玉山黑猪两种头型遗传机制的初步解析
2018-09-01王成斌王小鹏郑小明张俊杰郭源梅丁能水
王成斌,王小鹏,郑小明,陈 浩,张俊杰,郭源梅,丁能水*
(1.江西农业大学省部共建猪遗传改良与养殖技术国家重点实验室,南昌 330045;2.国家级玉江猪(玉山黑猪)保种场,玉山 334700)
猪是继狗和羊之后第三个被人类驯养的家畜[1]。经过长时间的人工选择和自然选择,家猪形成了丰富的品种和多样化的体型外貌特征。玉山黑猪是江西省第一个被列入国家级畜禽遗传资源保护名录的地方猪种,具有早熟易肥,肉质细嫩、鲜美等特点,其全身被毛黑色。玉山黑猪的头型有两类:“狮子头”型和“马脸”型。“狮子头”型头大,额宽,且有较深的皱纹;嘴筒短略上翘,躯干背腰不平;腹下垂,体短,体型较小,体质较疏松。“马脸”型头中等大,躯干背腰平直,体长[2]。迄今为止,导致这两种头型的遗传机制尚未被解析。深入揭示玉山黑猪不同头型的形成机制对玉山黑猪种质特性形成机理研究、品种多样性的保护及利用具有重要的指导意义。
在人工选择的过程中,对目标基因的定向选择会导致种群的某一性状发生改变。目的基因优势等位基因频率在选择的作用下会不断提高,其附近与之紧密连锁的等位基因频率将伴随其一同升高,此现象称为选择性清除(selective sweep)。因此,受到选择的基因组区域会表现出选择性清除信号,如连锁不平衡值(linkage disequilibrium, LD)升高、核苷酸多态性(DNA polymorphism)下降等[3]。选择和遗传漂变都能导致遗传分化和选择信号的产生,表现为不同种群间等位基因频率差异显著。遗传分化系数(Fst)是检测群体遗传分化程度的重要指标,由Wright[4]
提出,用于计算物种亚群之间的分化程度。
全基因组关联分析(GWAS)自2005年首次报道应用于人视网膜黄斑变性遗传机理研究以来,其在人类复杂疾病研究领域取得了迅速发展。研究人员通过GWAS发现了大量与疾病相关联的遗传变异。在家猪中,自2011年第一篇应用GWAS研究猪毛色的文章报道以来[5],GWAS在家猪重要经济性状关联基因的鉴别中同样得到了广泛应用[6-7]。
本研究尝试利用猪全基因组60K SNP芯片数据,检测玉山黑猪“狮子头”和“马脸”两种头型种群间的遗传分化信号,并进行病例-对照的全基因组关联分析,初步筛选与头型相关的染色体关联区域,并在这些区域内发现令人感兴趣的候选基因,以期初步解析玉山黑猪形成不同头型的分子机制,为深入研究玉山黑猪及中国地方猪种群体内不同头型的形成机制奠定基础。
1 材料与方法
1.1 试验动物及表型测定
本研究选取国家级玉山黑猪保种场的36头原种猪,其中“狮子头”型猪12头,“马脸”型猪24头(图1)。在出生时测定初生重,断奶时测定断奶重,猪只180日龄时,测定其体重、体长、体高、背高、胸围、胸深、腹围、管围、腿臀围,并做好记录。

a.马脸型;b.狮子头型a. Long-pointed head; b. Short head图1 玉山黑猪不同头型个体Fig.1 Different head types in Yushan Black pigs
1.2 DNA提取、60K SNP判型及质控
采集耳组织样品,按常规的酚/氯仿法提取耳组织DNA,将其溶于TE缓冲液中,并统一稀释至50 ng·μL-1,用ILLumina猪60K SNP芯片进行扫描分型。芯片分型工作委托北京怡美通德公司按照标准操作流程完成。用Plink软件(Plink v1.07)对SNP进行质控,剔除个体检出率<90%、最小等位基因频率<0.05、极显著偏离哈代温伯格平衡(P<0.000 001)、染色体位置不确定的SNP位点和剔除SNP检出率<90%的个体,最后剩余26 480个SNPs用于后续的统计分析[8]。
1.3 统计分析
1.3.1 玉山黑猪两种头型间表型差异性分析 用R语言(R v3.3.1)中的aov()函数对所记录的11个性状进行组间方差分析[9]。将性别、批次(年、季)和180日龄体重作为协变量,比较这些表型性状在“狮子头”型和“马脸”型两组间是否存在显著差异。
1.3.2 玉山黑猪两种头型间遗传分化系数(Fst)的计算 用R语言(R v3.3.1)中的Genepop软件包计算“狮子头”型和“马脸”型两类群体之间每个SNP位点的遗传分化系数(Fst)值:
其中,MSG为群体内的观测均方差:
MSP为群体间的观测均方差:
nc是指校正后群体间的平均样本大小:

1.3.3 玉山黑猪两种头型的全基因组关联分析 采用病例-对照的全基因组关联分析,应用GEMMA(genome-wide efficient mixed-model association)软件[11]中的单变量线性混合模型(univariate linear mixed model)来比较全基因组上“狮子头”型个体和“马脸”型个体间等位基因及基因型频率的差异,搜寻与目标性状基因座处于连锁不平衡的位点。为了消除群体分层对GWAS结果的影响,GEMMA将提前生成的一个分子亲缘关系矩阵作为随机效应带入到模型中:
y=Wα+xβ+u+ε
式中,y为所有个体的表型值向量;W为协变量矩阵;α为包含截距的相应的系数向量;x为SNP的基因型向量;β为SNP的影响效应;u为随机效应向量;ε为误差向量。将表型对性别和批次(年、季)进行回归分析,并用残差作为新的表型用于上述分析[12]。
采用Bonferroni方法确定关联显著性阈值,基因组显著水平阈值为0.05/N,N为有效SNP个数,即0.05/26 480=1.89E-06。通过与最高点SNP连锁不平衡(LD)r2≥0.8的SNP位点确定置信区间。使用Haploview程序[13]生成单倍型及单倍型框。采用QQ-plot通过比较无效假设的分布与实际关联性分布的差异来检测是否具有群体层化效应。
1.3.4 玉山黑猪两种头型候选基因的鉴别参照SusscrofaBuild 10.2基因组注释版本(http://www.ensembl.org/Sus_scrofa/Info/Index),查找遗传分化显著区域和GWAS置信区间内的基因。利用GeneCards (http://www.genecards.org/) 数据库查找这些基因的生物学功能。为避免遗漏基因,针对家猪基因组中尚未注释的RefSeq基因,利用UCSC Genome Browser 数据库(http://genome.ucsc.edu/)选出在人和小鼠中注释的同源基因。
2 结 果
2.1 玉山黑猪两种头型群体间表型差异分析
本研究统计分析了玉山黑猪“狮子头”型和“马脸”型两个类群的11个表型性状(表1),发现“马脸”型猪比“狮子头”型猪的体长要长,体型要大,但差异未达到显著水平(P>0.05)。“马脸”型猪的管围显著大于“狮子头”型猪(P<0.05)。
2.2 玉山黑猪两种头型群体间遗传分化信号的检测
玉山黑猪两种头型的群体间总体上不存在明显的遗传分化,所得Fst值平均值为0.031(图2)。Fst值最高的10个SNP涉及11个基因(表2)。遗传分化最显著的选择信号位于10号染色体上,Fst值最大的SNP位点CASI0009931位于SSC10的11.36 Mb处(图3a)。

蓝线代表Fst的平均值The blue line indicates the average Fst value图2 所有SNP位点遗传分化系数(Fst)值的密度分布Fig.2 Distribution of genetic differentiation coefficient (Fst) values of all the SNPs

a.全基因组SNP的Fst值分布图;b. GWAS曼哈顿图:图中虚线代表5%基因组显著水平阈值a. Distribution of Fst values of all qualified SNPs across the genome; b. Manhattan plot of GWAS: The dashed lines indicate the 5% genome-wide significant threshold values图3 全基因组Fst值分布图和GWAS曼哈顿图Fig.3 The distribution of Fst values of SNPs across the genome and Manhattan plot of GWAS
2.3 玉山黑猪两种头型的全基因组关联分析
通过GEMMA软件对所有个体进行病例-对照的全基因组关联分析,并经Q-Q plot检验,结果显示,膨胀系数(λ)为1.037,表示没有明显的群体层化效应(图4)。我们在SSC10上检测到了2个基因组显著水平的SNPs,最高点SNP(ALGA0056978,P=5.38E-07)位于11.76 Mb处(图3b),与Fst最高值的SNP(CASI0009931,SSC10:11.36 Mb)相距仅400 kb。通过与最高点连锁不平衡(LD)r2≥0.8及单倍型分析,确定该位点的置信区间为11.76~12.05 Mb(图5)。


横坐标为预期P值对数的相反数(将负数转换为正数),纵坐标为计算P值对数的相反数X axis means the expected values of the opposite of exponential P values, the Y axis means the observed values of the opposite of exponential P values图4 Quantile-quantile (QQ) 图Fig.4 Quantile-quantile (QQ) plot
3 讨 论
近年来,中国地方猪因其优异的种质特性受到研究者的广泛关注,为科研工作在繁殖性状、肉质性状和遗传进化等领域提供了重要的研究材料[14]。在《中国畜禽遗传资源志·猪志》收录的猪种中,有18个猪种(东串猪、山猪、岔路黑猪、金华猪、仙居花猪、皖南黑猪、滨湖黑猪、杭猪、乐平猪、玉江猪、大蒲莲猪、确山黑猪、阳新猪、大围子猪、通城猪、德保猪、文昌猪、汉江黑猪)存在头型的分离。从其中心产区的分布地域来看,这些具有头型变异的猪种主要集中在长江中下游流域。不同头型的产生可能与长江中下游流域祭祀习俗有关。清吴谷人《新年杂咏》中写到“杭俗,岁终礼神尚猪首,至年外犹足充馔。定买猪头在冬至前,选皱纹如‘寿’字者,谓之‘寿头猪头’。”“狮子头”型猪符合了当时社会活动和人们祭祀的文化需求而受到了选择。
我们对玉山黑猪两种头型的群体进行了遗传分化信号检测,Wright[4]建议,Fst为0~0.05表明群体间遗传分化程度较小,可以忽略,Fst为0.05~0.15表明群体间分化程度为中等,Fst为0.15~0.25表明群体间分化程度较大,Fst大于0.25则表明群体间分化程度很大。本研究所得到的Fst平均值为0.031(图2),说明玉山黑猪两种头型的群体间总体上不存在明显的遗传分化。我们对玉山黑猪“狮子头”型和“马脸”型两个类群的11个表型性状进行了差异性分析,结果表明大部分性状(除管围外)均差异不显著。这些结果符合了玉山黑猪品种选育的实践-即无论是“狮子头”型还是“马脸”型都是按照相同的方向进行选育的,玉山黑猪并没有形成两种特定的、存在明显遗传分化和表型巨大差异的亚群。这为后续基于“病例-对照”的两种头型群体的全基因组关联分析奠定了基础。将仅存在头型差异的两组玉山黑猪群体进行了基于“病例-对照”的全基因组关联分析,结果显示,在10号染色体上检测到了与遗传分化信号相互重合的显著关联位点。通过连锁不平衡(LD)和单倍型分析,将置信区间定位在11.76~12.05 Mb(图5)。此区间内包含两个可能的候选基因:RAB3GAP2和IARS2。
RAB3GAP2(RAB3-GTP酶激活蛋白催化亚单位2)编码的蛋白质属于RAB3蛋白质家族,参与调节胞外分泌的神经递质和激素,可能参与神经发育过程如增殖、迁移和分化前突触的形成[15]。在人中,与RAB3GAP2相关的疾病有Martsolf综合征和Warburg Micro综合征,其症状表现为颅面部骨骼发育异常(颧骨扁平、高颚)和智力障碍[16]。Aligianis等[17]报道,RAB3GAP2基因的一个低倍纯合剪切突变在一个家族中造成Martsolf综合征。Borck等[18]报道,一个纯合移框突变造成一个土耳其女孩的Warburg Micro综合征。IARS2(异亮氨酰-tRNA合成酶2)编码蛋白可氨酰化与其tRNA同源的氨基酸,氨酰合成酶被认为是在进化过程中最早出现的蛋白质。在人中,与IARS2相关的疾病包括骨骼发育异常、生长激素缺乏症、白内障和感音神经性听力损失等[19-20]。GO注释表明,与该基因相关的生物学活动包括核苷酸装配和氨酰-tRNA编辑[21]。近年来多项研究表明,IARS2基因在人类中与多种疾病相关,如肥厚型心肌病[22]、非小细胞肺癌[23]、胃癌[24]等。Hägg等[25]利用全基因组基因荟萃分析认为,IARS2等基因与欧洲人肥胖相关。Gomes等[26]发现,IARS2等蛋白质合成和降解基因与内洛尔牛的重要经济性状相关。根据与这两个基因相关的人疾病综合征表现特征(骨骼发育,尤其是颅面部骨骼发育),笔者推测,RAB3GAP2和IARS2基因可能参与调控玉山黑猪两种头型的形成过程。验证这一推测需要更多深入的后续研究,如精细定位和多组学联合分析。
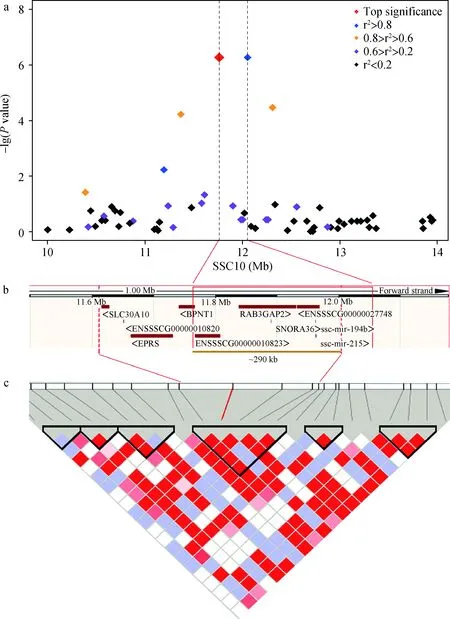
a.最显著位点附近4 Mb的区域GWAS,四边形块代表不同显著程度的SNP位点,不同颜色代表与最高点不同的连锁不平衡程度;b. 290 kb置信区间内的基因;c. 包含最高点的LD热图,最高点SNP用红线表示,一段包含最高点的392 kb单倍型框用黑色线框圈出a. Regional plot of a 4 Mb region around the most significant SNP, diamonds represent the SNPs with differet significance, the colored diamonds indicate different linkage disequilibrium (LD) extents between the top SNP and other SNPs; b. Genes within the 290 kb confidence interval; c. LD heatmap for the SSC10 locus, the top SNP is indicated in red line, a haplotype block with 392 kb harbors containing the top SNP are highlighted by black triangles图5 10号染色体上的显著关联位点Fig.5 The significant associated region on SSC10
4 结 论
本研究基于全基因组60K芯片数据,结合群体遗传分化分析和全基因组关联分析,在SSC10上鉴别到了与玉山黑猪两种头型形成显著关联的遗传位点。结合统计分析结果和目标区域内的基因功能,笔者推测,RAB3GAP2和IARS2是影响玉山黑猪头型的重要候选基因。该结果为玉山黑猪及中国地方猪种群体内不同头型形成机制的深入研究奠定了基础。
