中国沿海新报道的三种产毒拟菱形藻
2018-09-13董焕嫦黄春秀
董焕嫦 黄春秀 李 扬
(1. 华南师范大学生命科学学院,广州市亚热带生物多样性与环境生物监测重点实验室,广州 510631; 2. 华南师范大学生命科学学院,广东省水产健康安全养殖重点实验室,广州 510631)
拟菱形藻属(Pseudo-nitzschiaPeragallo)物种是全球近岸水域广布的浮游硅藻, 因部分物种能够产生记忆缺失性贝毒(Amnesic Shellfish Poisoning,ASP)—多莫酸(Domoic acid, DA)而备受关注[1,2]。目前全球已报道49个拟菱形藻物种, 其中26种能够产生多莫酸[3,4]。另一方面, 拟菱形藻产生多莫酸的特征又是不稳定的, 如中国海域的福氏拟菱形藻P. fukuyoiLim, Teng, Leaw & Lim能够产毒[5], 但是其马来西亚株系却是无毒的[6]。
以往研究已经证实拟菱形藻在我国沿海广泛分布[7,8], 但相关研究还相对有限。截止目前, 我国报道的拟菱形藻物种有24个[4,5,8—10], 但对于部分物种的认知还仅停留在形态学水平, 现代分子生物学技术的引入还十分有限, 造成物种信息不全、鉴定结论不可靠, 因此目前对于我国海域拟菱形藻属物种多样性还难以有清晰的认识。另外, 关于我国海域的产毒拟菱形藻, 近两年才陆续报道了2个物种:伪装拟菱形藻P. simulansLi, Huang & Xu[4]和福氏拟菱形藻P. fukuyoi[5]。但相对于我国辽阔的近海海域而言, 仅有2种产毒拟菱形藻显然是不切实际的。近年我国多处海域的海产品中已检出DA[11—13],表明中国海域DA的分布是广泛的, 其DA生物来源也应该是多样的。为了明确我国海域产毒拟菱形藻的物种多样性, 本文基于建立的单克隆培养株系,开展了种源信息和产毒特征的工作。
1 材料与方法
1.1 单克隆藻株的建立
用浮游植物网(孔径10 μm)进行水平拖网, 采集活体水样, 尽快带回实验室。利用毛细管复洗法在倒置显微镜(Mshot MI-12)下分离目标藻细胞, 经过多洗水洗和转移, 以确保单克隆藻株的纯化。最后转移至预先滴有培养基的48孔细胞培养板中, 以普通荧光灯为光源, 光周期12h∶12h, 温度(20±2)℃的条件下培养。待其存活并繁殖达到约100个藻细胞之后, 转移到盛有培养基的100 mL锥形瓶中培养,以MC (Marine Collection)序列进行编号(表1)。
1.2 卤虫的培养及诱导
购买商品化卤虫卵, 用灭菌人工海水冲洗干净至无气味, 投入经过灭菌处理的L培养基中, 孵化温度等条件与拟菱形藻培养条件一致。约2d后, 将10—20个成体卤虫转移到拟菱形藻培养液中(细胞密度为106cell/L)。放置在光周期12h∶12h, 温度在(20±2)℃的条件下混合培养约7—8d[5]。
1.3 LC-MS/MS法检测藻毒素
取处于生长稳定期中后期的藻液400 mL左右,经0.22 μm醋酸纤维滤膜过滤收集藻细胞。用5 mL甲醇水(1∶1)将滤膜上的藻细胞洗脱到离心管中。将藻液用超声波破碎仪冰浴破碎3—5min, 经0.22 μm滤膜过滤, 滤液于–20℃下保存备用。样品送至农业部水产品质量安全检测与评价重点实验室检测DA含量。
采用Prominence UFLC超快速液相色谱(Shimadzu公司)和5500 QTRAP四极杆-线性离子阱复合质谱检测系统(SCIEX公司)对预处理的样品进行DA检测, 参见Wu等[14]方法进行分析。DA标准品购自德国Sigma公司。
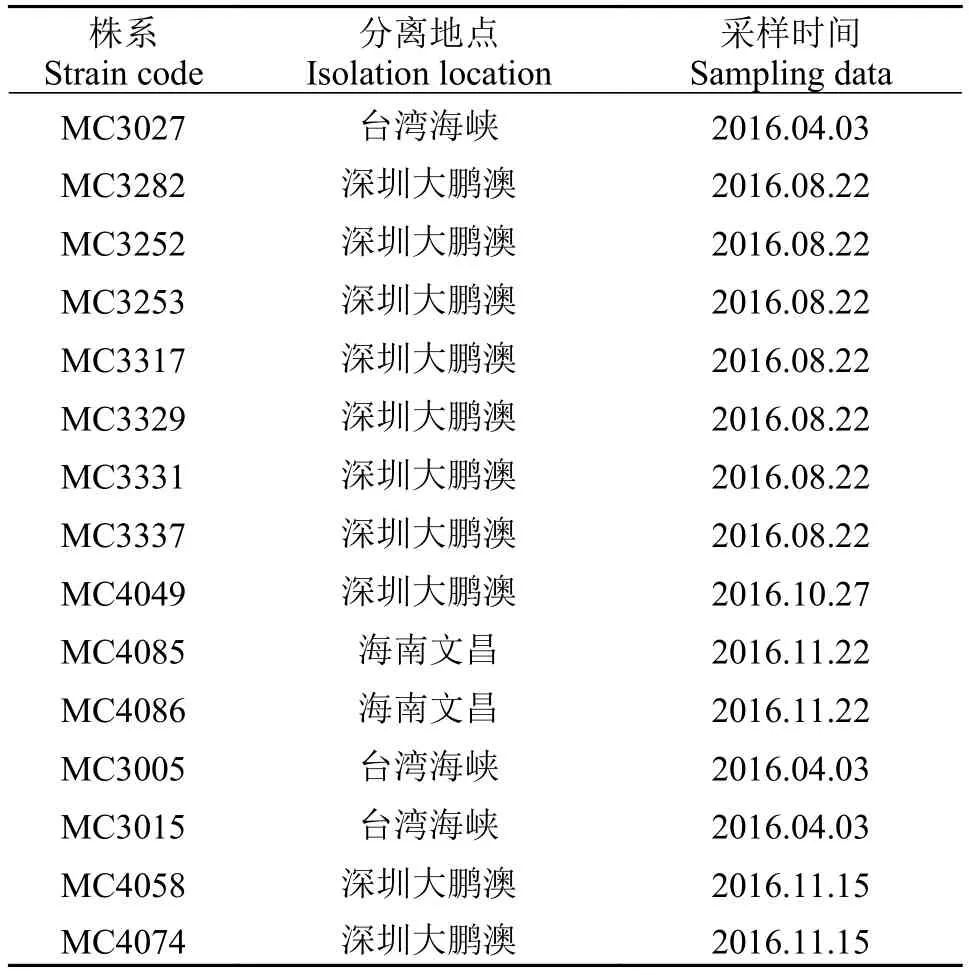
表1 本文建立的拟菱形藻单克隆培养株系Tab. 1 List of monoclonal Pseudo-nitzschia strains investigated in this study
1.4 形态学观察
光学显微镜(Light Microscopy, LM)观察: 取处于对数生长期的藻液0.1 mL, 滴在载玻片上, 盖上玻片后, 利用Olympus BX53进行微分干涉(Differential Interference Contrast, DIC)的观察, 并使用Olympus DP27数码相机拍照, 在Olympus Cell-Sens软件上获取图像信息。主要观察群体特征、细胞色素体形态等。
透射电子显微镜(Transmission Electron Microscopy, TEM)观察: 取对数生长期的藻液2 mL, 加入等量浓硫酸(>95%)以去除有机质, 然后用蒸馏水多次水洗至中性[15]。用微量进样器吸取5—10 μL酸化后的水样, 滴加在喷镀碳膜的铜网(100目)上, 自然晾干后, 即可在JEM-1010 TEM下观察和拍照。主要观察壳面超微结构, 如点条纹、肋突、孔纹等。
1.5 分子系统学分析
离心法收集处于对数生长期的藻细胞, 进行总DNA的提取[16]。利用引物ITS1和ITS4进行核糖体转录间隔区(ITS)的扩增和测序[6]。PCR产物送至上海英潍捷基生物技术有限公司进行纯化和测序。从NCBI下载拟菱形藻物种的ITS nrDNA序列, 使用BioEdit软件进行序列的比对和矩阵[17]。利用Mr-Modeltest 2.3计算选择贝叶斯推理树最适模型[18]。用MrBayes 3.2[19]构建贝叶斯推理树(Bayesian Inference, BI), 用RAxML-HPC2[20]构建最大似然树(Maximum Likelihood, ML), 其中ML分析的自检值(bootstrap test)设定为1000。
2 结果
2.1 多莫酸的LC-MS/MS检测
以DA标准品浓度(ng/mL)为横坐标, 峰面积为纵坐标, 建立DA的LC-MS/MS检测的标准曲线,DA浓度为10—1000 ng/mL时, 其峰面积与质量浓度有良好的线性关系(R2=0.99934), 回归方程为:y=164.64875x–3947.22457。本方法的检测下限为10 ng/mL。DA的出峰保留时间为0.77min。分别对纯种培养株系和卤虫混培诱导株系进行检测, 详细检出结果见表2。
本文建立的15个纯种培养藻株中, 6个株系未检出DA, 其余9个株系检出DA, 单细胞产毒量0.4—17.5 fg。
挑选了3个株系进行卤虫混培下的DA诱导,MC4049与MC3015的产毒水平略有下降, 单细胞产毒水平分别由2和1 fg降至0.2和0.4 fg, 而MC4074的产毒特征则有明显变化, 单细胞产毒水平为17.5 fg,而其纯种株系并未有DA检出。
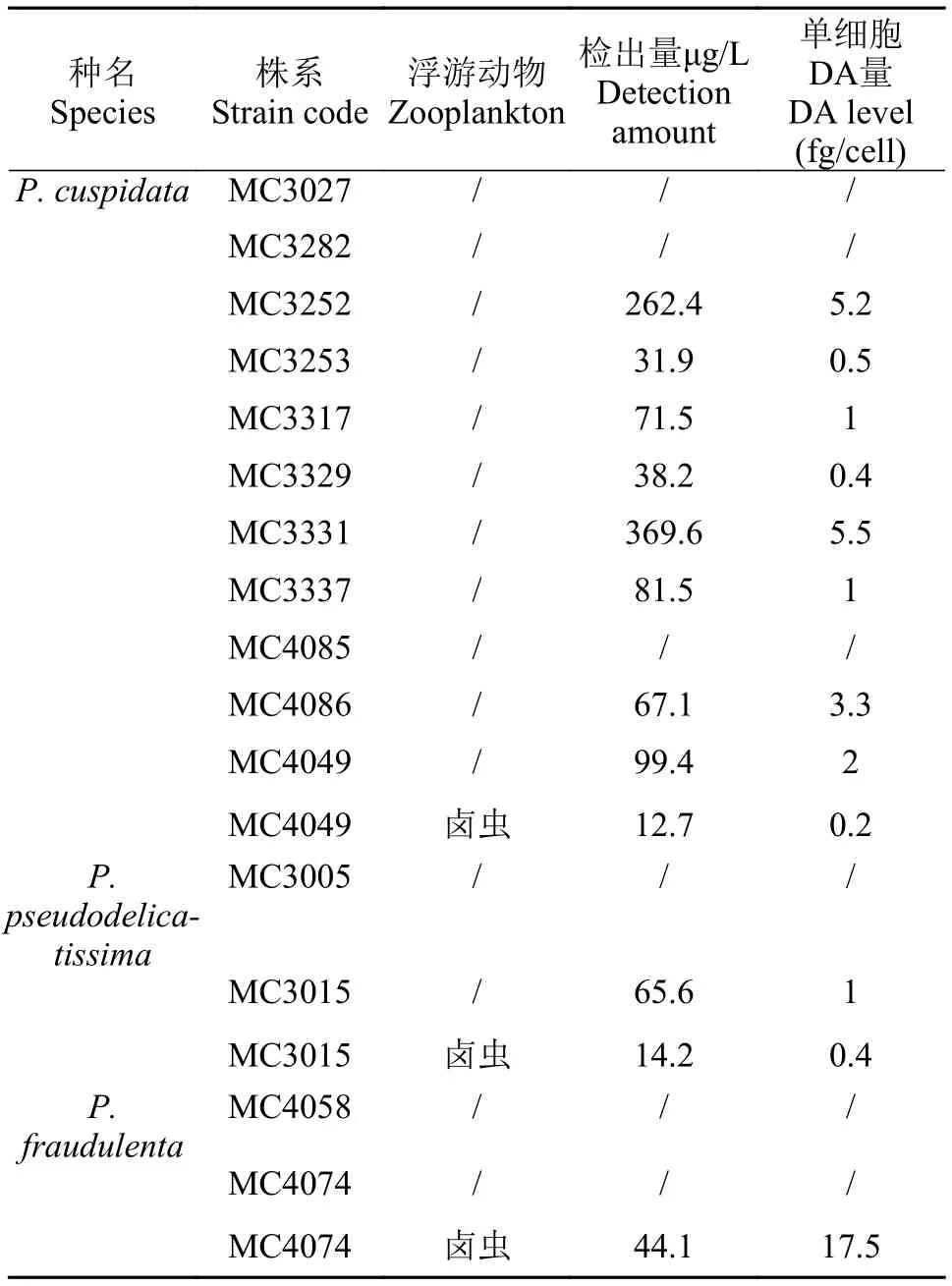
表2 本文的DA检出结果Tab. 2 The results of DA’s content detected in this study
2.2 产毒拟菱形藻的物种鉴定
基于形态学和分子系统学的数据, 对10个产毒株系进行了物种鉴定, 确认它们分别隶属于3个物种: 尖细拟菱形藻P. cuspidata(Hasle) Hasle、伪柔弱拟菱形藻P. pseudodelicatissima(Hasle) Hasle、伪善拟菱形藻P. fraudulenta(Cleve) Hasle, 其中伪善拟菱形藻是我国的新记录种。
尖细拟菱形藻Pseudo-nitzschia cuspidata(Hasle) Hasle(图版 Ⅰ) 细胞可形成阶梯状群体, 重叠部分占壳面全长的1/6—1/8左右(图版 Ⅰ-1)。壳面披针形(图版 Ⅰ-2)。长41—43 μm, 宽1.6—1.9 μm。壳端尖(图版 Ⅰ-3), 管壳缝强烈偏心,具有中央较大船骨点(图版Ⅰ-4)。肋突分布均匀,10 μm内20—22条。点条纹由一排椭圆形的孔纹组成(图版 Ⅰ-5), 密度为40—43条/10 μm。孔纹密度为7个/μm。孔纹多分成两半(图版 Ⅰ-5), 也有分成2—5个部分的(图版 Ⅰ-4)。本文只观察到二条环带(图版 Ⅰ-6), 壳环带高一个孔纹, 点条纹密度为56—57条/10 μm。
产毒特征: 本种在美国西海岸和澳大利亚均有产毒报道, 单细胞产毒量分别为6.3×104fg[21]和4.3×103—2.5×104fg[22]。本文株系亦有DA检出, 单细胞产毒水平为0.5—5.5 fg。
生态: 海水浮游生活。
分布: 全球广布种, 澳大利亚、墨西哥湾、西班牙、美国西海岸均有分布[21—25]。本文标本采自广东大亚湾(8月、10月)和海南文昌(11月), 共建立11个培养株系(表2)。
伪柔弱拟菱形藻Pseudo-nitzschia pseudodelicatissima (Hasle) Hasle(图版 Ⅱ) 细胞可形成阶梯状群体, 重叠部分占壳面全长的1/5—1/6(图版Ⅱ-1)。壳面披针形(图版 Ⅱ-2), 壳端尖(图版 Ⅱ-3)。壳面长52—61 μm, 宽1.4—1.6 μm。管壳缝强烈偏心, 具有中央较大船骨点(图版 Ⅱ-4)。肋突密度为19—24条/10 μm。点条纹由一排孔纹组成, 密度为39—43条/10 μm。孔纹圆形或椭圆形, 多分成两部分(图版 Ⅱ-5), 6—7个/μm。壳环带高一排孔纹(图版 Ⅱ-6)。第二条环带孔纹椭圆形(图版 Ⅱ-7),环带上孔纹密度为48—53条/10 μm。
毒性: 本种在地中海曾有产毒报道, 单细胞产毒量为4.5—9.3 fg[26], 本文株系亦有DA检出, 单细胞产毒水平为1 fg。
生态: 海水浮游生活。
分布: 全球广布种, 澳大利亚、丹麦、墨西哥、新西兰、美国西海岸等有分布[23,26,27]。本文标本采自台湾海峡(4月), 共建立2个培养株系(表2)。
伪善拟菱形藻Pseudo-nitzschia fraudulenta(Cleve) Hasle(图版 Ⅲ) 细胞可形成阶梯状群体, 重叠部分占壳面全长的1/7(图版 Ⅲ-1)。壳面线形, 两侧对称, 在壳面中部几乎平行(图版 Ⅲ-2)。壳端略尖(图版 Ⅲ-3)。管壳缝强烈偏心, 具有中央较大船骨点(图版 Ⅲ-4)。壳面长60—66 μm, 宽3.9—4.5 μm。肋突分布不均匀, 密度为13—17条/10 μm。点条纹由两排紧密排列的孔纹组成, 密度为25—30条/10 μm。孔纹圆形, 内部被分成3—5个部分(图版 Ⅲ-6)。壳环带高6—7个孔纹, 宽2个孔纹(图版 Ⅲ-5), 环带上点条纹38—45条/10 μm。
毒性: 本种在美国加利福尼亚曾有产毒报道[28]。本文的纯种株系中未有DA检出, 但在卤虫混培下可检出DA, 单细胞产毒水平为17.5 fg。
生态: 海水浮游生活。
分布: 本种在美国西海岸[28]、澳大利亚东南部海域有分布[22]。本文标本采自广东大亚湾(11月),共建立2个培养株系(表2)。
基于ITS序列的分子系统学分析基于ITS1-5.8S-ITS2序列建立了分子系统树, ML树和BPP树的拓扑结构一致(图1)。本文的尖细拟菱形藻株系与以往报道的尖细拟菱形藻墨西哥株系(Mex12)聚在同一分支上, 尖细拟菱形藻8个产毒株系中, 株系MC3252、MC3329和MC3337的ITS基因完全相同, 株系MC3253、MC3317、MC3331和MC4049的ITS基因完全相同, 前者与后者有10个碱基差异。前者株系与Mex12株系有13个碱基差异,后者与Mex12株系有6个碱基差异。两个伪柔弱拟菱形藻株系与伪柔弱拟菱形藻希腊株系(8A10, AL-15)聚在一起, MC3005和MC3015的基因完全一样,与系统发育树上8A10株系有2个碱基差异, 与AL-15株系有2个碱基差异。2个伪善拟菱形藻株系与伪善拟菱形藻西班牙株系(Limens1)、澳大利亚株系(KC017467)聚在同一分支上, MC4058与MC4074碱基完全一样, 与伪善拟菱形藻西班牙株系(Limens1)有9碱基差异, 与澳大利亚株系(KC017467)有20个碱基差异。ITS序列相对高变,同一物种的不同地理株系之间亦会存在差异。系统树显示上述株系间碱基差异并未影响物种的聚类, 属于种内差异, 因此分子鉴定支持形态鉴定的结果。
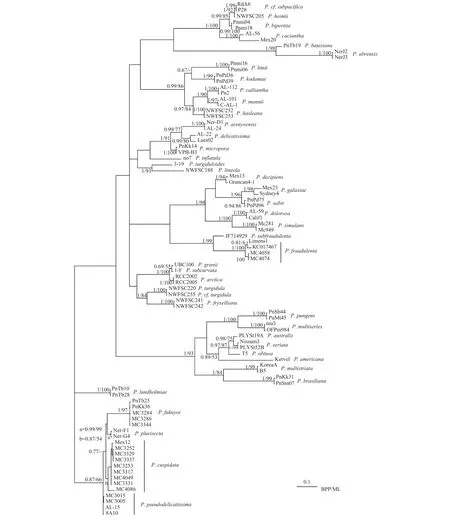
图1 基于核糖体转录间隔区ITS1-5.8S-ITS2的分子系统树Fig. 1 Molecular phylogenetic tree inferred from ITS1-5.8S-ITS2 region sequences
3 讨论
3.1 中国海域产毒拟菱形藻的物种多样性
本文报道的3种产毒拟菱形藻, 曾在国外海域有产毒报道。如采自澳大利亚[22]的5株尖细拟菱形藻, 其中2株检出DA, 单细胞产毒量分别为2.5×104和4.3×103fg。Trainer等[21]在2009年报道了尖细拟菱形藻藻华, 自然海水中DA含量高达43 nmol/L, 单细胞产毒量为6.3×104fg。地中海东北部海域[26]的6株伪柔弱拟菱形藻中, 有3株检出DA, 单细胞产毒量为4.5—9.3 fg。Gárate-Lizárraga等[28]报道了美国加利福尼亚州海湾发生的伪善拟菱形藻藻华, 自然海水中DA含量为52 ng/filter。
在过去的20余年, 我国学者持续关注中国海域拟菱形藻的产毒特征, 但一直未能发现产毒株系。最早的工作开始于2005年, Li等[29]对分离自长江口的一株尖刺拟菱形藻P. pungensHasle进行了检测,未有DA的检出。之后, 邢小丽等[30]、杨军霞[31]、黄迎艳[32]、徐国双和李扬[9]、黄春秀等[10]陆续检测了100多个拟菱形藻株系, 均未有DA的检出。直至近年, 才陆续有中国海域产毒拟菱形藻的报道。Li等[4]在广东沿海发现了1个新种: 伪装拟菱形藻(P. simulans), 并在培养株系中检测到DA, 单细胞产毒水平是1.05—1.54 fg, 这是我国产毒拟菱形藻的首次报道。随后, 黄春秀等[5]报道了1个新记录种-福氏拟菱形藻(P. fukuyoi), 在5株培养株系中均能检测到DA, 单细胞产毒水平为20—1630 fg。截止目前, 我国已有2种产毒拟菱形藻的报道。然而,相关研究仍相对有限, 检测藻株较少, 尚不能全面反映中国海域拟菱形藻的产毒情况。本文报道了3个产毒种, 丰富了我国产毒拟菱形藻的物种多样性, 将其物种数提高为5个。但海产品中DA的检出,显示DA在我国沿海分布广泛[11—13], 产毒拟菱形藻的物种多样性也应该是多样化的, 应该还有更多的产毒种未被认知。
3.2 产毒特征与浮游动物的关系
拟菱形藻的产毒特征是不稳定的, 受到多种环境因素的影响, 如光照、温度、不同生活史阶段、磷、硅、微量元素等[1,2]。近年有研究发现浮游动物摄食会提高某些拟菱形藻物种的产毒水平[33], 甚至能够诱导未曾报道过产毒的拟菱形藻物种开始产生DA[34]。如Harðardóttir等[34]将不产毒的钝头拟菱形藻P. obtusa与桡足类Calanus copepodites混和培养, 发现钝头拟菱形藻开始分泌DA, 桡足类体内富集DA浓度可达到0.4×10−4ng DA/μg C。本文的伪善拟菱形藻(MC4074)在纯种培养条件下并未检出DA, 但在卤虫混培条件下可检出DA, 也验证了这一观点。关于浮游动物诱导拟菱形藻产生DA的机制和生态学意义, 有学者认为桡足类的刺激作用并不是通过与藻细胞直接接触而实现的, 应该是通过释放的某种小分子化学物质来完成, 但化学物质的具体成分尚不明确[33]。另外, 拟菱形藻产生更多DA可以抑制桡足类的摄食, 甚至影响桡足类的产卵率和孵化率, 从而减少藻类被摄食的概率, 具有显著的生态学意义[33]。
本文利用卤虫诱导拟菱形藻产毒, 希望能够提高产毒量, 但是也出现了产毒量下调的现象, 如在卤虫混培条件下, 尖细拟菱形藻(MC4049)和伪柔弱拟菱形藻(MC3015)的产毒水平都有所下降, 单细胞产毒量分别前者从2 fg降低至0.2 fg, 后者从1 fg降至0.4 fg。分析其原因, 或许与拟菱形藻株系的人工保种时间过长有关, 因为随着培养时间的延长,拟菱形藻的产毒能力会快速降低。本文2个株系(MC4049和MC3015)纯种条件下的产毒检测, 是在株系建立后的一个月内完成的, 而浮游动物诱导实验则是在保种半年之后进行的。
3.3 与相似种的比较研究
尖细拟菱形藻在形态上与伪柔弱拟菱形藻较难区分。Lundholm等[23]认为二者细微差异在于外形和壳面观: 伪柔弱拟菱形藻呈线形, 尖细拟菱形藻呈披针形, 而本文报道的伪柔弱拟菱形藻则是披针形。李扬等[35]报道的伪柔弱拟菱形藻的环面观是线形或披针形, 壳面外形并不能作为尖细拟菱形藻与伪柔弱拟菱形藻的重要区别。另外, 本文中尖细拟菱形藻的肋突密度(20—22条/10 μm)、点条纹密度(40—43条/10 μm)与伪柔弱拟菱形藻(肋突密度为19—24条/10 μm, 点条纹密度为39—43条/10 μm)的相接近, 但伪柔弱拟菱形藻(长52—61 μm,宽1.36—1.6 μm)比尖细拟菱形藻(长41—43 μm, 宽1.58—1.9 μm)较细长。在分子系统树上, 尖细拟菱形藻与伪柔弱拟菱形藻的分子亲缘关系也是最为接近的, 但两者的二级结构有4个HCBCs的差异。
伪善拟菱形藻与亚伪善拟菱形藻P. subfraudulenta(Hasle) Hasle的形态相似, 分子亲缘关系最为接近。Hasle等[36]认为两者之间的最大区别是壳面外形, 伪善拟菱形藻是披针形, 亚伪善拟菱形藻则是线形。但是此后的研究发现这两个物种的壳面外形可能是多变的, 兼具披针形和线形, 如美国加利福尼亚州海湾[28]和澳大利亚海域[22]的伪善拟菱形藻壳面是线形的, 墨西哥海域[37]的亚伪善拟菱形藻是披针形。本文亦支持这一观点。此外, Moschandreou等[26]认为壳环带特征或许可以两者之间的区别依据。伪善拟菱形藻的壳环带高有6—7个孔纹,宽2个孔纹[22], 而亚伪善拟菱形藻的壳环带高2个孔纹, 宽2个孔纹[37]。
本文报道的尖细拟菱形藻和伪柔弱拟菱形藻,此前在我国海域都曾有分布记录[7,8], 但以往研究的物种鉴定手段较为单一, 仅仅依据形态学特征。近年研究认为, 拟菱形藻物种之间的形态区别细微,某些特征也易发生变化, 因此形态特征不能作为物种鉴别的主要依据, 分子生物学数据的引入是有效手段之一[16]。

图版Ⅰ 尖细拟菱形藻PlateⅠ Pseudo-nitzschia cuspidata

图版Ⅱ 伪柔弱拟菱形藻PlateⅡ Pseudo-nitzschia pseudodelicatissima
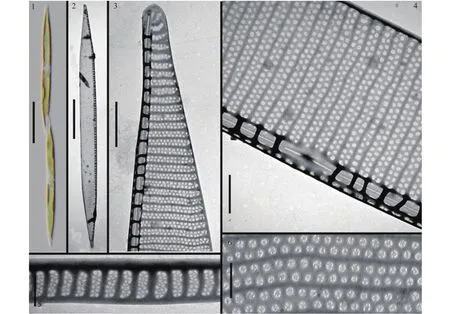
图版Ⅲ 伪善拟菱形藻PlateⅢ Pseudo-nitzschia fraudulenta
