厦门沿海的砂壳纤毛虫(原生动物, 纤毛门, 砂壳目)
2018-09-13廖昱宇刘昭远何如玥焦念志徐大鹏
廖昱宇 刘昭远 何如玥 焦念志 徐大鹏
(1. 厦门大学近海海洋环境科学国家重点实验室, 海洋微型生物与地球圈层研究所, 厦门 361102;2. 厦门大学海洋与地球学院, 厦门 361102)
砂壳纤毛虫是一类具有外壳的纤毛虫原生动物, 个体大小在5—200 μm, 大多在海洋中营浮游生活, 目前共发现约900种[1], 是海洋微型浮游动物群落的重要组成部分。随着20世纪80年代“微食物环”概念的提出, 即异养细菌、微微型浮游植物→原生动物→后生动物, 包括砂壳纤毛虫在内的原生动物的作用日益受到关注[2]。砂壳纤毛虫一方面能摄食微微型(pico-级)和微型(nano-级)浮游生物, 是初级生产力的消费者, 一方面又被中型(meso-级)浮游动物与幼鱼所摄食, 是连接微食物环与经典食物链的重要枢纽, 在海洋生态系统物质循环与能量流动中发挥着重要作用[3—5]。
自1776年Müller报道了第一种砂壳纤毛虫客居筒壳虫(Tintinnus inquilinus)以来, 对于砂壳纤毛虫的研究已历经了2个多世纪[6], 一直以来, 砂壳纤毛虫的分类鉴定及系统发育大多基于外壳形态。近年来, 以基因测序为代表的分子生物学技术越来越多的应用到砂壳纤毛虫研究中[7—10]。最近的研究发现, 砂壳纤毛虫存在外壳多态性(lorica polymorphism)[8]及“隐秘种”(Cryptic species)现象[11,12],基于壳体形态的鉴定及系统发育关系构建受到一定程度的质疑。然而, 由于砂壳纤毛虫(尤其是基于完整形态学记录的)分子数据的匮乏, 多态性及“隐秘种”在砂壳纤毛虫中是否是普遍现象还未为可知。目前, 相关研究中砂壳纤毛虫的鉴定仍主要基于外壳形态, 分子数据的缺乏更凸显了形态学鉴定的重要性。
我国砂壳纤毛虫的分类学研究开始较晚。20世纪30年代至50年代, 倪达书、尹光德等[13—17]先后在厦门沿海、海南岛沿海、胶州湾等海域开展了砂壳纤毛虫调查。自此之后, 一直处于空白状态。直到21世纪初, 才陆续开始对台湾海峡、青岛沿海、广东沿海等海域进行砂壳纤毛虫的分类学研究[18—20]。
厦门沿海砂壳纤毛虫的分类研究始于20世纪30年代倪达书等[13,14]零星的工作。本文在前人研究的基础上, 在厦门沿海进行了多站位(6个)、长时间序列(2015年5月至2016年3月)的样品采集, 以期最大限度获得厦门沿海砂壳纤毛虫信息。同时对获得的27种砂壳纤毛虫的壳体形态进行了详尽的观察、描述与记录及各个种在6个站位以及4个季节的分布情况, 以期为未来的研究提供佐资。
1 材料与方法
分别于2015年5月、6月、7月、9月、10月、11月、2016年1月及3月在厦门沿海6个代表性站位(图1)进行样品采集, 其中S03站位于九龙江口, 受九龙江口冲淡水影响, 盐度变化较大; S05站位于南部黄厝海域; S07站位于东部五缘湾海域、S08站位于大嶝海域, 2个站位靠近外海, 水质较好; S12站位于同安湾、S19站位于西部海域靠近马銮湾, 受陆源生活污水的排放影响较大, 且水流交换缓慢, 污染较为严重。原位环境参数(水温、盐度、pH)以YSI (YSI Professional Plus)测定(表1)。样品以采水器采自表层, 用孔径20 μm的筛绢过滤富集, 平均过水量约10 L。随后用不低于80%(终浓度)的酒精溶液固定, 室温下避光保存。固定后样品在解剖镜(Nikon SMZ1270)下分离并于200—1000倍显微镜(Olympus BX53)下观察, 同时拍照记录(CAMMS软件)。鉴定、系统安排及名词术语参照Kofoid 和Campbell[21]及Corliss[22]。
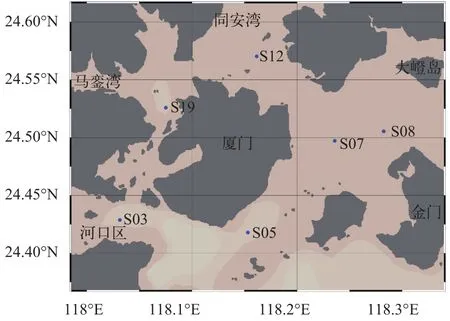
图1 样品采集站位图(黑色圆点代表采样点)Fig. 1 Locations of the sampling stations (black dots show the sampling sites)
2 结果与讨论
Wang和Nie[13]对厦门沿海的砂壳纤毛虫研究中共报道13种, 本研究发现了其中的6种。Nie[14]对厦门沿海砂壳纤毛虫研究中共报道17种, 本研究对其中4种进行了重新描述 (表2)。
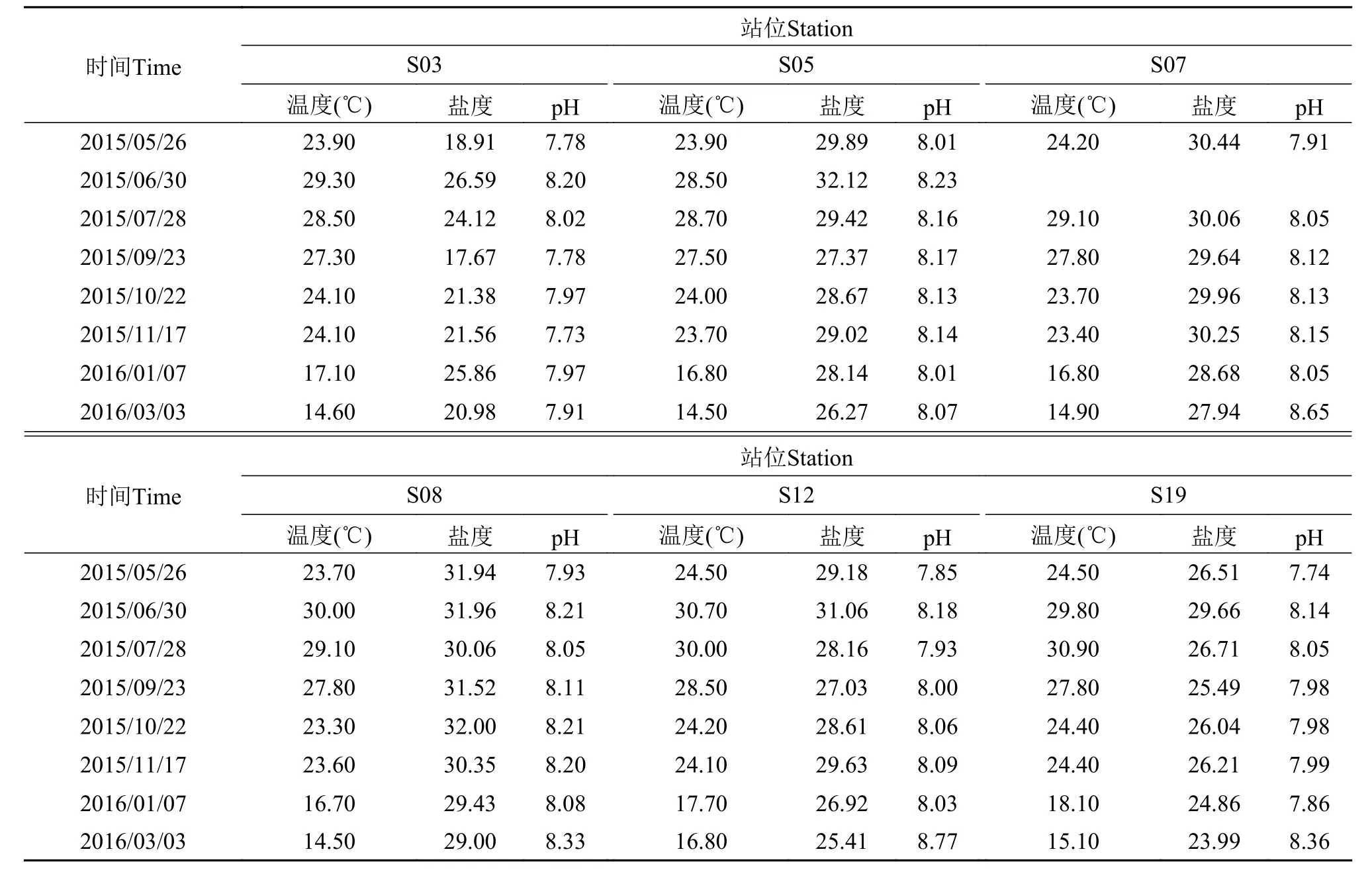
表1 厦门沿海各站位温度、盐度、pHTab. 1 Temperature, salinity, and pH of sampling stations in different months
2.1 长形拟铃虫Tintinnopsis elongate Daday, 1887(图版Ⅰ-1)
发现于夏季, 站位S07。壳体呈子弹形, 表面黏附大量异质颗粒, 壳体前部呈圆柱形, 壳口与壳体前部等宽, 向下渐变为锥形。壳长约83 μm, 口径约40 μm, 壳长约为口径2.1倍, 约于壳体1/2处开始向下缩小呈锥形, 锥形部分长约43 μm。
本种首次由Daday[23]发现于那不勒斯湾。与之相比, 本研究种群个体较小(83 μmvs. 162—198 μm);与Hada[25]于Tropical Pacific西部发现的种群特征吻合。
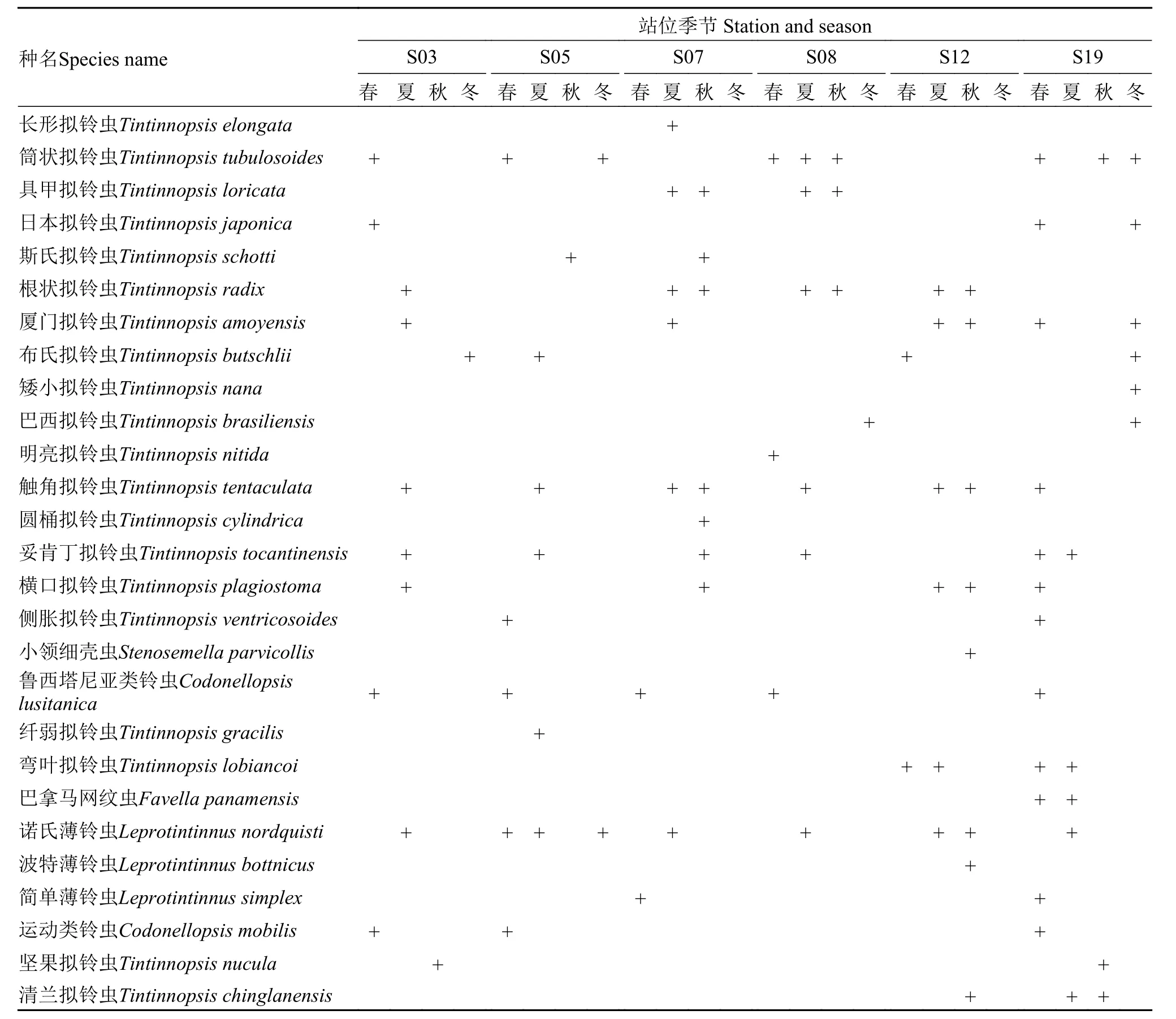
表2 砂壳纤毛虫在6个站位以及4个季节分布情况(+表示在对应站位或季节发现该种, 春季为3—5月, 夏季为6—8月, 秋季为9—11月, 冬季为12—2月)Tab. 2 Distribution of tintinnid ciliates in six stations and four seasons (+ showed the presence of the species. We defined March to May as spring, June to August as summer, September to November as autumn and December to February as winter)
2.2 筒状拟铃虫Tintinnopsis tubulosoides Meunier,1910 (图版Ⅰ-2)
在春季、夏季、秋季、冬季均有发现, 出现在站位S03、S05、S08及S19。壳体呈子弹形, 表面黏附许多异质颗粒。壳体前2/3呈圆柱形, 壳口与壳体前部等宽, 自壳体反口端1/3处向下渐变为锥形。壳长约100—120 μm, 口径约34—43 μm, 锥形部分长约36—40 μm。
本种群与徐大鹏和宋微波[19]在青岛沿海发现的种群特征吻合。
2.3 具甲拟铃虫Tintinnopsis loricata Brandt, 1906(图版Ⅰ-3)
发现于夏季、秋季, 站位S07及S08。壳体呈长钟形, 表面黏附有众多碎屑物质, 分为外展的领部和倒钟形壶部, 口部边缘破碎不规则, 领部和壶部相接处不形成明显的缢缩, 壳室反口端钝圆。壳长约为口径1.6倍, 壳体长约130—135 μm, 口径约84—86 μm, 外展的领部约占壳长的1/4, 领部和壶部交接处直径约为60 μm。
本研究种群与Wang和Nie[13]在厦门湾发现的种群特征吻合。
2.4 日本拟铃虫Tintinnopsis japonica Hada, 1932(图版Ⅰ-4、5)
发现于春季、冬季, 站位S03、S09。壳体呈倒钟形, 表面黏附有众多碎屑物质, 口缘较规则, 领部具有明显环纹, 环纹间大约相隔7 μm, 壳体前2/3呈圆柱状, 宽度几乎相等, 壳体后1/3逐渐缩小呈锥形,末端钝尖。壳体长约为180—187 μm, 口径约115—130 μm。
与尹光德[16]在青岛胶州湾发现的种群相比, 本研究种群个体较大(180—187 μmvs. 114 μm)。
2.5 斯氏拟铃虫Tintinnopsis schotti Brandt, 1906(图版Ⅰ-6)
发现于秋季, 站位S05及S07。壳体呈倒钟形,表面黏附有众多异质块状颗粒, 口缘不规则, 分为领部和壶部, 领部向外伸展, 领部和壶部相接处不形成明显的缢缩, 壶部呈倒锥形, 壳室反口端略有突出。壳体长约114—116 μm, 口径约82—85 μm,壶部长度略小于3/4壳长, 领部和壶部交接处直径约为62 μm。
本种群与T. loricataBrandt, 1906的区别在于,本种群壳体偏小(114—116 μmvs. 130—135 μm),壶部较短, 领部外展程度较低, 且壳体末端略有突出(vs. 钝圆)。
2.6 根状拟铃虫Tintinnopsis radix (Imhof, 1886)Brandt, 1907 (图版Ⅰ-7、8)
发现于夏季、秋季, 出现在站位S03、S07、S08及S12。壳体呈不对称长管状, 壳壁薄, 壳体与壳口近似等宽, 壳口略向外展, 壳体自2/3处向下渐变为锥形, 壳体末端逐渐变细, 末端有一开口。壳长约188—368 μm, 口径约36—47 μm。
与Nie[14]在厦门湾发现的种群相比, 本研究种群个体较小(188—368 μmvs. 330—492 μm); 与尹光德[16]于青岛胶州湾发现的种群特征吻合。
2.7 厦门拟铃虫Tintinnopsis amoyensis Nie, 1934(图版Ⅰ-9、10)
在四个季节均有发现, 出现在站位S03、S07、S12及S19。壳体呈圆筒形, 表面附着较多异质颗粒。口部略微外展, 口缘处黏附大量碎屑, 反口端收缩为锥形, 自壳体末端变细且略微弯曲。壳体长约48—56 μm, 口径约26—30 μm, 锥形部分长约9—14 μm。
该种首次由Nie[14]采自厦门沿海, 本研究种群特征与之吻合。
2.8 布氏拟铃虫Tintinnopsis butschlii Daday, 1887(图版Ⅰ-11)
发现于春季、夏季与冬季, 出现在站位S03、S05、S12及S19。壳体呈花瓶状, 壳体表面黏附碎屑不透明, 领部向外扩展呈喇叭形, 壳体反口端略有膨大且钝圆。壳长约57—91 μm, 口径约52—88 μm, 膨大壶部约占壳长5/8, 领部与壶部交接处约为31 μm。
本研究种群与Nie[14]在厦门湾发现的种群特征一致。
2.9 矮小拟铃虫Tintinnopsis nana Lohmann,1908 (图版Ⅰ-12)
发现于冬季, 出现在站位S19。壳体杯形, 表面黏附有较多异质颗粒, 壳口与壳室几乎等宽, 壳室反口端尖削。壳长约35 μm, 口径约22 μm。
本种与T. minutaWailes, 1925的区别是前者个体较大(35—38 μmvs. 25 μm)且壳室末端尖削(vs.钝圆)。
2.10 巴西拟铃虫Tintinnopsis brasiliensis Kofoid& Campbell, 1929 (图版Ⅰ-13)
发现于冬季, 站位S08及S19。壳体呈倒锥形,壳壁厚, 表面黏附有大量异质颗粒, 约于壳体1/2处略有膨大, 壳体1/2部分收缩呈锥形, 壳体反口端略尖。壳长约70—76 μm, 口径约40—43 μm。
本研究种群与 Hada[25]在赤道太平洋西部发现的种群相比, 壳体更长(76 μmvs. 42—48 μm)。
2.11 明亮拟铃虫Tintinnopsis nitida Brandt, 1896(图版Ⅱ -14)
发现于春季, 出现在站位S08。壳体呈子弹形,表面黏附有众多的异质颗粒, 壳口与壳体几乎等宽,壳室反口端略尖。壳体长约60 μm, 口径约27 μm。
本研究种群与Nie[14]在厦门湾发现的种群特征基本吻合。
2.12 触角拟铃虫Tintinnopsis tentaculata Nie &Cheng, 1947 (图版Ⅱ-15、20)
发现于春季、夏季与秋季, 6个站位均有发现。壳体呈陀螺状, 表面黏附较多异质颗粒, 口区略收缩, 领部伸展形成5根触角状凸起, 反口端膨大,末端稍尖不形成凸起或形成细锥形凸起。壳体长约85—89 μm, 口径约38—45 μm, 领部凸起部分长约18—23 μm, 末端若形成凸起, 长约15—18 μm。
本种首次由Nie和Cheng[15]发现于海南岛沿海,本研究种群与其相比体型较大(85—89 μmvs.52—63 μm), 描述特征基本一致。
2.13 圆桶拟铃虫Tintinnopsis cylindrica Daday,1887 (图版Ⅱ-16、17)
发现于秋季, 出现在站位S07。壳体呈长管状,表面黏附众多的异质颗粒, 口缘较规则, 壳体后1/5逐渐变细为锥形, 末端钝尖。壳体长约为147 μm,口径约36 μm, 锥形部分长约31 μm。
本研究种群与Urrutxurtu[26]在Nervion河口发现的种群特征一致。
2.14 妥肯丁拟铃虫Tintinnopsis tocantinensis Kofoid & Campbell, 1929 (图版Ⅱ-18、19)
发现于春季、夏季与秋季, 出现在站位S03、S05、S07、S08及S19。壳室前1/2呈圆柱形, 壳室后1/2膨大呈球状, 表面黏附众多的异质颗粒, 不同个体间膨大部分直径大小不同, 壳体反口端呈尖削的锥形。壳体长约109—122 μm, 口径约25—28 μm,膨大部分直径大多在38—41 μm, 发现一个样品膨大部分偏小直径仅为29 μm, 尖削的锥形部分长约26—29 μm。
本种群首次由 Kofoid和Campbell[21]报道, 但与原描述相比, 个体较大(109—122 μmvs. 85 μm), 与Nie和Cheng[15]采自海南的种群特征吻合。
2.15 横口拟铃虫Tintinnopsis plagiostoma Daday,1887 (图版Ⅱ-21)
发现于春季、夏季与秋季, 站位S03、S07、S12及S19。壳壁厚, 表面黏附众多的异质颗粒, 分为稍微外展的口部和膨大的壶部, 壳体末端钝圆。壳体长约43—51 μm, 口径约43—50 μm。
本研究种群与丰美萍、张武昌等[27]在南海北部发现的种群特征一致。
2.16 侧胀拟铃虫Tintinnopsis ventricosoides Meunier, 1910中国新记录 (图版Ⅱ-22)
发现于春季, 出现在站位S05及S19。壳体呈倒锥形, 壳壁厚, 表面黏附有大量异质颗粒, 壳体反口端尖削, 口部略收缩。壳体长约72—82 μm, 口径约51—62 μm。
本研究种群与Marshall[28]在Arctic Seas发现的种群特征一致。
2.17 小领细壳虫Stenosemella parvicollis(Marshall) Hada, 1938 (图版Ⅱ-23)
发现于秋季, 出现在站位S12。表面黏附许多异质颗粒, 分为细缩的领部和膨大的壶部, 领部透明无环纹, 壶部膨大, 壳体末端钝圆。壳体长约70 μm,领部长约10 μm, 口径约25 μm, 壶部长约60 μm,宽约50 μm。
比较Hada[25]的描述, 本研究种群壳室较长(70 μmvs. 48—60 μm)。
2.18 鲁西塔尼亚类铃虫Codonellopsis lusitanica Jörgensen, 1924 (图版Ⅱ-24)
发现于春季, 出现在站位S03、S05、S07、S08及S19。分为长圆柱形领部与倒锥形壶部, 领部黏附分散排布的异质颗粒呈半透明状, 大约2/5处开始略微膨大并且向下呈倒圆锥形, 反口端尖削,倒圆锥部壳壁厚黏附大量异质颗粒不透明, 口缘处较平整。壳长约91—100 μm, 口径长为23—27 μm。
本研究种群与Marshall[28]采自南欧与非洲西北沿海的种群特征相一致。
2.19 纤弱拟铃虫Tintinnopsis gracilis Kofoid &Campbell, 1929 (图版Ⅱ-25)
发现于夏季, 出现在站位S05。壳体呈圆柱形,壳壁薄, 表面黏附众多的异质颗粒, 口区无外展略收缩, 口缘较平整, 自壳体3/4处向下渐变为锥形,末端略尖。壳体长约110 μm, 口径约33 μm, 锥形部分长约26 μm。
本研究种群与Wang和Nie[13]采自厦门湾的种群特征相一致。
2.20 弯叶拟铃虫Tintinnopsis lobiancoi Daday,1887 (图版Ⅱ-26)
发现于春季与夏季, 出现在站位S12及S19。壳体呈长圆柱形, 表面黏附有许多异质颗粒, 壳口略宽于壳室, 壳体反口端约八分之一体长处向下渐变为钝圆形。壳长约为82—120 μm, 口径29—42 μm。
本种首次由Daday[23]于那不勒斯湾发现。与Marshall[26]在 Arctic Seas, Baltic, North Sea,Irminger Sea, Newfoundland Banks, Nova Scotia, 欧洲与非洲西北沿岸发现的种群特征一致。
2.21 巴拿马网纹虫Favella panamensis Kofoid &Campbell, 1929 (图版Ⅲ-27)
发现于春季与夏季, 站位S19。壳体呈高脚杯形, 壳口与壳室体部近等宽。壳体长约169—210 μm,其中壳室长125—156 μm, 基柄长约44—51 μm, 口径约59—80 μm, 去除基柄长度后体长约为口径的2倍。
本种群描述与Wang和Nie[13]在厦门湾发现的种群特征基本吻合。
2.22 诺氏薄铃虫Leprotintinnus nordqvisti (Brandt,1906) Kofoid & Campbell, 1929 (图版Ⅲ-28)
发现于春季、夏季与秋季, 六个站位均有发现。壳体呈圆筒状, 表面黏附有分散排布的异质颗粒, 壳壁薄呈半透明, 壳口一端略微外展与壳体几乎等宽, 另一端也有开口, 外展成喇叭状。壳体长约132—177 μm, 与壳体等宽一端开口宽约33—46 μm, 外展一端开口宽约34—79 μm。
本种群描述与Wang和Nie[13]在厦门湾发现的种群特征基本吻合。
2.23 波特薄铃虫Leprotintinnus bottnicus(Nordqvist) Jörgensen, 1912 (图版Ⅲ-29)
发现于秋季, 出现在站位S12。壳体圆柱形, 表面黏附分散排布的异质颗粒, 壳壁薄呈半透明, 壳室两端均开放, 壳口略微外展与壳体几乎等宽, 自壳体约3/5处向下收缩为锥形, 末端开口。壳长约173 μm, 口径约45 μm, 反口端口径约17 μm。
比较Hada[24]的描述, 本研究种群壳室较短(173 μmvs. 205 μm)。本种可以根据壳体形态与L. simplexSchmidt, 1901相区别(壳室反口端开口收缩vs. 壳室两端开口等宽)。
2.24 简单薄铃虫Leprotintinnus simplex Schmidt,1901 (图版Ⅲ-30)
发现于春季, 出现在站位S07及S19。壳体长圆筒状, 一端略弯曲, 一端呈直筒状, 两端均开放, 壳口两端均与壳体几乎等宽, 壳壁薄呈半透明, 表面黏附有分散排布的异质颗粒。壳长约340—415 μm,壳口两端口径约61—67 μm。
本种群与徐大鹏和宋微波[19]在青岛沿海发现的种群相比, 壳长较长(340—415 μmvs. 250—306 μm),口径较大(61—67 μmvs. 50—60 μm), 其他种群特征一致。
2.25 运动类铃虫Codonellopsis mobilis Wang,1936 (图版Ⅲ-31、32)
发现于春季, 出现在站位S03、S05及S19。壳体呈花瓶状, 分为透明的领部和锥形壶部, 壶部黏附大量碎屑物质, 呈不透明状, 口缘不规则, 壳体反口端尖削。壳体长98—115 μm, 口径约50—55 μm,透明领部约占壳长1/9, 长约11—17 μm。
本种首次由Wang[17]发现于渤海, 与尹光德[16]于青岛胶州湾发现的种群特征基本一致, 但本研究种群个体较小, 口径偏小(50—55 μmvs. 62—86 μm)。
2.26 坚果拟铃虫Tintinnopsis nucula (Fol, 1884)Brandt, 1906 (图版Ⅲ-33)
发现于秋季, 出现在站位S03及S19。壳体鸭梨形, 分为细缩的领部和膨大的壶部, 领部透明, 壶部黏附众多的异质颗粒, 壳体反口端钝圆。壳体长约72 μm, 口径约19 μm, 壶部长度约占壳长的8/9。
本研究种群与丰美萍、张武昌等[27]在南海北部发现的种群特征一致。
2.27 清兰拟铃虫Tintinnopsis chinglanensis Nie& Cheng, 1947 (图版Ⅲ-34、35)
发现于夏季与秋季, 出现在站位S12及S19。壳体呈花瓶形, 分为明显的领部和壶部, 领部黏附分散排布的异质颗粒, 呈半透明状, 壶部膨大, 黏附有颗粒物质, 不透明, 领部短于壶部, 口缘较为规则。壳体长约100 μm, 领部长约35 μm, 口径约37 μm, 膨大部分直径约40 μm, 长约65 μm。
本研究种群与尹光德[16]1952年于青岛胶州湾发现的种群特征吻合。
致谢:
感谢厦门大学海洋微型生物与地球圈层研究所王建宁、李冉、张翊哲在样品采集过程中给予的帮助。

图版Ⅰ 砂壳纤毛虫的照片PlateⅠ The microphotographs of tintinnid ciliates

图版Ⅱ 砂壳纤毛虫的照片PlateⅡ The microphotographs of tintinnid ciliates
