尖峰岭热带山地雨林长期减弱的碳汇及其环境驱动因素分析
2018-09-01马素辉李意德陈德祥
李 翔 周 璋 马素辉 许 涵 李意德 陈德祥*
(1.中国林业科学研究院热带林业研究所,海南尖峰岭森林生态系统国家野外科学观测研究站,广州 510520; 2.北京大学城市与环境学院,北京 100781)
近几个世纪来由于人类活动的加剧,地球正以史无前例的速度经受着气候系统和地表结构的巨大变化[1]。温室效应的加剧[2]、降水格局的改变、极端气候事件发生频率和强度的增加[3]、氮沉降加剧[4]以及毁林和生境的破碎化等等,导致生物多样性急剧减少,生态系统结构与功能发生改变甚至退化,即使是保护最为完好的地区也无法避免气候变化带来的影响[5]。热带森林面积虽然只占陆地面积的7%~10%,植被面积的22%,但其但年碳交换量却高达400亿吨[6],生物量和土壤中储存的碳库高达4 600亿吨,是大气总碳库的一半还多[7]。由于世界热带森林复杂多样,热带森林生态系统是对气候变化的响应与适应研究中难度最大和了解最为不透彻的陆地生态系统类型之一[8]。近二十年来涌现了大量的学者着力开展气候变化情景下热带森林的响应与适应研究,初步发现气候变化导致降水格局改变而引起的干旱等极端气候事件是影响世界热带森林固碳功能最为重要的一个驱动因子[5,7~14]。
虽然热带森林固碳能力的环境影响机理研究方面已经取得了一定的进展,但就整体而言,仍然存在以下问题迫切需要进一步解决、补充和完善,才能更好的帮助人们了解热带森林固碳能力对气候变化的响应与适应机理,为人类应对气候变化提供更好的技术支撑。目前对热带森林固碳功能的环境影响机理研究主要针对南美亚马逊地区和非洲部分热带森林开展的。热带森林分布广阔,空间异质性强,不同地区的热带森林在环境因子、土壤状况、森林结构与类型、干扰历史等存在较大差异,由于缺乏足够多的世界范围热带森林的研究数据,尚不清楚降水格局改变形成的干旱或强降雨等极端事件对热带森林固碳能力的影响是否存在全球一致性?急需开展和补充更多热带地区,尤其是研究相对薄弱的亚洲热带森林对气候变化的响应与适应研究。
海南岛保存有我国天然林连片面积最大、最为典型的热带森林。由于地处热带亚洲北缘,又在物种组成和结构上与典型热带森林表现出明显的差异,龙脑香科(Dipterocarpaceae)物种较少,只有2~3种,但物种多样性却显著高于其他地区热带森林[15]。而且由于受季风性海洋气候的影响,旱雨季分明,旱季时间明显长于雨季时间,但约90%的年降水却集中在雨季。因此,鉴于我国海南岛热带森林的区位和环境条件的独特性,在当前气候变化情景下,海南岛热带森林固碳能力到底发生了怎么样的动态变化?驱动因素是什么?这些问题的回答将成为全球热带森林碳收支估算、碳循环对气候变化的响应与适应研究机理揭示不可或缺的重要补充。因此,本文通过对尖峰岭固定样地历史长期监测数据和降雨等环境参数数据的分析,开展海南岛热带森林碳循环及其各分量的长期动态研究,并结合对降水和干旱等环境因素的相关分析,试图探求海南岛热带森林固碳能力对气候变化的响应与适应机理。
1 研究区概况
研究样地位于海南省乐东市尖峰岭国家自然保护区内的山地雨林中。尖峰岭地处海南岛乐东县与东方县交界,地理坐标为18°20′~18°57′N,108°41′~109°12′E,林区总面积约为472.27 km2,森林覆盖率超过93%。林区内有丰富的森林植被资源,其中热带山地雨林是保存最为完整,结构最为复杂的森林类型。尖峰岭位于亚洲热带地区的北缘,属于热带季风气候,水热资源丰富,据尖峰岭海拔820 m气象站观测数据显示,年总辐射量为5 517.4 MJ·m-2,年平均气温为19.8℃,最低月平均气温和最高月平均气温分别为10.8℃和27.5℃,≥10℃的年平均积温7 204℃,平均相对湿度88%,年平均降水量2 449 mm,干湿季明显[16]。尖峰岭雨季受台风影响频繁,80%~90%的雨量集中在5~10月的雨季,其中暴雨总量约占全年降水总量的68%,特大暴雨占全年降水量的44.4%[16]。土壤类型为砖黄壤。
2 研究方法
2.1 研究样地选择
本研究选择3块尖峰岭长期固定样地(即P8401、P8402、P8901)来研究生物量长期变化动态,P8401(100 m×12 m)和P8402(100 m×10 m)为1984年设立并进行第一次调查。P8901(100 m×12 m)为1989年设立并进行第一次调查,林分以盘壳栎(Cyclobalanopsispatelliformis)、闽粤栲(Castanopsisfissa)、小叶白锥(Castanopsistonkinensis)等为主。样地选择尽量选择避免人为活动干扰,地形、母岩、土壤类型以及群落类型大致相同的样地,且能够进行长期的森林清查,不会出现中断情况。样地位于尖峰岭五分区原始林中,基本为南北(S/N)和东西(E/W)走向。
每个样地以10 m×10 m的小样方为基本单元,将小样方内的所有胸径≥1 cm的树木挂上标签,并记录每个植株的物种名称、坐标、胸径(DBH)、树高。胸径起测标准1984年为DBH≥7.0 cm,1992年为DBH≥5.0 cm,2000年以后为DBH≥1.0 cm,1.3 m处为板根的板根植物,测定胸径的方法选择在高于板根上端50 cm处测定,同时记录测定点高度。另外只需记录0~2.5 m高处有一点DBH≥10 cm的藤本植物,并记录该藤本是否延伸至样地外。
样地调查时间:P8401样地为1984,1989,1991,1995,1998,1999,2003,2005和2013年;P8402样地为1984,1989,1991,1995,1998,1999,2003,2005和2013年;P8901样地为1989,1991,1995,1998,1999,2003,2005和2013年。
由于P8901样地从1989年开始调查,缺乏1984年调查数据,故本研究中P8901样地1984年生物量数据使用P8401和P8402样地的均值代替。
2.2 森林生物量及碳密度的估算
利用样地清查数据估算林木(包括大的棕榈科(Palmae)及藤本植物)生物量。由于热带山地雨林结构复杂,优势种不明显,同种个体数目较少,所以无法使用单一的异速生长方程,而必须利用多树种的混合回归模型进行生物量估算。本研究中单株个体的生物量采用已知的尖峰岭山地雨林混合模型[17]进行计算:
树干:Wt=0.022 816(D2H)0.992 674
(1)
树皮:Wbk=0.006 338(D2H)0.902 418
(2)
树枝:Wbr=0.005 915(D2H)0.999 046
(3)
树叶:Wl=0.005 997(D2H)0.804 661
(4)
树根:Wr=0.003 612(D2H)1.115 27
(5)
式中:W为各器官生物量;D为胸径(DBH);H为树高(Height)。
由于样地在不同年份清查过程中,起测径级并不一致。为了便于比较,本文选择DBH≥7.0 cm的林木个体进行生物量计算。碳密度是指单位面积的生物量碳含量,估算采用方程:C=生物量密度×平均碳含量,平均碳含量采用已知的尖峰岭热带山地雨林平均碳含量50.73%[17]。碳密度年变化率为两次调查碳密度的年均差值,用以表征碳源汇的变化动态。
2.3 森林生物量各分量的计算
生物量净变化(biomassnetchange):
N=(biomassn2-biomassn1)/t
(6)
式中:N是生物量净变化;biomassn2是后一次调查总生物量;biomassn1是前一次调查总生物量;t是两次调查间隔时间,以年计。
死亡生物量(biomassmortality):
M=biomassdying(n2-n1)/t
(7)
式中:M是死亡生物量;biomassdying(n2-n1)是两次调查期间死亡个体总生物量;t是两次调查间隔时间,以年计。
新增生物量(biomassrecruitment):
R=biomassrecruit(n2-n1)/t
(8)
式中:R是新增生物量;biomassrecruit(n2-n1)是两次调查期间新增个体总生物量;t是两次调查间隔时间,以年计。
新增个体平均生物量(Growthgainsperstem):
G=R/nrcruit
(9)
式中:G是新增个体平均生物量;nrcruit是两次调查期间新增株数。
死亡个体平均生物量(Lossesperstem):
L=M/ndying
(10)
式中:L是死亡个体平均生物量;ndying是两次调查期间死亡株数。
2.4 气象数据分析
2.4.1 气象数据的获取
气象资料来自海南尖峰岭国家级森林生态系统定位站气象观测场1984~2013年的地面常规气象资料。气象观测场位于18°44′25″N,18°51′30″E,海拔820 m。观测场按国家地面气象站标准设计:南北和东西边长35 m×25 m,均质草皮地面,四周距离天然林保持约20 m以上。
2.4.2 直接环境因子的统计
日降水量、月总降水量、年总降水量、日平均蒸散量与年总蒸发率、极端降雨日期和总降雨雨量。
2.4.3 环境因子评价指数的计算
水分最大亏缺指数(MCWD)[8]:
MCWD定义为水分最大亏缺指数,通过月总降水量P(mm/month)与月总蒸发量E(mm/month)的差值来评定,当月蒸发量大于月降雨量时,则认为当月存在水分亏缺,且水分亏缺值越大,表示水分亏缺状况越严重。以第n月为例:
If Pn-En<0,ThenMCWDn=Pn-En,Else MCWDn=0
MCWD=MCWD1+MCWD2+…+MCWD12
不同等级暴雨事件:
记录研究期间以来的暴雨次数(暴雨划分采用国家气象局降水强度等级划分标准,选用24 h雨量>50 mm的暴雨等级与24 h雨量>100 mm的大暴雨等级)及暴雨对应日期。分别统计一年内24 h降雨量>50 mm以及24 h降雨量>100 mm的雨量总和。
2.5 数据处理
利用Excel 2013及SPSS 23.0对原始数据进行初步整理,计算及统计分析。采用R3.3.3 mgcv软件包的gam函数模拟生物量各分量长期动态变化的广义加性模型(GAM)。
3 结果与分析
3.1 尖峰岭山地雨林森林生物量长期动态变化
基于3块样地森林生物量总量及净变化长期动态变化显示,从1984~2013年尖峰岭山地雨林生物量密度在逐年增长(图1b),表明近年来尖峰岭山地雨林是作为一个稳定的碳汇存在,但生物量净变化呈现减少的趋势(图1a),表示其固碳能力正在逐渐减弱。尖峰岭山地雨林的生物量密度在320.20±40.37和360.73±46.83 mg·hm-2范围内波动,均值为346.14±13.15 mg·hm-2。生物量碳密度在162.44±20.48和182.99±23.75 mg·C·hm-2范围内波动,均值为175.60±6.67 mg·C·hm-2。尖峰岭平均碳密度年变化率为0.71±0.22 mg·C·hm-2·a-1,表示虽然其固碳能力在逐渐减弱,但仍属于碳汇能力较强的森林类型(表1)。
3.2 尖峰岭山地雨林死亡生物量长期动态变化
基于3块样地估算的死亡生物量长期动态变化显示,从1984~2013年尖峰岭山地雨林死亡生物量呈显著(P<0.01)的增加趋势(图2a),同时死亡的株数(图2b)以及死亡个体平均生物量(图2c)都呈增加(P<0.05)的趋势。这可能是由于近几十年来由于全球气候变化的加剧,导致极端气候事件(比如台风、暴雨以及季节性干旱等)的发生也更加频繁,一方面由于台风的影响不但使更多的大树发生倒伏、折断、死亡。同时,季节性干旱的发生也可能使得一些小树发生水分胁迫,从而枯死,使得死亡株数和死亡生物量都呈显著增加。例如2003年的达维台风导致尖峰岭山地雨林死亡生物量显著增加(图2a),但由于不同样地间表现出的地形差异及其他局地环境条件差异,使不同样地受台风影响程度各有不同,从而导致3个样地的死亡生物量存在较大差异,表现为置信区间幅度较大。
表1不同年份尖峰岭山地雨林生物量密度、生物量密度年变化率、生物量碳密度及碳密度年变化率
Table1Biomassdensity,annualrateofbiomassdensitychange,biomasscarbondensityandannualrateofbiomasscarbondensitychangeintropicalmountainrainforestofJianfenglingindifferentyears
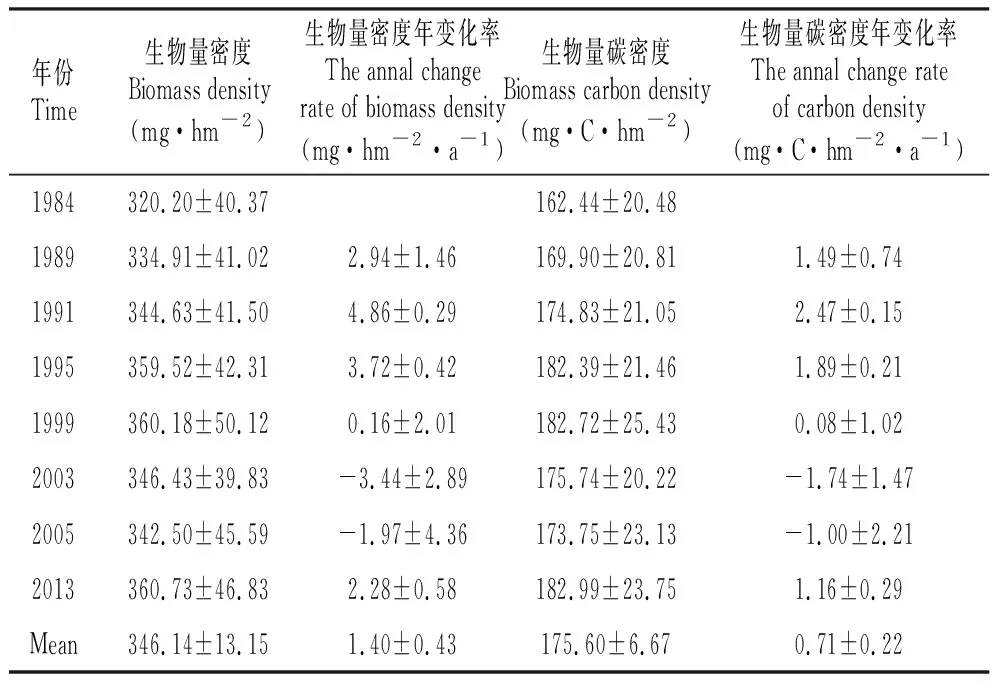
年份Time生物量密度Biomass density(mg·hm-2)生物量密度年变化率The annal change rate of biomass density(mg·hm-2·a-1)生物量碳密度Biomass carbon density(mg·C·hm-2)生物量碳密度年变化率The annal change rate of carbon density(mg·C·hm-2·a-1)1984320.20±40.37162.44±20.481989334.91±41.022.94±1.46169.90±20.811.49±0.741991344.63±41.504.86±0.29174.83±21.052.47±0.151995359.52±42.313.72±0.42182.39±21.461.89±0.211999360.18±50.120.16±2.01182.72±25.430.08±1.022003346.43±39.83-3.44±2.89175.74±20.22-1.74±1.472005342.50±45.59-1.97±4.36173.75±23.13-1.00±2.212013360.73±46.832.28±0.58182.99±23.751.16±0.29Mean346.14±13.151.40±0.43175.60±6.670.71±0.22
注:95%置信区间,SE由P8401、P8402、P8901三个样地结果计算得出。
Note:The confidence interval is 95%,and SE is composed of the results of the plots of P8401,P8402 and P8901.
3.3 尖峰岭山地雨林新增生物量长期动态变化
基于3块样地估算的新增生物量长期动态显示,从1984~2013年尖峰岭山地雨林新增生物量和新增加个体的株数(图3b)有一个显著(P<0.01)的增长趋势(图3a),而且单株个体生物量(图3c)也呈现增加的趋势。但是与死亡量的年际变化较为均一相比,新增个体生物量的增加趋势却表现出明显的年际波动。一方面是由于大树倒伏后光热等环境资源的显著改善,导致一些较小的受压迫体获得充足的环境资源以致其可以得到快速的生长;但是另一方面,若是死亡的多为中小个体,并不会使得周围个体的光热等环境资源发生显著改善,因此这些个体的生长仍然无法获得充足的资源,表现出新增加个体数量和生物量都不会大幅增加,从而导致新增加个体的生长出现比较大的年际波动。
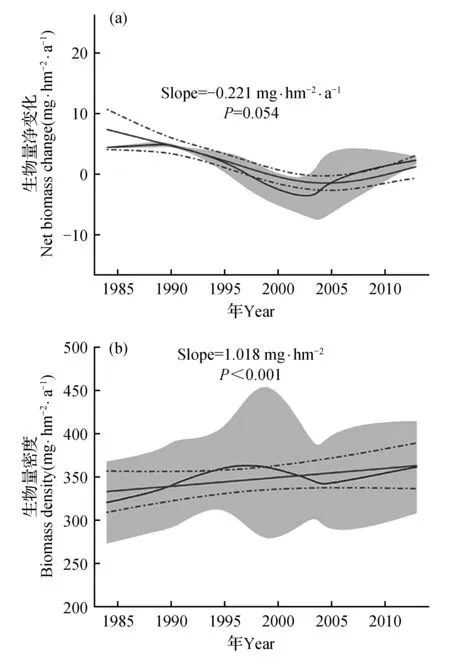
图1 所有样地生物量净变化及生物量密度的动态变化 a.生物量净变化;b.生物量密度 灰色线条代表3个样地的均值,灰色区域代表95%的置信区间,红色实线代表广义加性模型(GAM)拟合的均值,红色虚线代表标准误差,下同。Fig.1 Trends in net above-ground biomass change and biomass stock across all sites a.Net biomass change; b.Biomass stock Gray lines show the overall mean for 3 plots, and shaded area denote confidence interval(95%).The red lines indicate the overall mean of the long-term trends using generalized additive models(GAM),red broken lines denotes SE,the same as below.

图2 死亡生物量、死亡个体数及死亡个体生物量动态变化 a.生物量年均致死量;b.调查期间年均死亡株数;c.死亡个体年均生物量Fig.2 Trends in biomass mortality stem number dying and losses per stem across all sites a. Biomass mortality; b. Stem number dying; c. Losses per stem
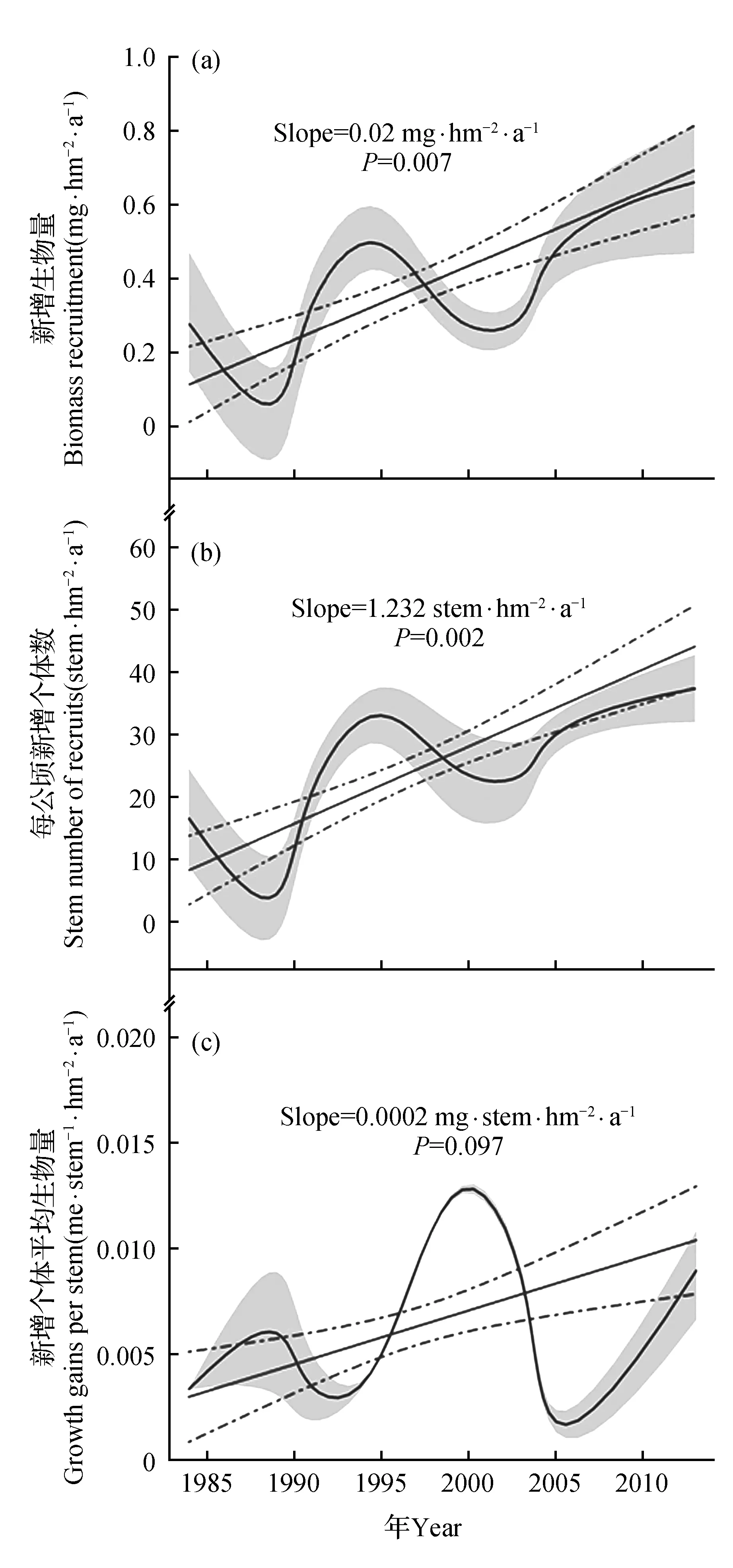
图3 新增生物量、新增个体数及新增个体生物量动态变化 a.生物量年均补充量;b.调查期间年均新增株数;c.新增个体年均生物量Fig.3 Trends in biomass recruitment, stem number of recruits and growth gains per stem across all sites a.Biomass recruitment;b.Stem number of recruits;c.Growth gains per stem
3.4 极端气候环境因子评价指数长期动态变化
3.4.1 水分最大亏缺指数(MCWD)长期动态变化
尖峰岭MCWD值在223.22和618.56 mm之间波动,平均值为371.19 mm。表现干旱较为强烈的年份为1989、1993、1994及2008年,MCWD值均大于500.00 mm。尖峰岭1980~2013年水分最大亏缺指数变化动态表现为:年际波动幅度大,并无明显的变化规律,总体呈现出弱增加的变化趋势(图4a)。
3.4.2 不同等级暴雨事件长期动态变化
通过分析尖峰岭山地雨林1980~2013年的50及100 mm两个不同等级暴雨事件年总暴雨降水量及年总暴雨天数动态变化。得出结论:在50 mm等级暴雨事件中,年总暴雨降水量在202.80和2 620.50 mm之间波动,平均值为1 258.32 mm(图4b)。其中年总暴雨降水量较多的年份分别是1980、1991、1992、1994、2003、2005及2011年。在100 mm等级暴雨事件中,年总暴雨降水量在206.20和2 334.00 mm之间波动,平均值为833.01 mm(图4c)。其中年总暴雨降水量较多的年份分别是1980、1991、1994、1996、2005及2011年。尖峰岭1980~2013年每年总暴雨降水量变化动态表现为:年际波动幅度大,并无明显的变化规律,在50 mm暴雨等级及100 mm暴雨等级两个尺度范围内,年总暴雨降水量均保持较为稳定的动态平衡。

图4 水分最大亏缺指数(a)、50 mm暴雨事件(b)年降水量及100 mm暴雨事件(c)年降水量动态变化Fig.4 Trends in Maxmium climatic water deflict(a),Annual precipitation of 50 mm storm events(b) and Annual precipitation of 100 mm storm events(c)
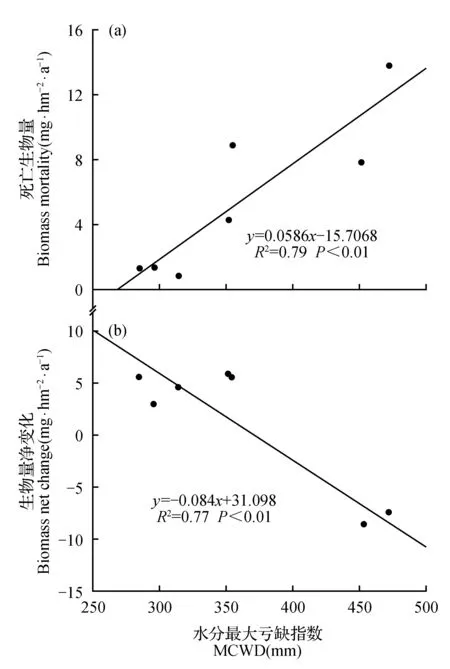
图5 水分最大亏缺指数(MCWD)与死亡生物量及生物量净变化的相关性Fig.5 The relationship between MCWD and biomass mortality,net biomass change

图6 不同等级暴雨事件与死亡生物量及生物量净变化的相关性Fig.6 The relationship between different levels of heavy rain events and biomass mortality,net biomass change
4 讨论
虽然尖峰岭山地雨林基本可以认为是个碳汇,但影响其动态变化的机制却不清楚。因此本文将从干旱、暴雨事件及台风等方面来对尖峰岭热带山地雨林生物量长期动态变化的驱动机理进行分析。
4.1 干旱对生物量长期动态变化的影响
干旱将导致热带森林个体死亡率显著上升[5,8,11,18~20]。从理论上来说,由于干旱抑制了光合作用导致代谢所需的碳底物供应不足,同时又会增加非结构性碳水化合物的消耗,导致碳饥饿发生,从而引发个体死亡[21];或者由于木质部水分传导的恶化导致气体交换、光合作用以及韧皮部传输等关键过程的限制使得组织脱水,从而导致个体死亡[22]。虽然尖峰岭热带森林仍然具备一定的碳汇能力,但由于干旱等极端气候事件导致个体死亡率的增加,使得热带森林的碳汇能力呈现下降的趋势。本研究通过对水分最大亏缺指数(MCWD)与死亡生物量和生物量净变化进行回归分析,结果显示水分最大亏缺指数(MCWD)与死亡生物量呈显著正相关(P<0.01);与生物量净变化呈显著负相关(P<0.01)(图5)。说明随干旱程度增加,死亡生物量会显著增加,生物量净变化则显著减少,表现出干旱对尖峰岭热带森林固碳能力具有明显的抑制作用。干旱对尖峰岭热带森林的这种影响可能是由于季节性干旱条件下,由于降雨减少,甚至长期无降雨,伴随蒸散强烈,土壤水分补充不足,产生干旱胁迫,直接影响植物的生理特性,抑制植物生长,甚至出现碳饥饿或组织脱水,从而使树木的死亡率显著增加,最终导致地上生物量显著减少。
4.2 不同等级暴雨事件对尖峰岭山地雨林生物量长期动态变化的影响
降雨是影响陆地生态系统的重要气候因子,NPP(净初级生产力)会随降雨量的升高而不断增加[23]。但超过一定降雨量临界值的暴雨及特大暴雨是否仍然会使树木生长产生正的效应?为探寻这个问题,本研究取每年内24 h降雨量>50 mm以及24 h降雨量>100 mm的暴雨事件,统计并分析降雨量与死亡生物量及生物量净变化之间的相关性。结果表明无论是24 h降雨量>50 mm,还是24 h降雨量>100 mm的暴雨事件,暴雨雨量与死亡生物量都呈显著正相关(P<0.01);与生物量净变化呈显著负相关(P<0.01)(图6)。在24 h降雨量>50 mm暴雨事件中,当年暴雨雨量低于1 800 mm时,暴雨并不会对死亡生物量与生物量净变化产生显著影响,而当暴雨雨量大于1 800 mm时,暴雨事件则对死亡生物量和净变化产生明显的负反馈作用。在24 h降雨量大于100 mm暴雨事件中,高于1 500 mm临界值后,死亡生物量同样大幅增加,生物量净变化大幅减少。说明当暴雨总雨量大于一定临界值以后,随暴雨雨量进一步增加,死亡生物量会显著增加,生物量净变化则会显著减少。强暴雨对NPP产生的负效应可能是源于降雨带来的多云量导致辐射输入的减少、土壤营养元素由于淋溶效应的增加而逐渐流失以及土壤孔隙氧气成分的减少所带来的一种综合反应。另外强暴雨往往伴随着台风的登陆,对森林生物量的影响表现为与大风共同作用的双重效应,产生较多大树的倒伏、折断和死亡。
4.3 台风对尖峰岭山地雨林生物量长期动态变化的影响
由于温室效应加剧,全球异常气候现象频发。海南岛尖峰岭临近海边,地处我国南端台风多发地段,则更易受台风侵害[24]。台风是影响尖峰岭山地雨林的一个重要自然干扰因子。对海南岛有影响的台风及热带风暴,平均每年有8.0个,其中强台风平均每年有2.7个[25]。台风带来的强降雨及大风,会对树木造成严重的生理危害与物理损伤,如树木倒伏等。台风对森林带来的损害可以分为直接性损害和间接性损害。直接性损害表现为乔木层大径级植株个体受损,间接性损害表现为风倒木作用于其他幼树层及下木层,导致其他植株死亡。例如2005年的达维台风,尖峰岭山地雨林共受损株数达到514株(占总株数的26.1%);其中风倒木达到206株(占总株数10.5%),DBH≥50 cm的风倒木占台风前该径级个体总株数比例最大,其次为7.5~20 cm胸径范围[24],表明大径级个体易受台风侵害。由于尖峰岭DBH≥10 cm以上的个体生物量占了90%以上,尤其是DBH≥45 cm的大径级个体,生物量所占比例高达32%,占据绝对的主导地位。因此台风导致的大径级个体死亡对生物量及碳密度变化具有显著的影响作用。
5 结论
基于1984~2013年3块样地固定清查数据显示,海南岛尖峰岭山地雨林单位面积平均碳汇速率为0.71±0.22 mg·C·hm-2·a-1,虽然整体还是作为一个长期稳定的碳汇存在,但其固碳能力在逐渐减弱。碳汇能力的减弱主要是源于干旱及台风暴雨等极端气候事件所导致的死亡生物量显著增加,而不同样地间存在明显差异,表示极端气候对热带森林固碳能力的影响也与样地间自身局地环境条件相关。
致谢感谢海南尖峰岭国家级自然保护区、中国林业科学研究院热带林业研究所尖峰岭试验站等单位在野外调查中给予的支持。
