六倍体小黑麦重要性状的改良潜力探究
2018-08-28崔紫霞张传量许小宛孙道杰
崔紫霞,张传量,冯 洁,许小宛,孙道杰
(西北农林科技大学农学院,陕西杨凌 712100)
近年来,小黑麦以强大的生物量以及优良的营养品质成为值得倡导的饲料作物[1-2]。小黑麦(Triticale)是由小麦属(Triticum)和黑麦属(Secale)物种经属间有性杂交和杂种染色体数加倍后人工合成的新物种,不仅拥有小麦高产和优质的特点,还具有生长茂盛、茎叶多汁、含糖量高、综合抗性强、分蘖力强、大穗大粒、籽粒中蛋白质及赖氨酸含量高等优良性状[3-5]。小黑麦在生产上应用最多的是八倍体和六倍体。六倍体(2n=6X=42,AABBRR)小黑麦是四倍体小麦和二倍体小黑麦杂交后再经过染色体加倍形成的异源六倍体。我国生产中应用的六倍体小黑麦分为粮用型及粮饲兼用型[4]。前者一般籽粒产量较高、株高与小麦相近,后者则株高较高,生物产量大。
苗相是鉴定小黑麦材料为冬性或春性的标准,越匍匐说明该品种冬性越强,但冬性越强并不能说明抗寒性越强。研究表明,春化和抗寒性由不同基因控制,冬性强的品种其春化温度低、时间长,低温春化过程也是幼苗抗寒性形成的过程,有利于增强幼苗的抗寒性[6-8]。
株高、穗长、穗颈长、穗下节长和倒二节长属于株型性状,单株穗数、小穗数、穗粒数和千粒重属于产量相关性状[9-10]。何贤芳等[9]研究表明,产量与旗叶宽和穗长显著或极显著正相关;吴同彦等[11]研究发现,穗长与千粒重呈不显著负相关,穗下节长与穗粒数呈正相关,倒二节长与旗叶宽呈显著正相关。海 燕等[12]利用小麦花药培养技术构建的DH群体对株高、单株穗数、穗长、穗下节长等性状进行了遗传研究,发现被测性状均有较高的遗传力。王瑞清等[13]用6个小黑麦品种按完全双列杂交方法配置杂交组合,对亲本、F1、F2的产量性状进行了遗传分析,发现提高穗粒数和千粒重可以有效增加产量。胡立芹等[14]分析了从CIMMYT引进的111份六倍体小黑麦材料的遗传多样性特点,发现农艺性状和品质性状均表现出较大的差异,具有丰富的多样性。
利用冬、春性小黑麦杂交培育广适性小黑麦的研究开始于20世纪70年代,杂交所产生的遗传变异能使春化和光照发育特性产生一系列变异,不仅表现在籽粒灌浆期、灌浆速度、成熟期的变异上,还表现在抗病性、抗逆性、品质及产量性状上[15]。艾尼瓦尔等[16]认为,在新疆选育与推广冬、春性杂交小黑麦品种能与新疆复杂的生态条件以及耕作制度相适应。马翎健等[17]曾对不同冬春性类型小麦进行组配,发现“冬麦×春麦”杂交组合的F1代穗分化优势、产量优势明显优于其他类型组合。曹新有等[18]研究发现,冬小麦在其他特性相当的情况下,抗寒性强的偏春性品种表现出更广泛的适应性,可见冬、春麦杂交可以作为提高麦类作物广适性的重要育种途径。
本研究以小黑麦冬性与春性、春性与春性杂交的F1、F2群体为材料,进行六倍体小黑麦性状的遗传分析及改良潜力探究,为粮饲兼用型小黑麦育种提供借鉴。
1 材料与方法
1.1 供试材料以及田间种植
供试材料为冬性小黑麦兰小黑(LXH)、春性小黑麦CM-12和CM-13及其杂交构建的CM-13/CM-12、CM-12/LXH和CM-13/LXH春/春型、冬/春型的F1和F2群体,试验材料LXH由本试验室保存,CM-12和CM-13由国际玉米小麦改良中心(CIMMYT)提供,3个材料均为粮饲兼用型小黑麦。3个群体的亲本以及F1代和F2代分别于2015年10月-2016年6月和2016年10月-2017年6月种植于西北农林科技大学试验田,群体小区行长2 m,行距25 cm,株距为15 cm,常规管理,生长期间未发生严重病虫害。
1.2 农艺性状的测定
小黑麦性状调查参照《小麦种植资源描述规范和数据标准》。在越冬前苗期调查苗相,苗相作为冬春性研究的一个鉴定标志,分为直立、半匍匐、匍匐三种,春性材料为3级直立,冬性材料为1级匍匐。两个春性亲本均为3级直立,冬性材料LXH为1级匍匐。按单株分别记录。
在小黑麦乳熟期,3个亲本以及3个F1代每个供试材料选取10株调查株高、单株穗数、穗长、小穗数、穗粒数、穗颈长、旗叶长与宽、穗下节长和倒二节长;收获后测定千粒重。3个F2群体每个群体随机调查200株用于表型及遗传分析。
1.3 数据处理
采用Microsoft Excel软件计算配合力、平均数、变异系数、偏度以及峰度值,用IBM SPSS Statistics 22软件计算3个小黑麦F2群体的9个性状的相关系数和遗传力。
遗传力计算公式:
H2=VG/Vp×100%=VG/(VG+VE)×100%
式中,VP代表表型方差,VG代表遗传方差,VE代表环境方差,H2代表遗传力。
2 结果与分析
2.1 亲本各个性状的配合力分析
苗相是鉴定麦类作物冬、春性的主要依据,苗相一般配合力较小的品种在田间表现出较强的抗冻性。由表1、表2可以看出,LXH苗相的一般配合力最低,以LXH为亲本的两个组合苗相的特殊配合力均低于春/春型小黑麦组合,最低的是CM-12/LXH,说明冬/春型杂交有利于小黑麦的抗寒性改良,印证了前人的结论[18]。
小麦穗颈长与收获指数及单株产量密切相关,在一定株高条件下,穗下节长度在30 cm左右时对产量有一定促进作用[19]。株高越高,越容易倒伏,不利于高产;但株高过低会减少小黑麦的生物量。亲本LXH的株高、穗颈长、穗下节长和倒二节长的一般配合力均最高,其次是CM-13,最后是CM-12,但后两者相差不大。株高、穗长、穗颈长、穗下节长和倒二节长特殊配合力最大的是CM-13/LXH组合,其次是CM-12/LXH组合,最小是CM-12/CM-13组合。关于穗长,亲本CM-12的一般配合力最大,CM-12/LXH组合的特殊配合力最大。
就小穗数、旗叶长和旗叶宽而言,LXH的一般配合力最大,CM-13/LXH组合的特殊配合力最大;关于单株穗数,LXH的一般配合力最大,CM-12/LXH组合的特殊配合力最大;关于穗粒数,CM-13的一般配合力最大,CM-13/LXH的特殊配合力最大。综合考虑,作为粮饲兼用型小黑麦最优的组合为CM-12/LXH,其次是CM-13/LXH,然后是CM-12/CM-13。
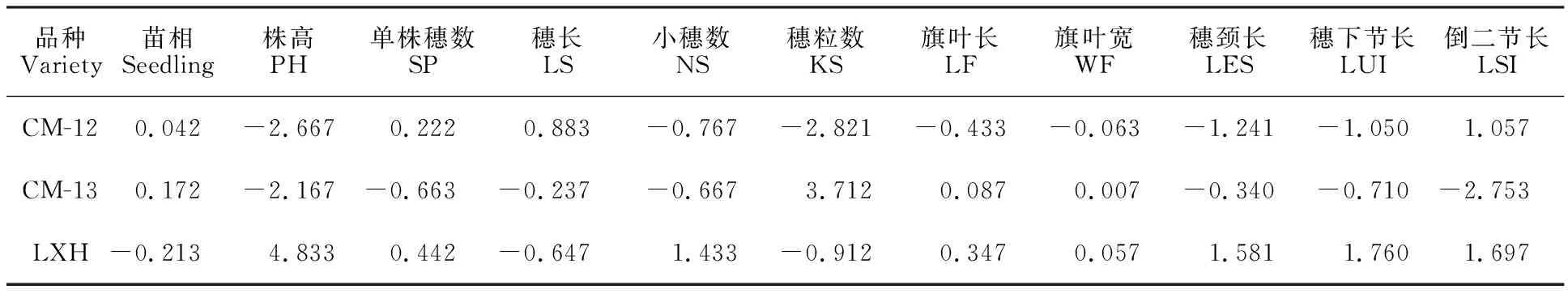
表1 3个亲本各性状的一般配合力Table 1 General combining ability of traits of three parents

表2 3个组合的特殊配合力Table 2 Special combining ability of three combinations
由表3可以看出,在CM-12/LXH的F2群体中,株高与穗长、小穗数、旗叶长、旗叶宽、穗颈长、穗下节长、倒二节长呈极显著正相关;穗长与小穗数、旗叶长、旗叶宽呈极显著正相关,与穗下节长呈显著正相关;单株穗数与穗颈长、穗下节长、倒二节长呈不显著负相关;穗长与穗颈长呈不显著负相关;小穗数与旗叶长、旗叶宽呈极显著正相关,与穗下节长呈显著正相关;旗叶长与旗叶宽、穗下节长呈极显著正相关;旗叶宽与穗下节长呈极显著正相关,与穗颈长呈显著正相关;穗颈长与穗下节长、倒二节长呈极显著正相关;穗下节长与倒二节长呈极显著正相关。
在CM-13/LXH的F2群体中,株高与穗长、小穗数、穗颈长、穗下节长以及倒二节长呈极显著正相关;单株穗数与穗长、旗叶宽、穗颈长、倒二节长呈极显著正相关,与旗叶长呈显著正相关,与倒二节长呈不显著负相关;穗长与小穗数、旗叶长、旗叶宽、穗颈长、穗下节长呈极显著正相关;小穗数与旗叶宽、穗下节长呈极显著正相关,与旗叶长呈显著正相关;旗叶长与旗叶宽、穗下节长呈极显著正相关,与穗颈长呈显著正相关,与倒二节长呈不显著负相关;旗叶宽与穗颈长、穗下节长呈极显著正相关,与倒二节长呈不显著负相关;穗颈长与穗下节长呈极显著正相关。
在CM-12/CM-13的F2群体中,株高与穗长、旗叶长、倒二节长呈极显著正相关,与旗叶宽、穗下节长呈显著正相关,与穗颈长呈不显著负相关;单株穗数与穗长、旗叶长、旗叶宽呈极显著正相关,与小穗数、穗下节长呈显著正相关,与穗颈长呈不显著负相关;穗长与小穗数、旗叶长、旗叶宽、穗下节长、倒二节长呈极显著正相关,与穗颈长呈不显著负相关;小穗数与旗叶宽、穗下节长呈极显著正相关,与穗颈长呈显著正相关;旗叶长与旗叶宽、穗下节长呈极显著正相关,与倒二节长呈显著正相关,与穗颈长呈不显著负相关;旗叶宽与穗下节长呈极显著正相关;穗颈长与穗下节长呈极显著正相关,与倒二节长呈不显著负相关。
在CM-12/LXH的F2群体中,株高、穗长、小穗数、旗叶长、旗叶宽、穗颈长、穗下节长、倒二节长之间相关性高,有些达到极显著水平,小穗数、旗叶长、旗叶宽和穗颈长都能影响产量,说明在提高生物量性状的同时,产量相关性状也会提高,但是在CM-12/LXH的F2群体中,单株穗数跟茎秆特性呈显著负相关或相关性较弱,所以在提高生物量性状的同时要考虑单株穗数,使粮饲兼用型小黑麦达到最优性状组合,从而达到高产目标。而其他两个群体的产量相关性状跟株型性状之间的相关性较CM-12/LXHF2群体弱,在CM-13/LXH的F2群体中,单株穗数、穗长、小穗数跟其他性状之间都具有正相关关系,相关程度不同,因此,在以收获籽粒为目的的小黑麦育种中,CM-13/LXH优于其他两个群体。
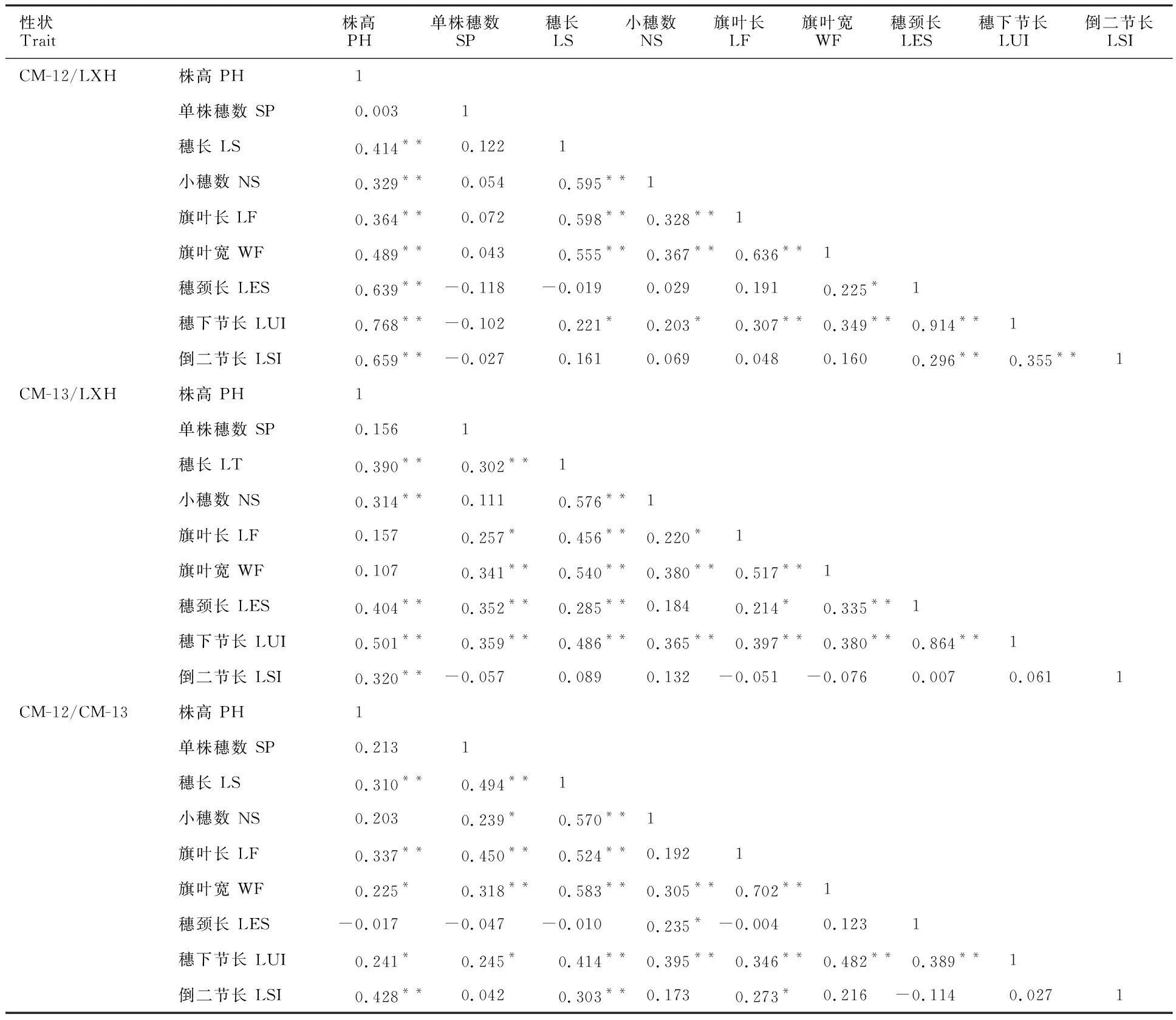
表3 3个群体各个性状的相关系数Table 3 Correlation coefficients of each trait in the three populations
2.2 各性状的变异系数及遗传力比较
由表4可以看出,3个群体中,株高的变异系数为6.08%~11.78%,单株穗数的变异系数为34.30%~46.57%,穗长的变异系数为9.78%~12.53%,小穗数的变异系数为8.67%~11.73%,旗叶长的变异系数为14.51%~17.81%,旗叶宽的变异系数为10.65%~16.56%,穗颈长的变异系数为13.31%~23.04%,穗下节长的变异系数为10.26%~15.38%,倒二节长的变异系数为12.61%~14.01%。9个性状变异程度表现为单株穗数>穗颈长>旗叶长>倒二节长>旗叶宽>穗下节长>穗长>小穗数>株高。CM-12/LXH的9个性状变异系数均高于其他两个群体,说明CM-12/LXH组合后代性状分离范围广,变异类型多,多样性丰富,进行新品种选育的潜力较大。

表4 F2群体与亲本的各性状值Table 4 Traits of F2 population and their parents
3个群体株高的遗传力为0.674~0.793,单株穗数的遗传力为0.352~0.443,穗长的遗传力为0.699~0.925,小穗数的遗传力为0.094~0.794,旗叶长的遗传力为0.626~0.761,旗叶宽的遗传力为0.738~0.820,穗颈长的遗传力为0.735~0.890,穗下节长的遗传力为0.477~0.888,倒二节长的遗传力为0.709~0.786。9个性状的遗传力大小为穗颈长>穗长>旗叶宽>株高>倒二节长>穗下节长>旗叶长>小穗数>单株穗数。
在CM-12/LXH的F2群体中,遗传力最小的是单株穗数(0.443),遗传力最大的是穗长(0.925),其他性状的遗传力普遍高于其他两个群体,说明CM-12/LXH的F2群体中各个性状的遗传力受环境影响较小。在CM-13/LXH的F2群体中,遗传力最小的是小穗数(0.094),遗传力最大的是穗颈长(0.890)。在CM-12/CM-13的F2群体中,倒二节长(0.786)的遗传力最大,而单株穗数(0.378)的遗传力最小。
分析3个群体9个性状正态分布的偏度和峰度,推测在CM-13/LXH的F2群体中,株高可能存在微效基因间的互作;穗长、小穗数和穗颈长可能存在微效基因的重叠作用;旗叶宽、穗下节长和倒二节长可能存在微效基因间的互补作用。在CM-12/CM-13的F2群体中,穗长、小穗数、旗叶宽和穗颈长可能存在微效基因间互补作用。其他性状的微效基因相互作用不明显。
3 讨 论
在小黑麦品种的选育中,正确选择亲本材料进行杂交组合配制是筛选优良品种的关键。亲本的配合力是决定优良性状传递稳定性的主要因素[20]。配合力受多种基因效应支配,一般配合力由基因的加性效应决定,是可以遗传的性状,一般配合力的高低由亲本所含的有利基因位点的多少决定;特殊配合力由基因的非加性效应决定,受基因间的显性、超显性和上位性效应控制,只能在特定的组合中由双亲的等位基因或非等位基因间的互作反映出来,是不能遗传的性状[21-22]。本研究发现,在3个亲本中,不论株型性状还是产量性状,LXH是最优亲本;3个F2群体比较,CM-12/LXH的配合力最好,其株型性状和产量性状的相关系数、变异系数、遗传力最高,具有较好应用潜力,可以用于粮饲兼用小黑麦的改良。CM-13/LXH的F2群体可以用于粮用小黑麦后期研究。本研究中,9个被测性状变异系数表现为单株穗数>穗颈长>旗叶长>倒二节长>旗叶宽>穗下节长>穗长>小穗数>株高,9个性状的遗传力表现为穗颈长>穗长>旗叶宽>株高>倒二节长>穗下节长>旗叶长>小穗数>单株穗数。
单株穗数一直是育种家关注的麦类作物产量构成的重要因素[23],冯春叶等[24]认为,拔节前小麦主茎和分蘖对营养物质的竞争会影响最终成穗数;赵广才等[25]提出了优势蘖组概念;封超年等[26]认为,茎蘖成穗率与产量线性正向相关。虽然单株穗数对于麦类作物很重要,但对于小黑麦的分蘖性状研究的较少,程治军等[27]用八倍体小黑麦研究了 Rht12对小黑麦株高、单株分蘖数和种植饱满度的影响发现, Rht12的导入提高了八倍体小黑麦的分蘖力。本研究中,单株穗数的变异系数高达46.57%,这对于粮饲兼用型小黑麦的单株穗数来说育种潜力较大,具体生理生化机制还需进一步研究。
旗叶是麦类植物籽粒碳水化合物的主要合成源之一,而旗叶长和旗叶宽直接影响光合速率。研究表明,旗叶性状和穗粒重、千粒重、穗粒数和小麦产量间存在显著正相关关系[28]。但是旗叶并不是越长越好,高产小麦株型一般要求叶片上挺、旗叶宽,跟茎秆的夹角小,而旗叶的长度对产量影响不明显[29-31]。田间观察发现,CM-12/LXH的F2群体和CM-13/LXH的F2群体的旗叶普遍上挺,说明这两个群体的旗叶符合高产株型对叶片的要求,并且遗传力均较高,可以用于高产小黑麦品种的选育。
