STAT3对宫颈癌HeLa细胞的增殖、凋亡及自噬的影响
2018-08-30张慧蓉申东翔罗敏李军鹏杨耀
张慧蓉 申东翔 罗敏 李军鹏 杨耀
1广州军区广州总医院妇产科(广州510010);2广州中医药大学(广州510006)
宫颈癌是育龄期女性生殖系统最常见的恶性肿瘤之一,居女性恶性肿瘤死亡第2位,且发病率逐年升高及呈年轻化趋势,药物和妇科手术治疗对肿瘤虽取得一定疗效,但中晚期恶性肿瘤普遍预后不良的局面并未改变[1]。近年来对宫颈癌的研究不断深入,但宫颈癌的具体发病机制仍不明确。因此,目前迫切需要进一步深入探讨宫颈癌的具体发病机制,为宫颈癌尤其是晚期及复发性宫颈癌的治疗与预后提供新思路。信号转导和转录活化因子3(signal transducers and activators of transcription,STAT3)是一种重要的调控基因,对宫颈癌细胞具有重要的调控作用,与宫颈癌的复发、淋巴转移及患者预后等明显相关,能够引起细胞自噬调控的异常从而影响肿瘤的进展[2]。同时有大量的研究表明,自噬与宫颈癌细胞的增殖、分化及转移等有密切联系[3]。细胞自噬在进化上具有高度保守性,其发生过程受自噬相关基因的调控[4]。
近年来有研究表明STAT3对宫颈癌HeLa细胞的增殖与凋亡具有重大影响,主要发挥促癌基因的作用,促进宫颈癌HeLa细胞的增殖,抑制其凋亡[5-6]。同时研究表明,术后宫颈癌的局部复发、远处转移及耐药性的产生成为宫颈癌治疗的难点,自噬在宫颈癌中的研究为宫颈癌的治疗提供了新的方向[7]。而STAT3与细胞自噬作为调控宫颈癌细胞生理和病理重要通路,其两者之间的关系仍未阐明,本文将进一步探讨通过调控STAT3基因的活性,观察其对宫颈癌HeLa细胞的增殖、凋亡及自噬的影响,从而阐明STAT3与自噬之间的调控机制及其对宫颈癌HeLa细胞增殖与凋亡的作用,将为宫颈癌尤其是晚期及复发性宫颈癌的治疗与预后提供新思路。
1 材料与方法
1.1 主要材料宫 颈癌细胞株HeLa由广州军区广州总医院医学实验科保存。STAT3过表达质粒、STAT3低表达质粒及空载体质粒均由广州复能基因生物公司合成,脂质体Lipfecta mineTM2000(Invitrogen),胎牛血清(Gibco),DMEM/F12培养基(Gibco),OPTI⁃MEM 培 养 基(Invitrogen),DMSO(Sigma),CCK⁃8(日本同仁),双抗(Hyclone),0.25%胰酶(不含EDTA 和酚红)(Gibco),0.25%胰酶(含EDTA 和 酚红)(Gibco),PBS缓冲液(吉诺),Annexin V/PI细胞凋亡检测试剂盒(日本同仁),MiniBEST Universal RNA Extraction Kit(TaKaRa),PrimeScript RT Master Mix Kit(TaKaRa),SYBR Premix EX TaqTMⅡ(TaKaRa),兔抗人STAT3抗体、兔抗人Bcl⁃2抗体、兔抗人Caspase⁃3抗体、兔抗人Beclin1抗体、兔抗人LC3⁃Ⅱ抗体和抗兔抗IgG(Cell Signaling Tech⁃nology),倒置相差荧光显微镜(Olympus),流式细胞仪LSR Ⅱ(BD Bio⁃science),定量PCR仪(QIA⁃gen Rotor⁃Gene⁃6000 Sequence Detection System)。
1.2 细胞的培养和转染宫 颈癌HeLa细胞用含10%胎牛血清和1%双抗的DMEM/F12培养液培养,置于37℃5%CO2恒温培养箱中,当细胞处于对数生长期时用于实验。将STAT3过表达质粒、STAT3低表达质粒及空载体质粒分别利用脂质体介导瞬时转染至宫颈癌HeLa细胞中,具体方法按照Lipfecta mineTM2000转染试剂盒说明书进行操作。转染48 h后,于倒置相差荧光显微镜下进行观察和拍照,估算细胞的转染效率。细胞转染效率(%)=荧光场红色荧光细胞数/相同视野明场细胞总数×100%。
1.3 实时荧光定量反转录聚合酶链反应(qRT⁃PCR)技术检测STAT3mRNA的表达H eLa细胞转染48 h后,用MiniBEST Universal RNA Extraction Kit试剂盒分别提取各组细胞的总RNA。以紫外分光光度计A280、A260定量测定,使提取总RNA测量结果纯度A280/A260为1.8~2.0,浓度≤500 ng/μL为合格。质量合格的RNA逆转录成cDNA,严格按照试剂盒说明书进行,反应体系为25μL。反应条件:预变性95 ℃ 5 min,94 ℃ 3 0 s,65 ℃ 3 0 s,重复40个循环,融解曲线分析:温度60~95℃。采用2⁃ΔΔCT法对数据进行相对定量分析,每个样品每次含有2个复孔,结果来自3次独立重复的实验。
1.4 CCK⁃8实验检测细胞增殖用 含10%FBS的DMEM/F12完全培养液重悬转染48 h后的HeLa细胞,调整细胞浓度为1×104/mL,取各组细胞悬液按100μL/孔接种于96孔板中培养。将培养板在37℃5%CO2条件下的恒温培养箱中预培养24 h后,向每孔加10μL CCK⁃8溶液,移入培养箱中继续孵育,分别在不同的时间点(1、2.5、4 d)取出培养板,用全自动酶标仪测定每孔的吸光度(OD)值,测定波长450 nm,并绘制细胞增殖曲线。每个样品每次含有3个复孔,结果来自3次独立重复的实验。
1.5 流式细胞仪检测细胞凋亡率转 染结束后将各组细胞置于37℃5%CO2恒温培养箱中继续培养48 h,用不含EDTA的胰酶消化细胞后,置于离心机中以1 000 r/min 4℃离心5 min收集细胞。严格按照Annexin V/PI细胞凋亡检测试剂盒的说明书进行避光染色,1 h内通过流式细胞仪检测细胞的凋亡情况。细胞总凋亡数=早期凋亡细胞数(Annexin V+/PI-)+晚期凋亡细胞数(Annexin V+/PI+)。
1.6 电镜观察各组细胞的凋亡小体及自噬小体转染结束后将各组细胞置于37℃5%CO2恒温孵箱中继续培养48 h,取生长状态良好的细胞用0.25%胰酶消化后,移入1.5 mL的EP管后置于离心机中以1 000 r/min离心5 min后弃上清液,收集足够量的细胞并立即加入适量的3.1%戊二醛进行固定,随后进行缓冲液漂洗、脱水剂梯度脱水、丙酮置换、包埋剂梯度浸透、加热聚合、莱卡EM UC 7超薄切片仪修快、切片(50~70 nm)、醋酸双氧铀50%乙醇饱和溶液100μL染色(15 min~1 h)、双蒸水冲洗、柠檬酸铅100μL染色15 min,日立H⁃7650透射电镜观察各组细胞的凋亡小体及细胞形态。
1.7 蛋白裂解及Westernblot法检测凋亡相关蛋白 B cl⁃2 和 C aspase⁃3 及 自 噬 相 关蛋白 B eclin1 和LC3⁃Ⅱ的表达收 集细胞,加入RIPA蛋白裂解液冰上裂解,使用BCA试剂盒测量蛋白浓度,进行SDS⁃PAGE电泳分离,将分离的蛋白转移至PVDF膜上,置于封闭液中在室温下于摇床上封闭2 h,加入相应的一抗,4℃孵育过夜,使用TBST润洗三次,加入带有荧光标记的二抗室温下避光孵育1 h,使用TBST洗膜三次,使用Odyssey®扫描仪进行扫膜,运用Image Studio软件分析结果。
1.8 统计学方法以 上所有的实验均重复3次,数据结果用均数±标准误表示。采用Graphpad Prism 5.0软件作图。多组间比较使用one⁃way ANOVA bonfferoni correction,post hoc unpaired t stu⁃dent test,P<0.05为差异有统计学意义。所有统计学检验均使用SPSS22.0进行分析。
2 结果
2.1 瞬时转染效率利用LipfectamineTM2000将STAT3过表达质粒、STAT3低表达质粒及空载体质粒分别瞬时转染至宫颈癌HeLa细胞中,48 h后,先在倒置相差荧光显微镜的明场下观察细胞,后在同一视野下转换成荧光场,可观察各组带红色荧光的细胞数均大于各组细胞总数的90%,提示转染效率均超过90%(图1)。为证实各组转染后HeLa细胞中STAT3的表达水平,在各组细胞转染48 h后,进一步采用qRT⁃PCR进行检测,结果显示STAT3过表达质粒组STAT3的表达量(7.35±0.56),较其空载体质粒组STAT3的表达量(1.00±0.08)明显上调,差异具有统计学意义(P<0.01);STAT3低表达质粒组STAT3的表达量(0.37±0.05),较其空载体质粒组STAT3的表达量(1.00±0.08)明显下调,差异具有统计学意义(P<0.01)(图1)。

图1 转染后各组细胞中红色荧光的表达及qRT⁃PCR检测转染后各组细胞STAT3 mRNA水平Fig.1 The expression of red fluorescence and the expression of STAT3 mRNA detected by quantitative RT⁃PCRin each groups after transfection
2.2 CCK⁃8检测STAT3对宫颈癌HeLa细胞增殖能力的影响结果显示,转染STAT3过表达质粒48 h后HeLa细胞的增殖活性较其空载体质粒组显著增强(P<0.01);转染STAT3低表达质粒48 h后,则相反(P<0.01)。这提示STAT3能显著提高宫颈癌HeLa细胞的增殖能力(图2)。

图2 不同转染组的宫颈癌HeLa的增殖曲线Fig.2 Proliferation of cell growth curve of each groups after transfection
2.3 流式细胞术检测STAT3对宫颈癌HeLa细胞凋亡率的影响结果显示,将上述各组细胞转染48 h后,转染STAT3过表达质粒后细胞凋亡率为(8.16±1.25)%,相对于空载体质粒组(14.78±1.85)%,凋亡细胞数的比例显著下降(P<0.01);转染STAT3低表达质粒后细胞凋亡率为(23.08±0.87)%,相对于空载体质粒组(14.78±1.85)%,凋亡细胞数的比例显著升高(P<0.01)。这说明STAT3能显著抑制宫颈癌HeLa细胞的凋亡(图3)。

图3 STAT3对宫颈癌HeLa细胞凋亡的影响Fig.3 Effect of STAT3 on the apoptosis of cervical cancer HeLa cells
2.4 电镜观察各组细胞的凋亡小体及自噬小体各组细胞转染48 h后,结果显示:如箭头所示,STAT3低表达质粒组可见多个凋亡小体及自噬小体,STAT3过表达质粒组未见凋亡小体及自噬小体,对照空载体质粒组可见少量自噬小体。与对照空载体质粒组相比,P<0.05。
2.5 Westernblot法检测各组细胞的 B cl⁃2、Cas⁃pase⁃3、Beclin1和LC3⁃Ⅱ的表达各 组细胞转染48 h后,Western blot法检测结果显示:与空载体质粒组相比,过表达STAT3质粒组可明显降低HeLa细胞 C aspase⁃3、Beclin1 和 L C3⁃Ⅱ蛋白的表达,增加Bcl⁃2蛋白的表达(P<0.01);而低表达STAT3质粒组,则相反(P<0.01)。这表明STAT3促进HeLa细胞增殖可能与降低 C aspase⁃3、Beclin1和LC3⁃Ⅱ蛋白表达相关(图5)。
3 讨论
自噬是一种高度保守的细胞降解过程,在正常的生理条件下,自噬起到维持细胞内稳态和调节细胞器更新的作用;为了应对细胞的应激状态,自噬防止受损的蛋白质和细胞器积累,从而抑制致癌作用[8-12]。自噬在清除有害成分并维持细胞稳态以应对一系列细胞外攻击时可能会激活其中一个重要的应激反应途径的下游信号传导通路,即STAT3信号通路,该通路已涉及自噬过程的多个方面[13]。在宫颈癌发展的各个阶段中,STAT3与自噬之间的相互调控是一个复杂的过程,二者之间的不同调控组合对宫颈癌的发展和转归都发挥着重要作用。然而,对于二者之间的调控作用与具体机制,目前尚未得出定论。因此,进一步阐明STAT3与自噬之间的调控机制及其对宫颈癌HeLa细胞增殖与凋亡的影响,将为宫颈癌尤其是晚期及复发性宫颈癌的治疗与预后提供新思路。
本研究进一步证实STAT3在宫颈癌中通过调控细胞自噬发挥着类似促癌基因的作用。选取宫颈癌HeLa细胞作为研究对象,通过瞬时转染,上调或者下调内源性STAT3基因的表达;并通过CCK⁃8实验、流式细胞术和电镜检测STAT3对宫颈癌HeLa细胞增殖与凋亡的影响。实验结果显示,通过上调STAT3基因能显著地抑制宫颈癌HeLa细胞发生凋亡,而促进HeLa细胞的增殖能力;通过下调STAT3基因,则相反;表明STAT3类似一种促癌基因发挥作用,促进宫颈癌的进展,与之前的报道相符合[5-6]。
有研究表明,STAT3在肿瘤细胞的自噬中发挥着重要的调控作用[14],但STAT3在宫颈癌细胞中与自噬的关系及作用机制仍不清楚。大量报道指出Beclin1和LC3⁃Ⅱ是自噬过程中必不可少的蛋白,高表达的Beclin1能够减缓肿瘤细胞的生长,抑制细胞周期的进程[15];在哺乳动物细胞中,抗凋亡蛋白Bcl⁃2在非饥饿条件下与Beclin1结合并抑制其自噬功能[16]。因此,本研究使用Western blot法检测 Bcl⁃2、Caspase⁃3、Beclin1 和 LC3⁃Ⅱ的蛋白表达情况。结果显示,上调STAT3基因后,明显地抑制HeLa细胞的Caspase⁃3、Beclin1和LC3⁃Ⅱ的蛋白表达,促进Bcl⁃2的蛋白表达;而下调STAT3基因后,则相反。这提示STAT3对HeLa细胞的增殖和凋亡的影响可能与其调控细胞自噬进而影响Bcl⁃2、Caspase⁃3、Beclin1 和 LC3⁃Ⅱ的蛋白表达有关,但STAT3调控的细胞自噬的信号通路仍不清楚。有研究表明,其中AMPK信号通路是最重要的细胞自噬通路之一,其在刺激因子作用下磷酸化,抑制mTOR,导致LC3⁃Ⅱ表达增加,引起细胞自噬[17-18]。STAT3在宫颈癌中是否可以直接靶向AMPK,或者通过靶向相关自噬基因从而介导下游的信号通路来调控宫颈癌细胞的自噬,从而影响宫颈癌的发生发展,仍有待进一步深入研究。
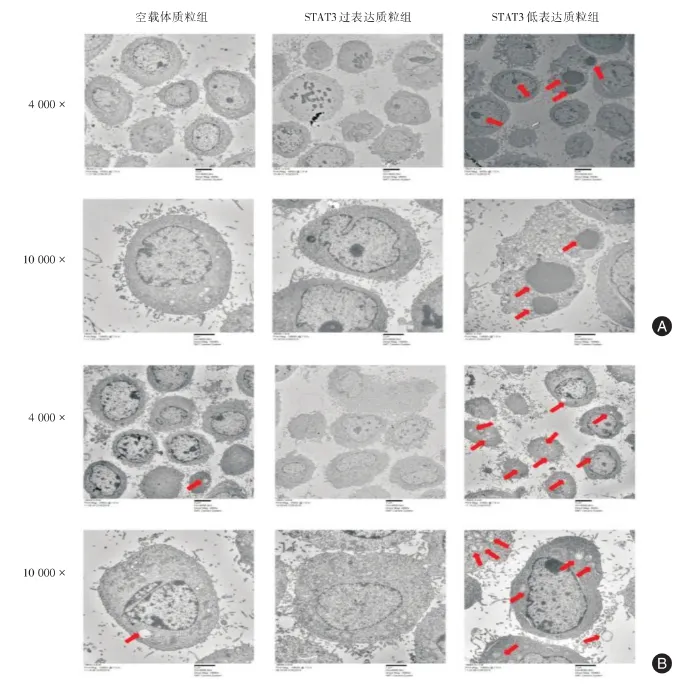
图4 透射电子显微镜观察各组细胞的凋亡小体及自噬小体Fig.4 The apoptotic bodies and autophagosomes of each groups were observed uesed by transmission electron microscope

图5 Western blot法检测各组HeLa细胞Bcl⁃2、Caspase⁃3、Beclin1和LC3⁃Ⅱ的蛋白表达Fig.5 The protein expressions of Bcl⁃2,Caspase⁃3,Beclin1 and LC3⁃Ⅱ in each group after transfection detected by Western blot analysis
综上所述,STAT3能促进人宫颈癌HeLa细胞增殖,抑制其自噬与凋亡,可能与其影响凋亡相关蛋白 Bcl⁃2和Caspase⁃3及自噬相关基因Beclin1 和LC3⁃Ⅱ的蛋白表达有关,但其具体机制仍有待进一步研究。本课题组后续将进一步研究STAT3的下游靶基因和可能作用的信号通路,评估STAT3能否成为针对宫颈癌细胞自噬的基因治疗的重要靶点,为晚期及复发性宫颈癌的治疗提供新的思路。
