,在外周组织有少量分布,而其受体NPSR mRNA则高表达于神经系统内,如运动皮质、前嗅核、梨状皮质、丘脑、下丘脑、杏仁核等,同时在外周组织中亦有广泛表达[7-8]。
NPSR已在人、兔、家猪等高等动物中得到克隆,且生理功能的研究主要集中于大鼠、小鼠等高等模式动物[9-10],在低等动物中的相关研究较少。目前研究表明,NPS系统在抗焦虑恐惧[11],调节动物的觉醒与睡眠[2],调节小鼠恐惧记忆[12-13],影响摄食行为[14-16],调节和参与成瘾过程[17-18]等诸多生理功能方面都有作用。先前的研究认为NPS及其受体仅存在于四足动物中(包括哺乳动物、鸟类、爬行动物和两栖动物),然而后续更广泛的系统发育分析显示,NPSR同源蛋白也存在于无脊椎动物中。例如,在果蝇Drosophila sp和其他昆虫中,NPS/NPSR同源基因编码的蛋白主要参与控制其蜕皮过程,被称为甲壳类心动肽(crustacean cardioactive peptide,CCAP)及其受体(crustacean cardioactive peptide receptor,CCAPR)[19-20]。因后口无脊椎动物中,NPS的同源肽在N端有着共同的Asn-Gly(NG)基序,也被统称为NG肽。由于NPS/NPSR基因在进化中出现较晚,且NPS系统广泛分布于中枢神经系统及外周组织,执行众多生理功能,因此还需要大量的工作来探究其功能特征。
刺参Apostichopus japonicus隶属于棘皮动物门Echinodermata、海参纲Holothuroidea、楯手目Aspidochirota、刺参科Stichopodidae[21],是中国北方重要的海水养殖种类,营养价值高且药用价值广泛。海参在水温上升至20℃以上会向深水移动,藏于岩石下,不动不食,进入“夏眠”[22]。在此期间,刺参停止摄食,基础代谢下降,消化道退化,体重减轻[23]。刺参夏眠期间,机体消耗自身组织维持基本的能量需求,体质量下降30%~50%,从而严重影响其养殖产量[24-25]。刺参的夏眠涉及到复杂的生理过程,目前对其发生机理及调控研究不断深入,随着刺参全基因组数据的发布,大量基础序列信息为刺参生物学研究提供极大支持[26];从神经内分泌调控角度出发,探讨相关神经激素及递质系统在夏眠等生理过程中的作用机制,将有助于进一步阐明刺参的夏眠机制。
本研究克隆了刺参NPSR-like基因(AjNPSR-like)的全长cDNA序列,利用生物信息学和细胞生物学的方法对AjNPSR-like的受体结构特点进行了预测鉴定,分析了GFP融合表达蛋白AjNPSR-like-EGFP的亚细胞定位特征,检测了其在刺参各个组织和时期的表达情况。为进一步研究NPS/NPSR系统在刺参夏眠期间摄食、代谢调控中的作用机制以及在无脊椎动物中的生理调控作用及进化演变提供支持。
1 材料与方法
1.1 实验材料
本研究所用非夏眠刺参于2016年4月取自山东青岛刺参胶南养殖场,采捕刺参时池内水温为12.35°C,体质量为(79.7±4.7)g;低温运输至实验室,暂养驯化 1 周,池内水温控制在(16±0.5)°C,盐度为 31.0±0.5,24 h不间断充气,每日按时投喂人工配合饲料;活体解剖获取刺参组织(呼吸树、消化道、体壁、肌肉、神经环),液氮冷冻保存。非夏眠刺参取样后,剩余刺参一直饲养至2016年9月,不控温自然升至25°C以上,解剖观察消化道退化成直径约1 mm的细线[27],采集各组织样品后液氮冷冻保存。
实验主要采用试剂为 Trizol(Invitrogen)、三氯甲烷(国药),异丙醇(国药)、DEPC水(碧云天),RQ1RNase-Free DNase(Promega)、M-MLV 逆转录酶(Promega)、琼脂糖(SunShineBio)、RNase-Free RNase inhibitor(Promega)、DNA Maker DL 2000(TaKaRa)、5xLoading Buffer(TaKaRa)、GelRedTM10000X in water(BLOTIUM)、0.25%胰蛋白酶(吉诺生物)、50×TAE(上海生工)、GoTaq Green Master Mix(Promega)、SMARTScribe Reverse Transcriptase(Clonetech)、TIANgel Mini Gel Extraction Kit(Tiangen)、pMD 18-T Vector(TaKaRa)、TOP10 感受态细胞(Tiangen)、引物合成及基因测序(Invitrogen)、L-Glutamine、opti-MEM(Gibico)、T4DNA 连接酶(TaKaRa)、pEGFP-N1 表达载体(Clontech)、pMD18-T(Takara)、限制性核酸内切酶(TaKaRa)、X-treme GENE HP DNA Transfection Regant(Roche)、HEK 293 cell line、膜荧光探针DiI(碧云天)、细胞核染料DAPI(碧云天)、盖玻片(碧云天)、SYBR Green Master Mix(TaKaRa)。
1.2 RNA提取和cDNA合成
将上述5种组织样品从液氮中取出,研磨后取30 mg样品,加入1 mL Trizol,用力振荡、裂解;加200 μL氯仿,振荡15 s室温放置5 min;置于冷冻离心机,12 000×g,4°C,离心15 min;离心后,用新离心管吸取400 μL上清水相,再加400 μL异丙醇,完全混匀于-20°C静置15 min以上;4°C下12 000×g离心15 min,使RNA沉淀;吸掉上清液,后加1 mL 75%的乙醇(75%的乙醇现配并-20°C预冷)洗涤RNA沉淀,12 000×g,4°C,离心10 min;弃上清液,重复洗涤1次;去上清液后离心1 min,吸去剩余液体,干燥RNA沉淀10 min;加30 μL DEPC水使RNA沉淀溶解,获得总RNA样品,经琼脂糖电泳检查RNA的完整性,NanoDrop 2000分光光度计检测RNA的浓度,检测后的RNA放在-80℃冰箱中保存。
1.3 刺参NPSR-like基因cDNA全长克隆及序列分析
根据GenBank中已知的刺参NPSR-like基因序列,利用Primer5.0软件设计NPSR基因的核心片段引物(表1);按照FirstChoiceRLM-RACE Kit试剂盒说明书(赛默飞世尔)设计3'-RACE和5'-RACE引物。将已经反转好并保存的刺参cDNA作为模板,用NPSR-F和NPSR-R引物进行AjNPSR-like基因核心片段的扩增。同时设计3'-RACE和5'-RACE特异性引物(表1),进行3'和5'末端扩增。扩增产物经凝胶电泳检测后按照胶回收试剂盒说明书切胶回收,TA克隆,筛选阳性克隆菌落送公司测序。
在 NCBI上利用在线 ORF Finder(http:www.ncbi.nlm.gov/orffinder/)查找开放阅读框(ORF);用 Net-Phos2.0Server(http://www.cbsdtu.dk/services/NetPhos/)预测蛋白质磷酸化位点;用ScanProsite tool(http://prosite.expasy.org/scanprosite/)预测二硫键位置;Signal4.1(http://www.cbs.dtu.dk/services/SignalP/)预测信号肽序列;用PredictProtein(http://www.predictprotein.org/)预测蛋白质的二级结构;用SWISS-MODEL(http://swissmodel.expasy.org/)预测蛋白质的三级结构;用 InterPro(http://www.ebi.ac.uk/interpro/scan.html)预测蛋白功能结构域;用TMpred(http://www.ch.embnet.Org/software/TMPREDform.html)预测蛋白跨膜结构域;氨基酸序列多重比对用ClustalX软件;采用邻位法(Neighbor-Joining)用MEGA 6.0软件构建系统进化树,1 000次bootstraps,其余参数使用默认值。
1.4 AjNPSR-like在细胞中的定位
为探究AjNPSR-like的亚细胞定位,通过构建表达载体pEGFP-N1,获得AjNPSR-EGFP的融合蛋白表达载体并通过测序验证载体准确性,构建好的载体命名为pAjNPSR-EGFP。所用HEK293细胞采用DMEM高糖完全培养基培养(DMEM高糖培养基+10%胎牛血清+1%双抗)。细胞培养在37°C、5%CO2、湿度衡定的恒温培养箱(Thermo,USA)中进行。使用X-treme GENE HP DNA Transfection Reagent(Roche)按其说明将AjNPSR-EGFP转染至HEK293细胞,12~16 h后,将瞬时转染AjNPSR-EGFP的HEK293细胞经胰蛋白酶处理后接种于载玻片上。24 h后,膜荧光探针DiI染膜,细胞核染料DAPI染核,多聚甲醛固定后封片,置于激光扫描共聚焦显微镜(Leica TCS SP5II)下检测荧光信号分布情况。

表1 刺参NPSR-like基因克隆与表达分析的引物Tab.1 Primers designed for cloning and expression analysis of AjNPSR-like gene from A.japonicus
1.5 AjNPSR-like基因表达特征分析
用实时荧光定量PCR方法,以刺参β-tubulin基因[28]作为实验的内参基因,检测分析不同组织和夏眠刺参AjNPSR-like基因的表达特征。其扩增体系为:cDNA模板0.4 μL,2×SYBR Premix Ex TaqTMII 5 μL,上下游引物各 0.4 μL(表 1),ROX Reference DyeII 0.2 μL,ddH2O 补足至 10 μL。反应程序:94 ℃ 40 s;95℃ 10 s,60℃30 s,40个循环;55~95℃逐渐升温2 min。实验进行3次重复。
标准曲线和CT值等由ABI7500 Real Time PCR仪自带的软件完成。将荧光定量PCR检测结果导出,采用2-△CT法,用SPSS 17.0软件分析。用Duncan法多重比较检验组间差异的显著性,P<0.05检查差异显著。相关柱形图用软件Origin 8.0来制作。
2 结果
2.1 AjNPSR-like基因全长cDNA序列分析
AjNPSR-like基因的cDNA全长2 498 bp(GeneBank登录号:MG199219),开放阅读框1 272 bp,编码423 个氨基酸,5'非编码区(5'UTR)716 bp,3'非编码区(3'UTR)510 bp。预测 AjNPSR-like蛋白的理论分子量为47.675 ku,理论等电点为9.45。ScanProsite tool预测二硫键位置在C112和 C187,有7次跨膜区;通过SignalP 4.1 Server分析发现没有信号肽酶切位点;同时含有1个糖基化位点,46个磷酸化位点(图1)。在线预测其3D结构发现具有7段跨膜结构域(图2-a),其二级结构预测亦同样表明有7段跨膜结构域(图 2-b)。
2.2 AjNPSR-like同源比对与系统进化树分析
通过NCBI BLAST软件对AjNPSR-like预测蛋白序列在线比对,结果显示该序列与紫海胆Strongylocentrotus purpuratus NPS/CCAP-type(CCAP:crustacean cardioactive peptide)受体(SpNPSR)的相似性最高,为47%。多序列比对显示AjNPSR-like与其他物种NPSR/CCAPR有中等程度的同源性(图3)。
系统进化树显示(图4),NPSR及其同源基因CCAPR具有明显的进化分布特征,脊椎动物中为NPSR聚为一支;无脊椎动物中主要聚为两支,一支为后口动物的棘皮动物,包括海百合Antedon mediterranea、海星Asterias rubens、刺参、紫海胆,一支为原口动物的软体动物、节肢动物等,包括长牡蛎Crassostrea gigas、光滑双脐螺Biomphalaria glabrata,温室希蛛Parasteatoda tepidariorum、埃及伊蚊Aedes aegypti、黑腹果蝇Drosophila melanogaster、赤拟谷盗Tribolium castaneum、大红斑蝶Danaus plexippus、二化螟Chilo sup-pressalis和棉铃虫Helicoverpa armigera,主要命名为CCAPR。刺参AjNPSR-like与紫海胆NPS/CCAP-type受体、海星NPS/CCAP-type受体的进化关系最近,之后与海百合NPS/CCAP-type受体聚为另一分支,与其他脊椎动物NPSR聚为另一大支。该进化树分析结果展示了棘皮动物中NPSR或其同源受体具有较高的相似性,并与其进化地位保持一致。

图1 刺参NPSR-like基因全长cDNA序列和对应的氨基酸序列Fig.1 Full-length cDNA sequence and corresponding amino sequence of NPSR-like gene of A.japonicus

图2 预测AjNPSR-like其蛋白结构和跨膜区域Fig.2 Predicted AjNPSR-like protein structure and domain organization
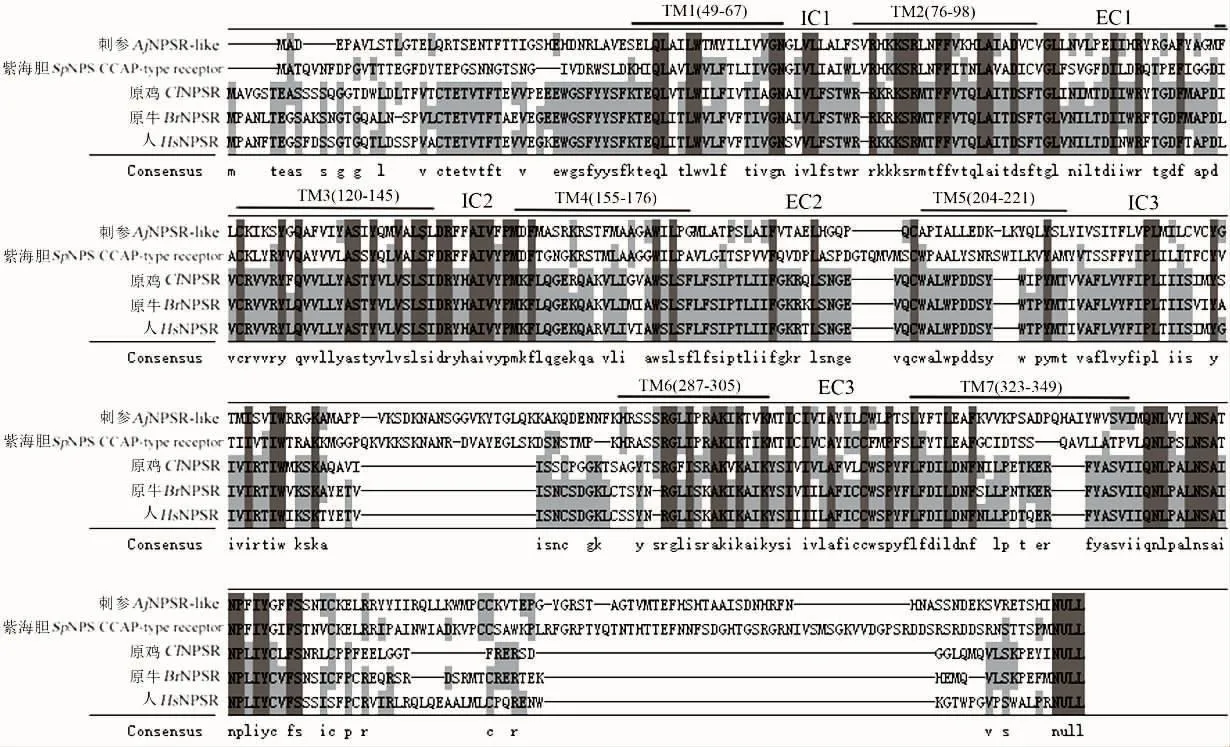
图3 AjNPSR-like氨基酸序列与其他物种同源蛋白的多序列比对Fig.3 Multialignment of AjNPSR-like with homologies from other species

图4 根据AjNPSR-like氨基酸序列构建的NJ系统进化树Fig.4 NJ phylogenetic tree analysis of AjNPSR-like amino acid sequence from A.japonicus and other species
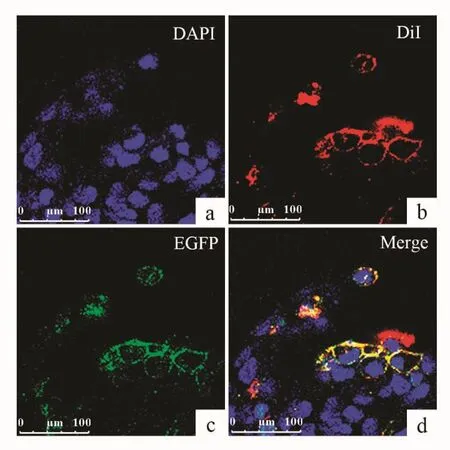
图5 融合蛋白AjNPSR-EGFP在HEK293细胞中的表达Fig.5 Confocal microscopy of HEK293 cells expressing the AjNPSREGFP fusion protein
2.3 AjNPSR-like在HEK293细胞中的亚细胞定位
为分析AjNPSR-like亚细胞定位,本研究将构建的pAjNPSR-like-EGFP的融合表达质粒瞬时转染至HEK293细胞,24 h后,多聚甲醛固定,用DiI染膜,DAPI染核,后封片,置于激光共聚焦扫描显微镜观察。观察显示,在成功转染pAjNPSR-like-EGFP的HEK293细胞中,能观察到明显的荧光,且细胞形态良好,转染成功的荧光主要分布在细胞膜上,与膜染料DiI荧光信号重叠(图5)。结果证明pAjNPSR-like-EGFP受体融合蛋白定位于细胞膜,其C端EGFP标记不影响AjNPSR-like蛋白表达和膜定位,证明本研究克隆得到的AjNPSR-like是跨膜蛋白,这与基于生物信息学预测的A-jNPSR-like属于G蛋白偶联受体家族结果一致。
2.4 AjNPSR-like基因在刺参成体不同组织中的表达差异分析
AjNPSR-like mRNA在非夏眠刺参呼吸树、消化道、体壁、肌肉、神经环等5个组织中均有表达,但表达水平有所差异,在肌肉中表达量最高,其次为体壁和神经环;在夏眠期间,刺参各组织中的AjNPSR-like mRNA表达水平显著上升,且在消化道、体壁、肌肉和神经环中AjNPSR-like的表达极显著高于非夏眠期间A-jNPSR-like的表达量(图6)。

图6 AjNPSR-like基因在5个组织中的表达差异分析(n=3)Fig.6 Expression level of AjNPSR-like mRNA in five tissues of A.japonicus
3 讨论
本研究通过RACE技术克隆获得刺参AjNPSR-like基因的cDNA序列的全长为2 498 bp,编码423个氨基酸,预测蛋白结构分析其具有GPCR家族典型的7次跨膜螺旋结构(7 TM helix),这些TM区在受体活化、磷酸化、受体表达、信号传递等方面起重要作用,跨膜蛋白在细胞中常以离子通道形式存在,执行信号传导或物质转运功能[29]。其氨基酸序列与紫海胆NPS/CCAP受体相似性最高,为47%,这与其进化地位一致。BERNIER,et al[30]研究了NPS结构和功能的关系,发现NPS N端1/3的氨基酸残基对受体的激活是必须的;前 6个氨基酸特别是第 2(Phe,苯丙氨酸)、3(Arg,精氨酸)、4(Asn,天冬氨酸)和 6个氨基酸残基(Val,缬氨酸)是激活受体所必需的。NPS与NPSR结合后,可引起细胞内Ca2+瞬时增加,也可使细胞内cAMP水平升高,表明NPS偶联的主要是Gs和Gq蛋白[31]。刺参NPSR-like氨基酸序列与多个物种的NPSR及同源基因CCAPR氨基酸序列进行系统发生分析,显示刺参NPSR-like与棘皮动物的NPSR/CCAPR聚类,并与脊椎动物的NPSR家族和原口动物的软体动物和节肢动物的CCAPR家族各自聚成一支,这一方面说明NPSR/CCAPR系统在进化过程中经历了明显的演变过程,亦说明深入开展棘皮动物NPSR/CCAPR具有重要的基础理论价值。
目前研究认为,棘皮动物NPSR/NPSR-like与无脊椎动物心脏加速肽受体(CCAPR)同源;且NPSR/NPSR-like/CCAPR这组受体与脊椎动物、非脊椎动物血管加压素样受体(VP/OT-type receptor)系统发育关系最为密切[32]。已有研究表明,棘皮动物中存在的NG肽,一方面与无脊椎动物中的NPS有着共同的Asn-Gly基序,另一方面NG肽的前体肽具有激素运载蛋白结构域,而这种结构域在过去的研究中认为是VP/OT-type(Vasopressin/Oxytocin-type)肽的前体肽中所特有的,因此棘皮动物中存在的NG肽及其受体可以被看做是进化过程中连接NPS/NPSR系统和VP/OT-type肽及其受体的桥梁[33-34]。在进化过程中,很可能是VP/OT系统在基因的加倍过程中,一个拷贝保持了VP/OT系统在演化中的稳定,另一个拷贝则发生了改变从而演化出无脊椎后口动物中的NG肽系统和原口动物的CCAP系统[32]。目前紫海胆中脊椎动物NPSR的同源受体被克隆和鉴定,被称为NPS/CCAP-type受体,它能够被紫海胆的一种NG肽:NGFFFamide所激活,该神经肽由包含运载蛋白的前体蛋白衍生而来[35]。这一发现将脊椎动物中的NPS型受体、无脊椎动物中的NG肽以及CCAP型受体联系起来。紫海胆NGFFFamide受体是第一个以棘皮动物为特征的NPSR-type受体。本研究后续的工作将使用刺参中的一种候选NG肽:NGIWYamide来探究其对A-jNPSR-like的激活情况,从而进一步揭示其在系统进化方面具有的重要意义。
实时荧光定量数据显示,AjNPSR-like mRNA在外周组织中均有分布,这与XU Yanling,et al[2]在大鼠中的研究结果相似,说明AjNPSR-like在刺参中可能也参与多种生理活动的调控。有研究人员发现,NPSR可以抑制大鼠、小鼠、鸡的摄食[14-15,36],也有研究报道NPSR能促进大鼠的进食[16]。NPSR在刺参夏眠阶段消化道中的表达量明显提高,提示可能抑制了夏眠期间刺参的摄食。另外,有研究指出,NPS通过激活其受体NPSR可抑制小鼠杏仁核和额叶皮质5-羟色胺(5-HT)的释放[37],而5-HT作为重要的单胺类神经激素,在动物的摄食调控中发挥着重要作用,与刺参夏眠代谢调控存在相互作用[38]。在运动方面,NPS通过激活NPSR调节基底神经节的活动从而发挥增强运动水平的作用[39]。但由于刺参属于无脊椎动物,缺乏高级的中枢神经系统,因此需要进一步的研究来阐明NPSR是如何通过其神经系统来调节摄食和代谢的,特别是其在刺参节律行为的调控作用方面的作用研究亦值得关注[40]。目前对于NPSR的体内体外研究主要集中于啮齿类动物(大鼠、小鼠、兔)[41-42]及哺乳动物(人、猪)[10,43]上,对于无脊椎动物NPSR的组织表达、生理功能等研究报道较少,因此有必要开展这方面研究,进一步阐明NPSR系统在物种进化中的演变,并为研究NPSR在棘皮动物等低等动物中的功能及生理调控机理提供理论基础。
综上所述,本研究对刺参AjNPSR-like的基因全长及其编码蛋白进行了特征分析,确定了AjNPSR-like受体蛋白的亚细胞定位特征,研究了其在刺参不同组织中的转录表达水平及夏眠期间的高表达特征,探讨了NPS/NPSR系统可能在刺参生理调控中的作用,为进一步探究该系统的生理调控功能及进化演变奠定基础。
