内侧前额叶的小清蛋白神经元调控抑郁样行为
2018-08-30耿飞陈远寿罗芸田丹卢癸凤
耿飞 陈远寿 罗芸 田丹 卢癸凤
遵义医学院1生理学教研室,2病理生理学教研室(贵州遵义 563000)
低沉的情绪、缺乏动机或希望和快感缺乏是重症抑郁的常见症状[1]。临床抑郁症治疗的副作用提示需要进一步加深对抑郁行为的理解。之前的研究表明,重症抑郁和前额叶的紊乱紧密相关[2]。电刺激内侧前额叶可以诱导出抗抑郁样的行为[3]。而且,激活内侧前额叶的兴奋性神经元可以调控焦虑抑郁样行为[4]。绝经后抑郁的女性存在突触传递的改变[5]。这些发现显示,抑郁可能是内侧前额叶的细胞或者突触改变所导致的。
在过去20~30年中,分子、细胞、基因和成像技术的发展,让人们对包括焦虑、抑郁在内的社会行为有了更深入的理解[6-7]。而基于动物行为的研究已经成为生物学和生物医学领域研究社会行为及其神经机制的基础[8-9]。前额叶及其众多的交互连接在社会行为中发挥了重要的角色[10-11]。
内侧前额叶存在不同种类的中间神经元,这些中间神经元和局部的锥体神经元进行联系。之前的研究[12]表明,用Hm4Di抑制内侧前额叶的小清蛋白神经元能促进无助行为。之前的研究[13-14]证据也显示了抑郁症和内侧前额叶中间神经元受损的功能及数目的减少紧密相关,因此,前额叶小清蛋白神经元可能参与抑郁相关行为。
为了验证此假说,实验中选择性地在内侧前额叶小清蛋白神经元调控其活动,通过悬尾实验、糖水偏好实验及旷场试验来检测小鼠的抑郁样行为及运动能力。本项研究拟明确内侧前额叶小清蛋白神经元在抑郁行为中的作用。
1 材料与方法
1.1 实验动物PV⁃cre转基因小鼠(7~9周;Jackson Labs,B6;129P2⁃Pvalbtm1(cre)Arbr/J)被用于此项研究。所有的小鼠在8周龄时进行实验。6只小鼠一笼,12 h的光/暗周期(开灯时间从早上8:00到下午8:00)。小鼠随意进水进食。
1.2 病毒腺相关病毒5型包装的病毒,滴度大于2 × 1012/mL。AAV⁃DIO⁃ChR2⁃EGFP和AAV⁃DIO⁃EGFP的病毒购自上海生博公司。
1.3 方法
1.3.1 立体定位手术,病毒注射和套管埋置小鼠用氯胺酮(100 mg/kg),去除头顶毛发后固定在立体定位仪上。手术钻孔开颅,用33⁃G(Hanmil⁃ton)的针头给予0.5μL的AAV5病毒到双侧的内侧前额叶(2,±0.4,-1.5)。10μL汉密尔顿的微量注射器(nanofil;WPI,Sarasota,FL)在微量注射泵(UMP3;WPI,Sarasota,FL)的控制下被用来注射腺相关病毒。注射速率0.1μL/min。随后埋管,比病毒注射位点高0.5 mm(2,0,-1.0),离子水门汀粘合,然后用套管帽封闭套管。行为学实验在埋管后的3周进行。
1.3.2 旷场实验旷场测试箱(40 cm×40 cm)分为一个中央区域和外部。单个小鼠放在外围的区域,并且动物的运动路径被摄像机记录。运动总距离和在中央区域所待时间通过软件进行分析,运动轨迹通过上海吉量公司行为软件分析。
1.3.3 糖水偏好实验小鼠事先禁水过夜,糖水偏好检测箱被用来检测小鼠吸吮糖水或普通水的次数。糖水的浓度是1%。
1.3.4 悬尾实验该实验采用MED Associates悬尾设备对小鼠的运动状态进行实时监测。悬尾箱大小为:32 cm×33 cm×33 cm。用胶布制作成小钩,将4只小鼠尾部(约2 cm处)粘住并悬挂于压力传感器处,计算机自动记录每只小鼠在每分钟内的累计不动时间。
1.3.5 脑片准备小鼠先用乙醚麻醉,迅速的剪断颈部,在冰冻的人工脑脊液上取出大脑。用震动切片机(VT 1000S;Leica)切下包含内侧前额叶的横切片。切下脑片后,脑片被放置在32℃的记录液中恢复活性。所有的溶液都通有95%O2/5%CO2(V/V)的混合气。
1.3.6 体外膜片钳电生理记录脑片被放置在记录槽内,细胞钳制在-70 mV。记录抑制性电流的电极內液包含的成分为:140 mmol/L CsCl,2 mmol/L MgCl2,1 mmol/L CaCl2,10 mmol/L EGTA,10 mmol/L HEPES⁃CsOH,2 mmol/L adenosine triphosphate,5 mmol/L QX⁃314。记录兴奋性突触后电流时的电极内液的成分为:130 mmol/L K⁃gluconate,10 mmol/L NaCl,10 mmol/L Hepes,2 mmol/L MgCl2,1 mmol/L EGTA,0.13 mmol/LCaCl2·2H2O,3.5 mmol/LMg⁃ATP,1 mmol/L Na⁃GTP(pH 7.35,285 mOsm)。所有的试剂购买自Sigma或Tocris公司。
1.3.7 免疫荧光将小鼠用水合氯醛麻醉,灌注固定的小鼠脑袋用冰冻切片机进行切片,切片厚度25~40μm。脑片首先使用含有0.3%Triton X⁃100的牛血清白蛋白(BSA)封闭1.5 h,接着4℃孵育小清蛋白一抗(Swant235;1∶1 000)过夜。第2天加入荧光二抗(ALEXA 594⁃Goat anti Rabbit 1∶800)。室温避光孵育1 h,封片,荧光显微镜或共聚焦显微镜下观察。
1.4 统计学方法数据采用SPSS 13.0的统计软件包处理,多组间比较采用双因素方差分析,组间两组比较采取方差分析后的多重比较,取α=0.05为检验标准。
2 结果
2.1 内侧前额叶携带光敏感通道病毒的表达首先对PV⁃cre小鼠的内侧前额叶脑区注射腺相关病毒5型(AAV5)病毒,该病毒ChR2下游插入增强型的绿色荧光蛋白,不含有光敏感通道的病毒作为对照病毒。荧光实验发现红色荧光可以和绿色荧光共表达,而且红色荧光细胞多于绿色荧光细胞(图1A)。
473 nm蓝光刺激脑片可以记录到携带荧光蛋白的神经元的光控诱导电流(图1B),并且可以随着频率的增加(5、10和20 Hz)而稳定的诱导(图1C)。激活内侧前额叶带荧光的神经元同样诱导出动作电位,而且也可以随着频率的增加(5、10和20 Hz)而稳定的诱导出来(图1D)。而对照病毒表达后,记录不到这些电生理结果(未列出)。这些结果表明了光敏感通道成功表达于内侧前额叶小清蛋白神经元。
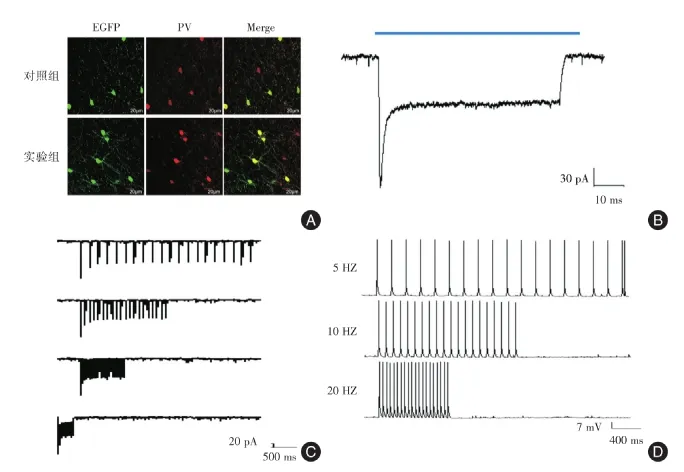
Fig.1 Expression of ChR2⁃EGFPvirus in mPFC图1 内侧前额叶中ChR2病毒的表达
2.2 悬尾实验为了探索内侧前额叶小清蛋白神经元在抑郁样行为中的作用,实验采用光控遗传学手段,通过悬尾实验来检测小鼠的行为学效应。首先比较了接受光控激活照射小鼠(PV:ChR2)和相应的对照小鼠(PV:EGFP)(图2A)。激活内侧前额叶的小清蛋白神经元可以减少悬尾实验中小鼠的不动时间(图2B:F=7.37,P=0.015;F=4.68,P=0.046。图2C:F=12.98,P=0.002 6)。
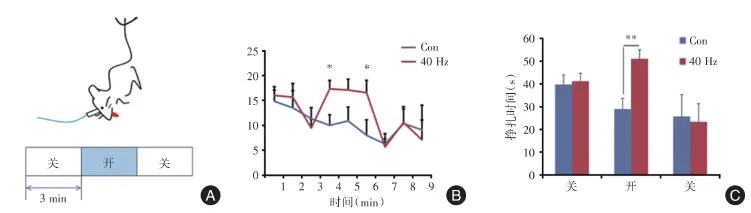
图2 兴奋内侧前额叶小清蛋白神经元对悬尾实验的影响Fig.2 Effect of excitation of the mPFCparvalbumin neurons in the tail⁃suspension test
2.3 旷场试验为了排除两组小鼠之间运动能力不同所导致的结果差异,应用旷场实验比较了接受光控激活照射小鼠(PV:ChR2)和相应的对照小鼠(PV:EGFP)的运动能力。结果显示,激活内侧前额叶的小清蛋白神经元对小鼠的运动功能并没有受到影响(F=1.67,P=0.33),两组小鼠的运动曲线比较接近。见图3。
2.4 糖水偏好实验为了探索内侧前额叶小清蛋白神经元在糖水偏好中的作用,采用糖水偏好实验来检测小鼠的行为。90 min的实验时程里,中间30 min给予光照(图4A)。结果显示,激活内侧前额叶的小清蛋白神经元可以增加小鼠的糖水偏好(F=5.35,P=0.039;图4B)。

图3 兴奋内侧前额叶小清蛋白神经元对旷场实验的影响Fig.3 Effect of excitation of the mPFCparvalbumin neurons in the open field test

图4 兴奋内侧前额叶小清蛋白神经元对旷场实验的影响Fig.4 Effect of excitation of the mPFCparvalbumin neurons in the sucrose preference test
3 讨论
人的前额叶被分为眶额叶皮层、背外侧前额叶、腹外侧前额叶及内侧前额叶,它们参与了众多的社会行为过程[15-16]。其中,内侧前额叶在人的社会认知等行为中发挥了关键的角色[17-18]。显而易见,缺乏内侧前额叶或前额叶紊乱会导致社会行为受损,产生类似焦虑、抑郁样等行为疾病[19-20]。
在本研究中,探讨了内侧前额叶小清蛋白神经元在抑郁样行为中的影响。首先,为了特异性调控小清蛋白神经元的活动,实验中将Cre酶依赖性表达的携带有光敏感通道的病毒表达在PV⁃cre转基因小鼠上。体内行为学结果显示,激活了小清蛋白神经元后小鼠表现为抗抑郁样效应:小鼠在悬尾实验中挣扎的时间增加,并且在糖水偏好实验中表现了更强的糖水偏好。而旷场实验的结果表明这些结果并不是由于不同组间小鼠运动能力的不同导致的。
免疫荧光结果显示,病毒所携带的荧光主要表达在小清蛋白神经元上。然而,荧光蛋白的表达并不能直接证明光敏感通道成功表达在细胞膜上。离体脑片光刺激可以记录到光控诱导电流和动作电位,为内侧前额叶中光敏感的成功表达提供直接证据。因此保证了行为上的可靠性。
激活小鼠内侧前额叶小清蛋白神经元表现了抗抑郁样的效应,这与之前的文献报道是相吻合的。之前的研究发现,增强锥体神经元的兴奋性传递促进无助行为[21],这显示了内侧前额叶局部脑区兴奋性和抑制性失去平衡,这可能是由于小清蛋白等中间神经元或者锥体神经元结构功能的改变所导致。而利用Hm4Di抑制内侧前额叶的小清蛋白神经元则更加直接地表明了小清蛋白神经元能促进无助行为[12],当然此无助行为模型与本研究的检测方法不同。
皮层小清蛋白神经元深刻地影响着锥体神经元发放的时程及模式,从而参与神经网络震荡的产生和维持[11]。文献报道内侧前额叶小清蛋白神经元在恐惧记忆和防御反应[22]等过程中发挥着重要角色,关于小清蛋白神经元在不同行为中的作用还有待于进一步的研究。然而,此研究应用新的方法,特异性地调控局部脑区的特定神经元,从而揭示了内侧前额叶小清蛋白神经元在抑郁样行为的作用。
本研究揭示了内侧前额叶小清蛋白神经元在无助行为及快感缺乏中的效应,从而强烈提示了其参与了抑郁相关行为的进程,而具体的作用机制还有待于进一步的探讨。
