不同助跑速度条件下45°急停变向动作的膝和踝关节肌肉激活特征分析
2018-09-07许贻林周苏坡
袁 鹏,许贻林,王 丹,周苏坡,4,陈 伟
不同助跑速度条件下45°急停变向动作的膝和踝关节肌肉激活特征分析
袁 鹏1,2,许贻林1,王 丹3,周苏坡1,4,陈 伟1
1.江苏省体育科学研究所 生物力学实验室, 江苏 南京 210033;2.苏州大学 体育学院, 江苏 苏州 215006; 3.上海体育学院 体育教育训练学院,上海 200438; 4.南京体育学院 运动健康科学系, 江苏 南京 210014
目的:比较不同助跑条件下男、女性运动员在45°急停变向动作中膝、踝关节肌肉激活特征的差异性并进一步分析预激活和后激活阶段膝、踝关节肌肉共激活比之间的相关性。方法:46名青年篮球和排球运动员分别以3、4和5 m/s的助跑速度完成45°急停变向动作。采用Delsys无线表面肌电仪收集股四头肌(股直肌、股外侧肌、股内侧肌)、腘绳肌(半腱肌、股二头肌长头和短头)、胫骨前肌以及腓肠肌外侧头的肌电信号。分析3种助跑速度条件下预激活和后激活阶段膝、踝关节肌肉的标准化均方根振幅以及肌肉共激活比。结果:助跑速度显著增加预激活阶段股四头肌和腓肠肌外侧头的激活水平(<0.05),后激活阶段股四头肌、腘绳肌、胫骨前肌和腓肠肌外侧头的激活水平(<0.05),但显著降低预激活阶段膝、踝关节肌肉共激活比(<0.05)。女性运动员预激活和后激活阶段股四头肌和胫骨前肌的激活水平显著高于男性运动员(<0.05),但预激活和后激活阶段膝关节肌肉共激活比显著低于男性运动员(<0.05)。膝、踝关节预激活和后激活阶段的肌肉共激活比显著相关(<0.05)。结论:随助跑速度的提高,中枢神经系统通过调节预激活和后激活机制来提高肌肉激活水平。预激活和后激活机制之间在45°急停变向动作中存在较高关联性,这提示,通过协调预激活和后激活策略能够调节肌肉和关节的刚度以达到保护下肢关节和组织并提高运动表现的目的。后激活阶段更低的膝关节肌肉共激活比以及股四头肌为主导的激活模式提示,女性运动员在完成45°急停变向动作时具有更高的ACL损伤风险。膝关节肌肉激活模式的性别差异性可能在一定程度上解释ACL损伤风险的性别差异性。
急停变向;预激活;牵张反射;共激活;神经肌肉控制
1 前言
急停变向(side-step cutting)是常见的人体动作模式之一。对于篮球、排球和足球等运动项目来说需要运动员在场上重复完成不同速度和角度的急停变向动作。然而,该动作也常常伴随着运动损伤的发生,如前交叉韧带(ACL)损伤等[26,30]。据Koga[26]和Krosshaug[30]等报道,有超过20%的非接触性ACL断裂发生在急停变向动作过程中。
研究表明,大多数ACL损伤发生在需要身体减速或改变运动方向这一类动作的足触地后瞬间(17~50 ms之内)[30]。当人体着地(landing)或改变姿势时,下肢刚度(lower extremity stiffness)的调节是最先启动的动作神经反射机制[3]。刚度作为一个物理学变量其表示的是物体在受载时抵抗变形的能力[3]。对于着地和急停变向等动作来说重要的调节因素就是腿刚度(leg stiffness),其是由足触地时刻身体环节构型(segmental configuration)和神经肌肉激活(neuromuscular activation)共同作用的结果[22,23]。在神经肌肉方面,人体在足触地前和着地缓冲阶段通过主动肌和拮抗肌的激活来实现对刚度的调节[22,23,28]。Thomas等人认为,肌肉的激活可以成倍地增加关节刚度[46]。而Blackburn等的研究进一步表明,在着地动作中与腘绳肌刚度较低的运动员相比,那些腘绳肌刚度更高的运动员表现出更低的胫骨前向剪切力[8]。从运动损伤预防的角度,更大的关节和肌肉刚度可能代表着更好的关节稳定性,更低的损伤风险;而从运动表现的角度,较低的刚度或较大的屈服(compliance)可能更有利于力学负荷的吸收和弹性势能的储备从而增加运动表现[36]。因此,人体在神经系统的支配下需要选择合理的肌肉激活策略,以便在避免运动损伤的同时实现更高的运动表现。
肌肉共激活(muscle co-activation)指关节周围不同肌肉的同步活动[25]。由于肌肉共激活与关节的稳定性紧密相关[2],因此可能在运动损伤预防中发挥着重要的作用。如腘绳肌和股四头肌的共激活不仅可以帮助膝关节对抗前向剪切力,还可以对抗膝关节的外展以及下肢动态外翻(dynamic lower extremity valgus)[7]。目前已有关于下肢肌肉共激活在步态(gait)[42]、跳深(drop jump, DJ)[31,39]、快速起跳[2]和着地[1,11,35]等动作的研究报道,但是,针对急停变向动作下肢肌肉共激活模式的研究并不多见[7,20]。
鉴于在急停变向和跳深等动作中,助跑速度在很大程度上决定了实际的碰撞负荷[29,47],助跑速度的变化可能会对神经肌肉的激活策略产生影响。Ruan等报道,着地前的助跑速度显著增加跳深动作中预激活和后激活阶段股直肌、股外侧肌、股二头肌等的激活水平[39]。然而,尚未发现针对助跑速度对急停变向动作中神经肌肉激活特征影响的研究。此外,已有研究表明,在急停变向动作中女性研究对象表现出了与男性研究对象不同的神经肌肉激活策略。Hanson等发现,与男性运动员相比,女性运动员在急停变向动作足触地前阶段以及缓冲支撑阶段股四头肌与腘绳肌的共激活比显著增加,女性运动员在完成急停变向动作时表现出明显的股四头肌主导的肌肉激活模式(quadriceps dominant muscle activation pattern)[20]。由此可以推断,性别可能会对急停变向动作中主动肌和拮抗肌的共激活策略产生影响。但是,针对不同助跑速度条件下男、女性运动员在急停变向动作中的神经肌肉控制机制尚不明确。
因此,本研究的目的包括:1)分析不同助跑速度条件下男、女性运动员在45°急停变向动作中膝、踝关节肌肉激活特征的差异性;2)分析预激活和后激活阶段膝、踝关节肌肉共激活比之间的关联性,进而对男、女性运动员在45°急停变向动作中的神经肌肉激活机制进行研究。本研究假设:1)助跑速度和性别显著影响膝、踝关节肌肉激活特征以及膝、踝关节肌肉共激活比;2)预激活和后激活阶段膝、踝关节肌肉共激活比之间存在显著性相关关系。
2 研究对象与方法
2.1 研究对象
选取46名青年篮球和排球运动员为研究对象。所有研究对象在实验前均无任何下肢手术史,在实验前三个月内无任何影响完成正常急停变向动作的损伤。在实验正式开始之前向所有的研究对象讲解研究目的、测试动作要求和实验流程等。研究对象基本情况见表1。
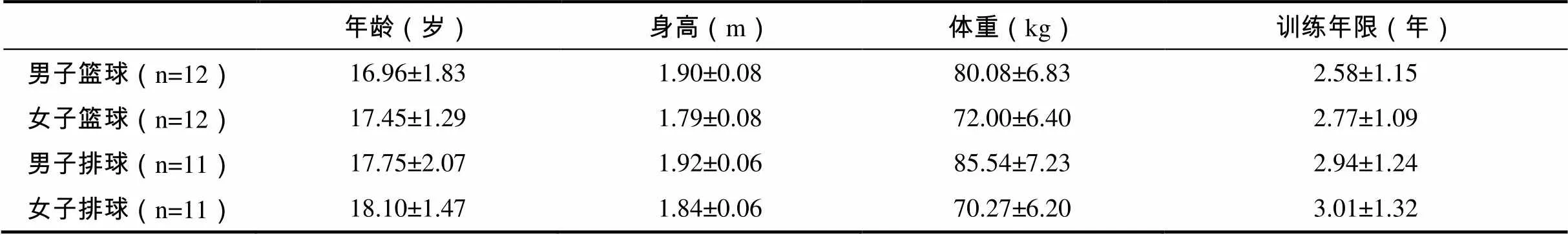
表1 研究对象基本情况一览表
2.2 研究方法
2.2.1 实验设计
研究对象分别以3 m/s(慢速)、4 m/s(中速)和5 m/s(快速)的助跑速度完成45º急停变向动作。本研究中使用的急停变向动作要求研究对象优势侧腿的足接触测力台之后开始减速缓冲,然后改变身体重心的方向并开始在新的动作方向上通过优势侧腿蹬地加速。测试现场示意图如图1所示,45º急停变向动作如图2所示。在测试开始前运动员完成10 min的准备活动,包括5 min的慢跑和5 min的动态拉伸。每个助跑速度条件下采集3次有效动作。有效动作被定义为助跑速度在规定的范围内,动作全程无肌电信号丢失且优势侧足完全落在测力台上。有效的助跑速度区间分别为2.7~3.3 m/s(慢速)、3.7~4.3 m/s(中速)和4.7~5.3 m/s(快速)。运动员每次完成的助跑速度完全随机,动作之间运动员有30 s的休息时间。
2.2.2 数据采集
两台Smartspeed测速仪(Fusion Sport, Australia)分别距测力台5 m和0 m用于记录运动员的助跑速度。Smartspeed测速仪的高度与运动员的髋关节同高[48]。运动员从距离测力台5 m之外的位置开始助跑。在每次数据采集之前运动员有机会熟悉测试动作并确定合适的起跑位置。一台三维测力台(9287C, Kistler Instruments, Winterthur, Switzerland)安装在测试区域的中央位置以采集45°急停变向动作过程中地面反作用力参数用于确定足触地时刻,采样频率为1 000 Hz。采用Delsys Trigno Lab 16通道无线表面肌电仪(Delsys, Boston, MA, USA)采集运动员在完成45°急停变向动作过程中优势侧股直肌(rectus femoris, RF)、股内侧肌(vastus medialis, VM)、股外侧肌(vastus lateralis,VL)、股二头肌长头(biceps femori: long head)、股二头肌短头(biceps femori: short head)、半腱肌(semitendinosus)、胫骨前肌(tibialis anterior, TA)和腓肠肌外侧头(lateral gastrocnemius, LG)的表面肌电信号,采样频率为2 000 Hz。优势侧下肢定义为运动员习惯性完成单腿起跳侧下肢[49]。在安置电极前先进行备皮工作,具体步骤如下:先使用剃须刀将肌肉表面的体毛剃除;再利用砂纸打磨皮肤表面直至皮肤呈微红色;然后用75%浓度的医用酒精处理打磨好的皮肤,最后待酒精完全挥发后利用双面胶将肌电电极沿着肌纤维的走向贴在相应肌肉的肌腹位置。电极放置的具体位置参考Perotto等人的方法[38]。电极放置完成后利用弹性绑带将其固定以避免脱落。利用Qualisys Track Manager 2.15三维红外动作捕捉系统(Oqus, Qualisys, Gothenburg, Switzerland)对表面肌电信号和三维测力台信号进行同步,采样频率为200 Hz。

图1 45°急停变向动作测试现场示意图
Figure1. Experimental Setup for 45°Side-step Cutting
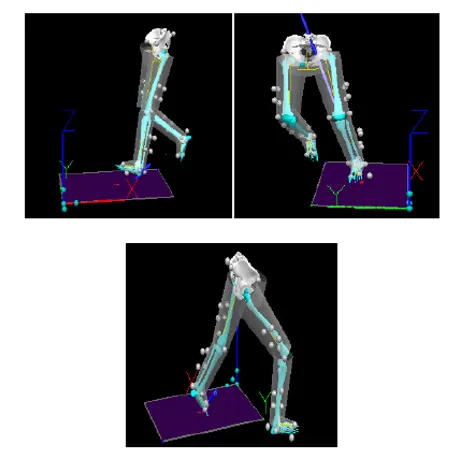
图2 45°急停变向动作示意
Figure2. 45°Side-step Cutting
注:A足触地时刻,B膝关节最大屈角时刻,C足离地时刻
2.2.3 数据处理
利用Delsys File Utility将由Delsys EMGworks 4.3软件所采集的原始肌电信号转换为C3D格式。然后,采用Visual 3D v6.0(C-Motion, Germantown, USA)对原始肌电数据进行处理。采用零滞后四阶递归巴特沃斯滤波器对原始肌电信号进行带通过滤,截断频率为10~500 Hz以除去人为干扰。对过滤后的肌电信号进行全波整流(取绝对值),再进行低通过滤,截断频率为6 Hz[7,10]。
本研究评价的参数包括预激活阶段(pre-activation)和后激活阶段(post-activation)标准化股四头肌(quadriceps, Q)、腘绳肌(hamstrings, H)、胫骨前肌(TA)和腓肠肌外侧头(LG)的均方根振幅,膝关节肌肉共激活比(knee co-activation ratio)和踝关节肌肉共激活比(ankle co-activation ratio)。肌肉共激活比的计算步骤如下:
1. 提取被测肌肉在慢速助跑45º急停变向动作中过滤后的肌电信号峰值。
2. 计算被测肌肉三次有效动作过滤后的肌电信号峰值的平均值。
3. 以提取过滤后的肌电信号峰值平均值为标准,计算其它助跑速度条件下相应肌电信号的相对值[1,9,37]。Branch等认为,利用功能性动作(functional movement)来标准化肌电信号与采用MVIC方法相比可以降低运动员之间的变异性[9]。目前,该方法广泛用于变向[7,37]和着地[1]等动作的肌电信号处理。


2.2.4 统计学分析
采用3×2(助跑速度×性别)重复测量方差分析(repeated-measures ANOVA)观察助跑速度和性别对股四头肌、腘绳肌、胫骨前肌和腓肠肌外侧头在预激活和后激活阶段肌电信号均方根振幅以及膝、踝关节肌肉共激活比的主效应。采用Tukey’s post-hoc test对显著性主效应进行事后检验。采用皮尔逊积距相关(pearson’s product moment correlation)观察预激活和后激活阶段膝、踝关节肌肉共激活比之间的相关性。利用相关性系数和显著性水平衡量相关性的强弱。=0.10表示低相关性,=0.30表示中等相关性,=0.50表示高相关性[12]。统计学分析的软件为SPSS 22.0,显著性水平定义为ɑ=0.05。
3 研究结果
3.1 助跑速度
男性运动员的实际助跑速度分别是3.20±0.17、3.95±0.23和4.72±0.21 m/s,女性运动员的实际助跑速度分别是3.16±0.20、4.02±0.20和4.92±0.37m/s。重复测量方差分析结果表明3个助跑速度之间均在显著性差异(2,88=6.374,<0.001),而性别对运动员的助跑速度没有显著性影响(1,44=3.292,=0.076)。事后检验结果表明,无论是男性运动员还是女性运动员,3个助跑速度之间均存在显著性差异(<0.05)。
3.2 膝、踝关节肌肉激活特征
3.2.1 助跑速度对预激活和后激活阶段膝、踝关节肌肉激活特征的影响
在预激活阶段,助跑速度对股四头肌(2,88=17.957,<0.001)、腘绳肌(2,88=3.452,=0.046)和腓肠肌外侧头(2,88=5.611,=0.011)的激活水平以及膝关节(2,88=11.018,=0.001)和踝关节肌肉共激活比(2,88=4.365,=0.016)的主效应有显著性影响。就男性运动员而言,事后检验结果表明,快速助跑条件下股四头肌激活水平与慢速(<0.001)和中速(=0.006)相比存在明显的差异;快速助跑条件下腓肠肌外侧头激活水平(=0.014)以及踝关节肌肉共激活比(=0.013)与慢速相比存在明显的差异;慢速助跑条件下膝关节肌肉共激活比与中速(=0.010)和快速(=0.001)相比存在显著性差异。就女性运动员而言,事后检验结果表明,快速助跑条件下股四头肌激活水平(=0.002)、膝关节肌肉共激活比(=0.008)与慢速相比存在明显的差异(表2和图3)。
在后激活阶段,助跑速度显著影响股四头肌(2,88=14.340,<0.001)、腘绳肌(2,88=6.125,=0.008)、胫骨前肌(2,88=5.921,=0.008)和腓肠肌外侧头(2,88=12.138,<0.001)的主效应。就男性运动员而言,事后检验结果表明,快速助跑条件下股四头肌激活水平与慢速(<0.001)和中速(=0.014)相比存在明显的差异;快速助跑条件下腘绳肌(=0.003)和腓肠肌外侧头(=0.004)激活水平与慢速相比存在明显的差异。就女性运动员而言,事后检验结果表明,快速助跑条件下股四头肌激活水平与慢速(<0.001)和中速(=0.010)相比存在明显的差异;快速助跑条件下腘绳肌(=0.012)、胫骨前肌(=0.016)和腓肠肌外侧头(=0.010)激活水平与慢速相比存在明显的差异(表3和图4)。

表2 预激活阶段膝、踝关节肌肉的标准化均方根振幅和肌肉共激活比一览表
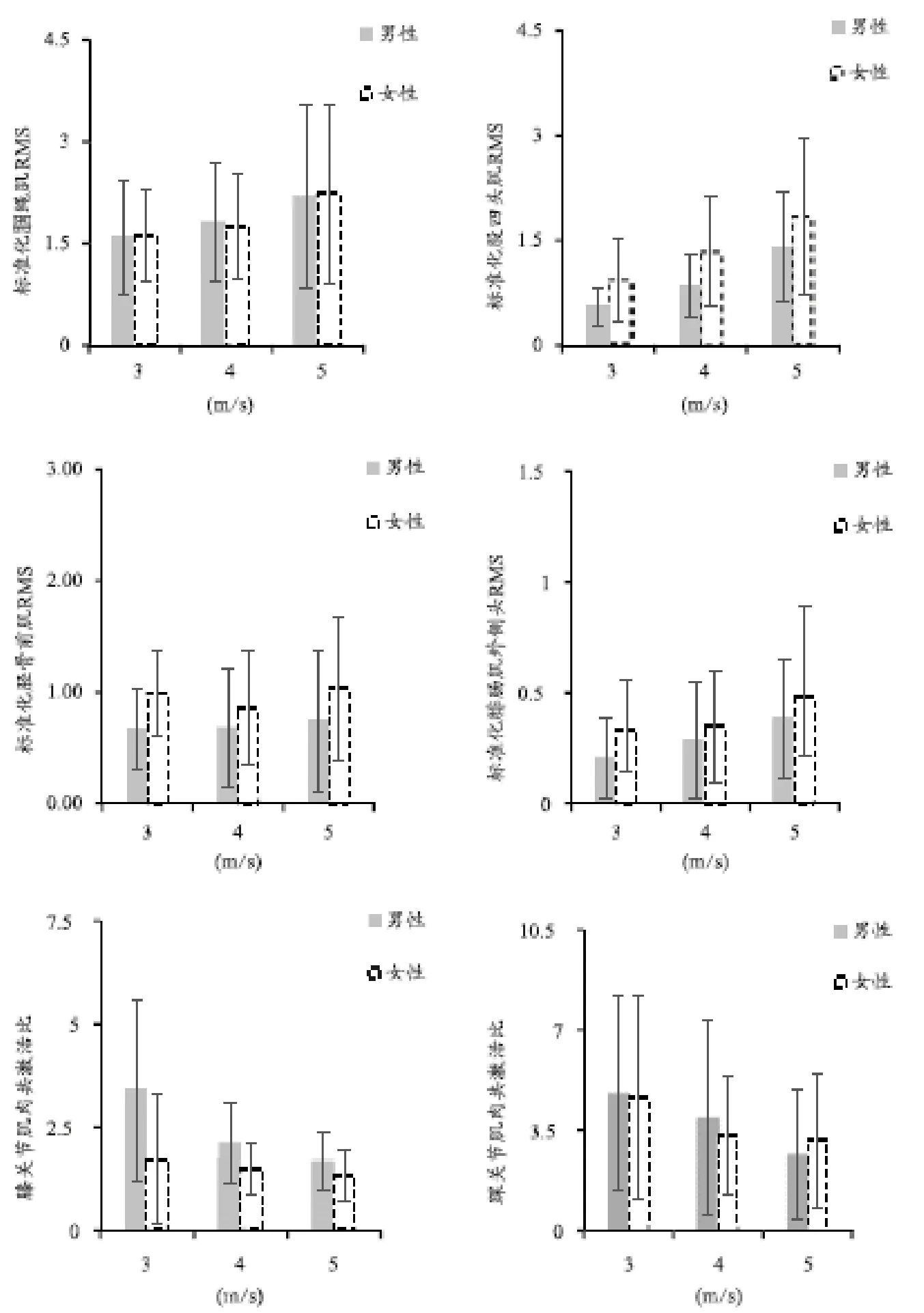
图3 助跑速度和性别对预激活阶段膝、踝关节肌肉的标准化均方根振幅和肌肉共激活比的影响
Figure3. Effects of Approach Run Speed and Gender on Normalized Root Mean Square Amplitudes and the Co-activation Ratio of Knee and Ankle Muscles in the Pre-activation Phase
注:*表示助跑速度之间存在显著性差异(<0.05),下同。
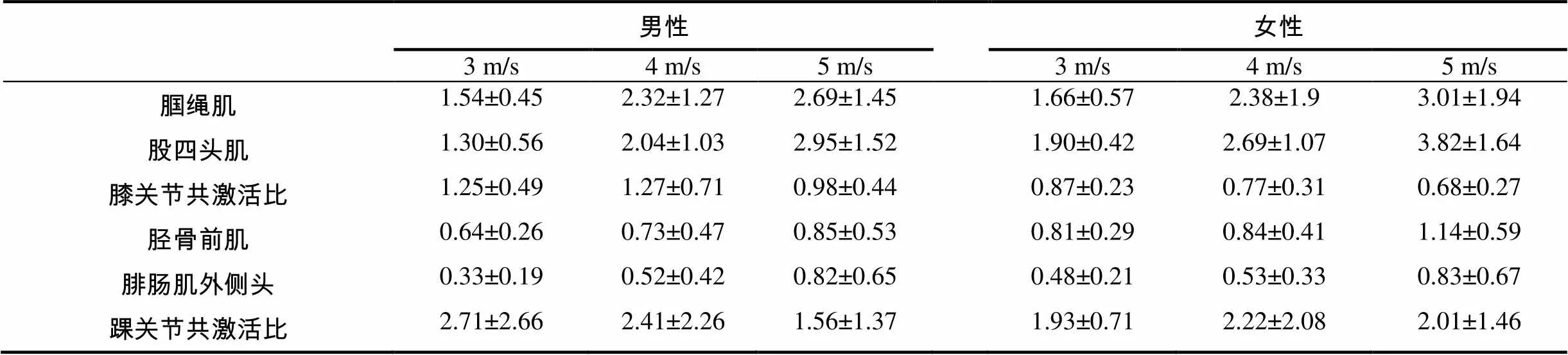
表3 后激活阶段膝、踝关节肌肉的标准化均方根振幅和肌肉共激活比一览表
3.2.2 性别对预激活和后激活阶段膝、踝关节肌肉激活特征的影响
在预激活阶段,性别对股四头肌(1.44=6.899,=0.013)和胫骨前肌(1.44=4.299,=0.045)激活水平以及膝关节肌肉共激活比(1.44=19.788,<0.001)的主效应有显著性影响(表2和图3)。在后激活阶段,性别显著影响股四头肌(1.44=8.379,=0.007)和胫骨前肌(1.44= 4.090,=0.045)激活水平以及膝关节肌肉共激活比(1.44=9.658,=0.004)的主效应(表3和图4)。助跑速度和性别对预激活和后激活阶段膝、踝关节肌肉的标准化均方根振幅和肌肉共激活比均无交互影响。
Figure4. Effects of Approach Run Speed and Gender on Normalized Root Mean Square Amplitudes and the Co-activation Ratio of Knee and Ankle Muscles in the Post-activation Phase
3.2.3 预激活和后激活阶段膝、踝关节肌肉共激活比的相关性皮尔逊积距相关分析结果表明,预激活和后激活阶段的整体(合并助跑速度和性别)膝关节肌肉共激活比(=0.790,<0.001)、踝关节肌肉共激活比(=0.652,<0.001)显著相关。就具体的助跑速度而言,慢速、中速和快速助跑条件下预激活阶段和后激活阶段的膝关节肌肉共激活比(0.692≤≤0.823,<0.001)、踝关节肌肉共激活比(0.604≤≤0.715,<0.001)均呈显著性正相关关系。无论是男性运动员还是女性运动员,其预激活阶段和后激活阶段的膝关节肌肉共激活比(=0.823和502,<0.001)、踝关节肌肉共激活比(=0.795和0.440,<0.001)均呈显著性正相关关系(表4)。

表4 预激活和后激活阶段膝关节和踝关节肌肉共激活比的相关系数一览表
注:显著性水平为<0.05。
4 讨论与分析
4.1 助跑速度对预激活和后激活阶段膝、踝关节肌肉激活特征的影响分析
急停变向和跳深等快速拉伸-缩短循环(stretch-shortening cycle,SSC)动作涉及前馈(如预激活)和牵张反射(如后激活)等神经控制机制[44]。在前馈机制方面,Dietz[16]和Gollhofer[18]等将预激活解释为预编程的神经元激活(pre-programmed neuronal activation)。预激活的时序和幅度受中枢神经系统的控制,其主要功能在于让肌肉在较短的时间内产生最大的力[40]。在类似于急停变向和跳深等动作中,肌肉需要100 ms左右的时间来产生最大的力[39];然而,大多数ACL损伤发生在足触地后瞬间(足触地后的17~50 ms之内)[30]。这表明在快速运动中如果肌肉没能被充分预激活,那么其对关节和软组织的功能将被抑制。当肌肉在足触地前被充分激活(预激活),肌肉的刺激动力学(stimulation dynamics)和兴奋动力学(excitation dynamics)开始工作,以至于在足触地后肌肉力量得以迅速提高[41]。因此,预激活可能通过调节肌肉力潜能[41]和关节刚度[27]来实现对关节或组织的保护。足着地后短潜伏期反应(short latency response, SLR)是最早的牵张反射反应期[45]。Cronin等的研究发现,SLR水平与碰撞加速度峰值显著相关,而与跖屈牵拉的幅度之间没有相关性[14]。基于这一发现,作者认为,扰动开始后振动刺激(vibratory stimuli)传递到下肢,增加了对肌梭Ia型传入神经纤维刺激从而提高肌肉的后激活水平[14]。Komi[27]和Cronin[13]等发现,后激活阶段肌肉激活水平的提高可能会增加着地动作早期离心阶段的肌肉刚度。基于此,缓冲支撑期间牵张反射可能通过增加下肢肌肉刚度来影响变向动作的表现。
本研究结果部分支持第1个研究假设。助跑速度显著影响膝、踝关节肌肉(股四头肌、腘绳肌和腓肠肌外侧头等)预激活和后激活阶段的标准化均方根振幅,具体表现为:预激活阶段男性运动员快速助跑条件下股四头肌激活水平显著高于慢速和中速助跑,快速助跑条件下腓肠肌外侧头激活水平显著高于慢速助跑;女性运动员快速助跑条件下股四头肌激活水平显著高于慢速助跑。对于后激活阶段,男性运动员快速助跑条件下股四头肌激活水平显著高于慢速和中速助跑,快速助跑条件下腘绳肌和腓肠肌外侧头激活水平显著高于慢速助跑;女性运动员快速助跑条件下胫骨前肌激活水平显著高于慢速助跑。预激活阶段肌肉活动的主要作用在于准备肌肉-肌腱复合体(muscle-tendon complex)以应对足着地后快速、强有力的拉伸动作[40]。当考虑整个肌肉-肌腱复合体在应对强有力拉伸的反应时,由于处于激活状态肌肉的肌腱刚度相对稳定,而肌肉刚度是唯一受神经调节的成分[50],因此,中枢神经系统通过增加肌肉的预激活水平来提高肌肉和关节的刚度以吸收由于更高的助跑速度所产生的更大地面反作用力和身体环节动能。如前所述,助跑速度在很大程度上决定了实际的碰撞负荷[29,47],助跑速度增加时中枢神经系统通过提高相关肌肉的预激活水平来预知更大的地面反作用力[40]。Tam等人的研究也发现跑动速度显著影响预激活阶段股二头肌和胫骨前肌等的激活水平,而且预激活阶段膝关节共激活比与膝关节的刚度存在显著性正相关关系[43]。这一研究发现证实了预激活阶段肌肉激活水平与关节刚度之间的关联性。目前,尚未发现有关于助跑速度对45°急停变向动作中下肢肌肉预激活水平影响的研究报道。而本研究的结果与Christoforidou等针对跳深动作的研究结果基本一致[11]。该研究表明,在跳深动作中随着着地高度的增加预激活和牵张反射的水平也随之增加[11]。此外,贾谊等通过对不同起跳方式下膝关节肌肉共收缩指数进行研究后认为,动作速度可能是影响中枢神经系统对主动肌与拮抗肌控制策略的主要因素[2]。这表明神经系统通过调节肌肉的预激活水平这一策略来为更大的碰撞负荷做准备。此外,更高的预激活和后激活水平可以通过准备拮抗肌(腘绳肌和胫骨前肌)以提高肌肉的刚度这种方式来更好地对抗由于更快的助跑速度所产生的更大碰撞负荷[27]。关于助跑速度对肌肉后激活水平的影响机制,助跑过程中所产生的肌肉组织振动可能具有一定的影响。振动假设(vibration hypothesis)表明,更快的助跑速度可能对肌梭感受器产生更大的振动刺激,从而引起更高的肌肉后激活水平[14]。本研究的结果证实,随着助跑速度的增加,后激活阶段膝、踝关节肌肉的激活水平显著增加。但是,这一激活水平的增加是否与不同助跑速度条件下所产生的振动刺激有关仍需进一步的验证。
除了前馈和牵张反射机制之外,主动肌和拮抗肌的共激活也可能发挥着重要的作用。Baratta等的研究表明,主动肌和拮抗肌的共激活可以提高关节的稳定性并促进关节周围力的合理分布[6]。本研究的结果表明,在预激活阶段,慢速和快速助跑条件下男、女性运动员膝关节肌肉共激活比分别为3.39和1.72以及1.67和1.34。尽管助跑速度显著降低预激活阶段膝关节肌肉共激活比,但是其均不小于1。膝关节肌肉共激活比大于1提示在预激活阶段膝关节以腘绳肌为主导的激活模式。这对于膝关节的损伤预防具有重要的意义。从力学的角度,当施加的负荷超过软组织的强度时便可能出现损伤。就ACL而言,其负荷取决于作用于膝关节的外部负荷(external load)、外力作用时的膝关节表面的运动学(kinematics)和几何位置(geometry position)以及膝关节周围肌肉的激活特征(muscle activation)[7]。在足触地前,中枢神经系统通过激活腘绳肌的方式来对抗足触地后胫骨相对于股骨的前向移动从而起到保护ACL的作用[7]。因此,从神经肌肉激活的角度,在预激活阶段由于助跑速度的提高而引起的膝关节肌肉共激活比的增加可能不会对ACL的损伤产生显著影响。
就踝关节而言,助跑速度显著增加预激活腓肠肌外侧头的激活水平,而踝关节肌肉共激活比显著下降。踝关节肌肉共激活比下降这一结果与傅维杰等的研究结果基本一致。傅维杰等的研究发现,随着着地下落高度的增加,预激活阶段踝关节肌肉共激活比表现出下降的趋势[1]。快速助跑条件下较低的踝关节肌肉共激活比提示胫骨前肌的激活水平与腓肠肌外侧头的激活水平相比明显不足,这可能在一定程度上影响踝关节的稳定性。Lin等发现,胫骨前肌的激活水平与踝关节动态稳定性呈正相关关系[32]。因此,在快速助跑条件下,较低的胫骨前肌激活水平可能不利于对踝关节的保护。这对于功能性踝关节不稳的患者来说尤其重要。
4.2 性别对预激活和后激活阶段膝、踝关节肌肉激活特征的影响分析
本研究结果部分支持性别对膝、踝关节肌肉激活特征存在显著性影响这一假设。本研究发现,女性运动员前、后激活阶段膝关节肌肉共激活比显著低于男性运动员。3种助跑速度条件下前激活阶段男、女性运动员的膝关节肌肉共激活比分别是3.39和1.72,2.13和1.50以及1.67和1.34,3种助跑速度条件下后激活阶段男、女性运动员的膝关节肌肉共激活比分别是1.72和0.89,1.27和0.77以及0.98和0.68。膝关节肌肉共激活比小于1提示以股四头肌主导的激活模式[4]。这表明预激活阶段尽管膝关节肌肉共激活比存在性别差异性,但是膝关节仍以腘绳肌激活为主导。然而,除了男性运动员在慢速和中速助跑条件下后激活阶段膝关节以腘绳肌激活为主导外,其他条件下,男、女运动员后激活阶段膝关节均以股四头肌激活为主。此外,这些数据还提示,随助跑速度的增加,男、女运动员在45°急停变向动作的后激活阶段更多地依靠股四头肌主导的激活模式。
当腘绳肌激活不充分或激活时序不适宜时其对膝关节的保护功能将被抑制,膝关节损伤风险也随之上升[21]。一项前瞻性研究报道,与未损伤的运动员相比,经历ACL断裂的运动员在完成变向动作时半腱肌的前激活水平显著降低;相反,股外侧肌的前激活水平显著提高[51]。这一发现进一步证实了股四头肌和腘绳肌的共激活在预防ACL损伤中的重要作用。针对膝关节主动肌和拮抗肌的共激活现象,De Luca等提出了股四头肌和腘绳肌之间的“共驱动”(common drive)机制[15]。作者认为主动肌和拮抗肌的激活水平受中枢共激活机制的控制。当主动肌和拮抗肌参与某一特定动作时,共驱动通过让它们作为一个整体来控制了肌肉的运动单位[15]。膝关节的共驱动通过合理地量化股四头肌和腘绳肌激活比来最小化膝关节胫骨的前向剪切力,从而降低作用在ACL上的力学负荷。但是,本研究和Hanson等[20]的研究结果表明“共驱动”机制似乎也存在性别差异性。女性运动员表现出与腘绳肌不匹配的股四头肌激活水平。这对于ACL损伤预防来说具有重要的意义。在膝关节屈角小于45°时,股四头肌产生的力通过牵拉髌韧带的方式增加ACL的负荷[34]。有研究报道,在45°急停变向动作足触地时刻膝关节的屈角在15°~30°之间[20]。由此可见,女性运动员膝关节肌肉共激活比的下降可能是其ACL损伤风险提高的潜在危险因素之一。
4.3 预激活和后激活阶段膝、踝关节肌肉共激活比的相关性分析
本研究的结果支持第2个研究假设。无论是从整体还是从助跑速度和性别的角度,膝、踝关节预激活和后激活阶段肌肉共激活比均存在中等到高度的相关性(0.440≤≤0.823)。Avela等认为,肌肉的预激活水平可能影响肌肉的后激活水平[5]。Lesinski等的研究发现,在着地起跳动作中,前馈和牵张反射机制之间存在关联性[31]。本研究进一步证实,45°急停变向动作的前馈和牵张反射机制之间存在关联性。Linnamo等认为,只有在牵张之前产生最大的预激活时更高的牵张速度才能发挥更大的肌肉力潜能[33]。从功能性的视角,预激活和后激活之前的关联性主要用于调节肌肉刚度。Gottlieb等认为,预激活通过提高α-γ共激活来增加肌梭的敏感性,从而促进牵张反射[19]。而牵张反射作为一种补偿机制可以增加拉长阶段的肌肉刚度。因此,人体在中枢神经系统的支配下通过协调预激活和后激活策略来促进肌肉刚度的调节从而实现对下肢关节和组织的保护并提高运动表现。
5 结论
助跑速度显著影响膝、踝关节周围肌肉的激活特征。随助跑速度的增加,中枢神经系统通过调节预激活和后激活水平来提高肌肉激活水平。在45°急停变向动作中预激活和后激活之间存在较高关联性,这提示,中枢神经系统通过协调预激活和后激活策略调节肌肉和关节的刚度以达到保护下肢关节和组织并提高运动表现的目的。后激活阶段女性运动员的膝关节肌肉共激活比显著低于男性运动员,且均以股四头肌为主导的激活模式。膝关节肌肉激活模式的显著性别差异提示,女性运动员在完成45°急停变向动作时较男性运动员相比具有更高的ACL损伤风险。
[1] 傅维杰,刘宇,黄灵燕,等.不同着地方式下鞋缓冲特性对下肢肌肉活化及共激活的影响[J].中国运动医学杂志,2014,33(9):860-868.
[2] 贾谊,薛瑞婷,魏亮.人体快速起跳动作的下肢表面肌电信号特征研究 [J].中国体育科技,2017,53(2):64-70.
[3] 刘宇,魏勇.运动科学领域的下肢刚度研究 [J].上海体育学院学报,2008,32(5):31-35.
[4] AAGAARD P, SIMONSEN E B, ANDERSEN J L,. Antagonist muscle coactivation during isokinetic knee extension [J]. Scan J Med Sci Sports, 2000, 10(2):58-67.
[5] AVELA J, SANTOS P M, KOMI P V. Effects of differently induced stretch loads on neuromuscular control in drop jump exercise [J]. Eur J Appl Physiol, 1996, 72(5-6):553-562.
[6] BARATTA R, SOLOMONOW M, ZHOU B H,. Muscular coactivation. The role of the antagonist musculature in maintaining knee stability [J]. Am J Sports Med, 1988, 16(2):113-122.
[7] BESIER T F, LLOYD D G, ACKLAND T R. Muscle activation strategies at the knee during running and cutting maneuvers [J]. Med Sci Sports Exerc, 2003, 35(1):119-127.
[8] BLACKBURN J T, PADUA D A, WEINHOLD P S,. Comparison of triceps surae structural stiffness and material modulus across sex [J]. Clin Biomech, 2006, 21(2):159-167.
[9] BRANCH T P, HUNTER R, DONATH M. Dynamic EMG analysis of anterior cruciate deficient legs with and without bracing during cutting [J]. Am J Sports Med, 1989, 17(1):35-41.
[10] BROWN T N, MCLEAN S G, PALMIERISMITH R M. Associations between lower limb muscle activation strategies and resultant multi-planar knee kinetics during single leg landings [J]. J Sci Med Sport, 2014, 17(4):408-413.
[11] CHRISTOFORIDOU A, PATIKAS D A, BASSA E,. Landing from different heights: Biomechanical and neuromuscular strategies in trained gymnasts and untrained prepubescent girls [J].J Electromyogr Kinesiol, 2017, 32(2):1-8.
[12] COHEN J. A power primer [J]. Psychol Bull, 1992, 112(1):155-159.
[13] CRONIN N J, CARTY C P, BARRETT R S. Triceps surae short latency stretch reflexes contribute to ankle stiffness regulation during human running [J]. PloS One, 2011, 6(8):e23917.
[14] CRONIN N J, RANTALAINEN T, AVELA J. Triceps surae fascicle stretch is poorly correlated with short latency stretch reflex size [J]. Muscle Nerve, 2015, 52(2):245-251.
[15] DE LUCA C J, MAMBRITO B. Voluntary control of motor units in human antagonist muscles: coactivation and reciprocal activation [J]. J Neurophysiol, 1987, 58(3):525-542.
[16] DIETZ V, SCHMIDTBLEICHER D, NOTH J. Neuronal mechanisms of human locomotion [J]. J Neurophysiol, 1979, 42(5):1212-1222.
[17] FU W, FANG Y, GU Y,. Shoe cushioning reduces impact and muscle activation during landings from unexpected, but not self-initiated, drops [J]. J Sci Med Sport, 2017, 20(10):915-920.
[18] GOLLHOFER A, KYROLAINEN H. Neuromuscular control of the human leg extensor muscles in jump exercises under various stretch-load conditions [J]. Int J Sports Med, 1991, 12(1):34-40.
[19] GOTTLIEB G L, AGARWAL G C, JAEGER R J. Response to sudden torques about ankle in man. IV. A functional role of alpha-gamma linkage [J]. J Neurophysiol, 1981, 46(1):179-190.
[20] HANSON A M, PADUA D A, TROY B J,. Muscle activation during side-step cutting maneuvers in male and female soccer athletes [J]. J Athl Train, 2008, 43(2):133-143.
[21] HEWETT T E, MYER G D, FORD K R. Anterior cruciate ligament injuries in female athletes: Part 1, mechanisms and risk factors [J]. Am J Sports Med, 2006, 34(2):299-311.
[22] HOFFREN M, ISHIKAWA M, KOMI P V. Age-related neuromuscular function during drop jumps [J]. J Appl Physiol, 2007, 103(4):1276-1283.
[23] HORIRA T, KOMI P V, NICOL C,. Interaction between pre-landing activities and stiffness regulation of the knee joint musculoskeletal system in the drop jump: implications to performance [J]. Eur J Appl Physiol, 2002, 88(1-2):76-84.
[24] JONES P A, HERRINGTON L C, MUNRO A G,. Is there a relationship between landing, cutting, and pivoting tasks in terms of the characteristics of dynamic valgus? [J]. Am J Sports Med, 2014, 42(9):2095-2102.
[25] KELLIS E, ARABATZI F, PAPADOPOULOS C. Muscle co-activation around the knee in drop jumping using the co-contraction index [J]. J Electromyogr Kinesiol, 2003, 13(3):229-238.
[26] KOGA H, NAKAMAE A, SHIMA Y,. Mechanisms for noncontact anterior cruciate ligament injuries knee joint kinematics in 10 injury situations from female team handball and basketball [J]. Am J Sports Med, 2010, 38(11):2218-2225.
[27] KOMI P V, GOLLHOFER A. Stretch reflexes can have an important role in force enhancement during SSC exercise [J]. J Appl Biomech, 1997, 13(4):451-460.
[28] KRAMER A, RITZMANN R, GRUBER M,. Leg stiffness can be maintained during reactive hopping despite modified acceleration conditions [J]. J Biomech, 2012, 45(10):1816-1822.
[29] KRISTIANSLUND E, FAUL O, BAHR R,. Sidestep cutting technique and knee abduction loading: implications for ACL prevention exercises [J]. Brit J Sports Med, 2014, 48(9):779-783.
[30] KROSSHAUG T, NAKAMAE A, BODEN B P,. Mechanisms of anterior cruciate ligament injury in basketball: video analysis of 39 cases [J]. Am J Sports Med, 2007, 35(3):359-367.
[31] LESINSKI M, PRIESKE O, BEURSKENS R,. Effects of drop height and surface instability on neuromuscular activation during drop jumps [J]. Scan J Med Sci Sports, 2017, 27(10):1090-1098.
[32] LIN C F, CHEN C Y, LIN C W. Dynamic ankle control in athletes with ankle instability during sports maneuvers [J]. Am J Sports Med, 2011, 39(9):2007-2015.
[33] LINNAMO V, STROJNIK V, KOMI P V. Maximal force during eccentric and isometric actions at different elbow angles [J]. Eur J Appl Physiol, 2006, 96(6):672-678.
[34] MARKOLF K L, BURCHFIELD D M, SHAPIRO M M,. Combined knee loading states that generate high anterior cruciate ligament forces [J]. J Orth Res, 1995, 13(6):930-935.
[35] MOKHTARZADEH H, YEOW C H, GOH J,. Antagonist muscle co-contraction during a double-leg landing maneuver at two heights [J]. Com Met Biomech Biomedical Eng, 2017, 20(13):1382-1393.
[36] NEEDLE A R, BAUMEISTER J, KAMINSKI T W,. Neuromechanical coupling in the regulation of muscle tone and, joint stiffness [J]. Scan J Med Sci Sports, 2014, 24(5):737-748.
[37] NEPTUNE R R, WRIGHT I C, VAN DEN BOGERT A J. Muscle coordination and function during cutting movements [J]. Med Sci Sports Exerc, 1999, 31(2):294-302.
[38] PEROTTO A, DELAGI E F. Anatomical Guide for the Electrom-yographer: The Limbs and Trunk [M]. Charles C. Thomas, 1994.
[39] RUAN M, LI L. Approach run increases preactivation and eccentric phases muscle activity during drop jumps from different drop heights [J]. J Electromyogr Kinesiol, 2010, 20(5):932-938.
[40] SANTELLO M. Review of motor control mechanisms underlying impact absorption from falls [J]. Gait Posture, 2005, 21(1):85-94.
[41] SCHENAU G J V I, BOBBERT M F, HAAN A D. Mechanics and energetics of the stretch-shortening cycle: A stimulating discussion [J]. J Appl Biomech, 1997, 13(4):484-496.
[42] SOUISSI H, ZORY R, BREDIN J,. Comparison of methodologies to assess muscle co-contraction during gait [J]. J Biomech, 2017, 24(57):141-145.
[43] TAM N, SANTOSCONCEJERO J, COETZEE D R,. Muscle co-activation and its influence on running performance and risk of injury in elite Kenyan runners [J]. J Sports Sci, 2016, 35(2):175-181.
[44] TAUBE W, LEUKEL C, GOLLHOFER A. How neurons make us jump: the neural control of stretch-shortening cycle movements [J]. Exec Sport Sci Rev, 2012, 40(2):106-115.
[45] TAUBE W, LEUKELC, SCHUBERT M,. Differential modulation of spinal and corticospinal excitability during drop jumps [J]. J Neurophysiol, 2008, 99(3):1243-1252.
[46] THOMAS S J, SWANIK C B, HIGGINSON J S,. Neuromuscular and stiffness adaptations in division I collegiate baseball players [J]. J Electromyogr Kinesiol, 2013, 23(1):102-109.
[47] VANRENTERGHEM J, VENABLES E, PATAKY T,. The effect of running speed on knee mechanical loading in females during side cutting [J]. J Biomech, 2012, 45(14):2444-2449.
[48] YEADON M R, KATO T, KERWIN D G. Measuring running speed using photocells [J]. J Sports Sci, 1999, 17(3):249-257.
[49] YU B, LIN C F, GARRETT W E. Lower extremity biomechanics during the landing of a stop-jump task [J]. Clin Biomech, 2006, 21(3):297-305.
[50] ZAJAC F E. Muscle and tendon: properties [J]. Crit Rev Biomed Eng, 1989, 17(4):359-410.
[51] ZEBIS M K, ANDERSEN L L, BENCKE J,. Identification of athletes at future risk of anterior cruciate ligament ruptures by neuromuscular screening [J]. Am J Sports Med, 2009, 37(10):1967-1973.
The Muscle Activation Characteristics of Knee and Ankle During 45°Side-step Cutting Task under Different Approach Run Speeds
YUAN Peng1,2, XU Yi-lin1, WANG Dan3, ZHOU Su-po1,4, CHEN Wei1
1. Jiangsu Research Institute of Sports Science, Nanjing 210033, China; 2. Soochow University, Suzhou 215006, China; 3. Shanghai University of Sport, Shanghai 200438, China; 4. Nanjing SportInstitute, Nanjing 210014, China.
Objective: To compare the differences of muscle activation characteristics of knee and ankle between male and female participants under different approach run velocities. In addition, the correlation between pre co-activation ratio and post co-activation ratio of muscles in knee and ankle joint was further analyzed. Methods: Forty-six youth basketball and volleyball players completed 45° side-step cutting task at 3, 4 and 5 m/s, respectively. Delsys wireless surface electromyography (EMG) was used to collect the EMG signals of quadriceps (rectus femoris, vastus medialis and vastus lateralis), hamstrings (biceps femori long head and short head, semitendinosus), tibialis anterior and lateral gastrocnemius. The normalized root-mean-square amplitudes and the co-activation ratio of knee and ankle muscles in the pre-activation and post-activation phases at three approach run velocities were analyzed. Results: The approach run velocities significantly increased the activation levels of quadriceps and lateral gastrocnemius during the pre-activation phase (<0.05), the activation levels of quadriceps, hamstring, tibialis anterior and lateral gastrocnemius muscles during the post-activation phase (<0.05), but significantly reduced the muscle co-activation ratio of the knee and ankle in the pre-activation phase (<0.05). The activation level of quadriceps and tibialis anterior muscle in females were significantly higher than that of the males (<0.05) during pre-activation and post-activation phases, but muscle co-activation ratio of the knee in females during pre-activation and post-activation phases was significantly lower than in males (<0.05). Muscle co-activation ratio in knee and ankle was significantly correlated between pre-activation and post-activation phase (<0.05). Conclusions: With increasing approach run speed, central nervous system increased the muscle activation level by adjusting the pre-activation and post-activation mechanisms. There was moderate to high correlation between the pre-activation and post-activation mechanisms in the 45° side-step cutting task, which suggests that the coordination of pre-activation and post-activation can adjust the stiffness of muscles and joints to protect the joints and tissues of the lower extremities and to improve the athletic performance. The lower knee joint muscles co-activation ratio in post-activation phase quadriceps dominant activation pattern suggested that the females may suffer higher risk of ACL injury in 45° than their male counterparts. To some extent, gender difference in activation patterns of the knee muscles may explain the gender difference in ACL injury risk.
G804.6
A
10.16469/j.css.201808006
1000-677X(2018)08-0049-10
2018-04-13;
2018-08-13
江苏省体育局局管指向性课题(ST14019307)。
袁鹏,男,副研究员,在读博士研究生,主要研究方向为运动生物力学,E-mail:limid_yp@126.com。
