绿豆不同品种对绿豆象的抗性初探
2018-08-10王宏民成小芳樊艳平郑海霞张耀文张仙红
王宏民 成小芳 樊艳平 郑海霞 张耀文 张仙红,*
绿豆不同品种对绿豆象的抗性初探
王宏民1,**成小芳2,**樊艳平2郑海霞2张耀文3张仙红2,*
1山西农业大学经济管理学院, 山西太谷 030801;2山西农业大学农学院, 山西太谷 030801;3山西省农业科学院作物研究所, 山西太原 030031
为明确抗虫绿豆抗绿豆象的有效成分, 采用室内人工接虫方法, 进行了13个不同绿豆品种(品系)对绿豆象的抗虫性鉴定, 并对筛选获得的抗豆象品种的抗虫成分进行了研究。结果表明, 绿豆象对B18、B20、B23、B27、A22和晋绿7号绿豆的为害率均低于10%, 为高抗型绿豆; 其余7个绿豆品种受害率均在90%以上, 属高感型绿豆。绿豆象卵的孵化率在抗、感绿豆品种间无显著性差异, 而绿豆象发育历期、雌、雄成虫体重及成虫羽化率在抗、感绿豆品种间差异显著, 同一品种去皮绿豆与带皮绿豆相比, 绿豆象卵孵化率、成虫羽化率及种子受害率等指标均无显著性差异。抗虫成分试验表明, 当绿豆象取食添加抗虫绿豆蛋白25%和50%的人工(合成)绿豆后, 其成虫羽化率由30.48%降低到0, 但不随淀粉比例的增加而变化。可见, 抗虫绿豆抗绿豆象的主要成分为其种子中的蛋白质。
绿豆; 不同品种; 绿豆象; 抗性
绿豆象[(L.)]是绿豆、红豆、豇豆等豆科作物生长期及籽粒储藏期的主要害虫, 对产量和品质有较大影响。一个生活周期内, 绿豆象可使绿豆产量受损30%~56%[1], 若成虫羽化引起2次侵染, 危害更加严重, 危害率可达100%[2]。采用温度、湿度、微波处理等物理方法防治绿豆象, 受仪器限制和影响种子发芽而难以推广应用, 化学药剂防治又对绿豆的安全性有一定的影响。
利用植物品种的抗虫性来防治害虫是最经济、有效、安全的措施。早在20世纪40年代, 国外利用抗黑森瘿蚊[(Say)]的小麦品种有效地控制了该虫对小麦的危害; 我国利用抗白背飞虱()、褐飞虱()的水稻品种也大大减轻了这2种害虫对水稻的危害。Kitamura等[3]以抗豆象绿豆野生种TC1966与感豆象栽培种Osaka-ryodutou和No.3-ryokutou进行人工杂交试验, 成功培育出抗豆象绿豆栽培种。之后, 中国、日本、澳大利亚等国家对抗豆象的种质创新进行了研究, 以TC1966为抗源分别与VC1973A、VC2778A、VC2768A等进行杂交并培育出一批农艺性状优良的抗豆象材料[4]。
基于作物利用自身的抗虫性来降低害虫为害的许多优点, 本研究采用人工接虫方法, 筛选出抗绿豆象的绿豆品种, 并从抗虫品种中提取总蛋白和淀粉, 测定了绿豆象取食抗、感绿豆及人工(合成)绿豆后成虫体重、羽化率及种子受害率, 探讨这些因素与抗虫性的关系, 进而为明确绿豆抗绿豆象的生理生化机制及寻求有效杀虫成分提供依据。
1 材料与方法
1.1 材料
1.1.1 供试绿豆 试验绿豆A22、B18、B20、B23、B24、B27、晋绿7号、潍绿2117、宝绿2号、冀绿7号、串辐1号、晋绿1号、VC2768B共13个品种均由山西省农业科学院作物研究所提供, 分别挑选健康豆粒于-20℃冷冻备用。
1.1.2 供试虫源 将实验室饲养的绿豆象成虫雌雄配对后, 释放于装有潍绿2117绿豆的塑料袋中使其自由产卵7 d后, 弃去成虫。着卵绿豆置于(27±1)℃、相对湿度(60±5)%的培养箱中进行饲养繁殖。
1.1.3 主要仪器 Y9600型电子分析天平(上海精密科学仪器有限公司); HPS-250恒温培养箱(哈尔滨东连电子技术开发有限公司); TDL5M台式大容量冷冻离心机(长沙湘仪离心机仪器有限公司); SZX16-3111体视显微镜(奥林巴斯, 日本)。
1.2 方法
1.2.1 不同品种绿豆对绿豆象的抗性测定 抗虫鉴定采用人工室内接虫方法[5], 将以上13个绿豆品种提前从冰箱取出, 各取50粒称重后, 分别放入自制无盖小纸杯(直径5 cm、高l cm)中, 随机放入带盖的大纸盒内(30 cm×20 cm×10 cm), 将室内饲养羽化1~3 d的绿豆象雌雄成虫100对接于纸盒内, 让其在盒内各品种绿豆上随机产卵, 5次重复。产卵5 d后, 除去所有成虫, 培养10 d后统计每份材料的着卵量和孵化率。第21天开始观察是否有成虫出现, 每12 h观察一次, 记录羽化时间及数量, 且收集从各品种羽化出的雌雄成虫并分别用电子天平称重; 到30 d后观察结束, 统计其羽化率(rate of adult emergence, RAE)和种子受害率(seeds dam rate, SDR), 同时, 剥开绿豆检测是否有死亡幼虫或因发育滞后而未羽化的成虫。绿豆种子对绿豆象抗性等级评价采用国家科技攻关项目选用的抗性评价分级标准, 共分为5个等级: 被危害率在0.1%~10.0%、10.1%~35.0%、35.1%~65.0%、65.1%~90.0%和>90.0%, 分别属高抗、抗、中抗、感及高感型品种。
1.2.2 抗、感去皮绿豆对绿豆象的抗性测定 晋绿1号、潍绿2117、晋绿7号和B20绿豆品种健康种子各取300粒, 分别用4℃蒸馏水浸泡12 h, 去皮后在50℃干燥24 h, 将干燥种子置于湿度为12%~14%的HPS-250恒温培养箱, 48 h后取出用于试验。选取完整无损的各处理绿豆种子每组50粒, 抗虫试验方法同上, 5次重复。
1.2.3 人工绿豆抗虫成分的试验 晋绿7号与潍绿2117绿豆种子分别磨粉, 过80目筛。将晋绿7号绿豆粉每100 g溶于1000 mL去离子水, 室温静置2 h后, 上清液和沉淀分别用于蛋白提取[6]和淀粉提取[7]。人工种子的制备参照Somta等[8]方法, 感虫绿豆潍绿2117豆粉与从晋绿7号提取到的蛋白和淀粉分别按25%、50%的比例混合, 与水按1.0∶0.5的比例混合后, 做成球状, 40℃烘干后将其含水量调为12%~14%, 每粒种子约重0.30 g, 10粒种子为1组, 5次重复。接虫方法同1.2.1, 统计着卵量、成虫羽化率及种子受害率。
1.3 数据统计分析
利用Microsoft Excel 2007统计和处理数据,采用SPSS19.0统计软件进行单因素方差分析(one-way ANOVA), Dunnett’s (<0.05)进行多重比较。
2 结果与分析
2.1 绿豆不同品种对绿豆象的抗性反应
由表1可知, 13个参试品种中, 绿豆种子的大小有显著性差异, 宝绿2号最大, B24最小, 每50粒种子分别重3.90 g和2.88 g, 而晋绿7号和潍绿2117分别重3.29 g和3.07 g, 但着卵量与种子的大小有关, 种子越大着卵量越多; 所有品种对绿豆象成虫产卵都有吸引力, 各品种间卵孵化率没有显著差异, 说明卵的孵化与绿豆的品种无关; B18、B20、B23、B27绿豆完好无损, 种子受害率为0, A22和晋绿7号种子受害率分别为3.60%和8.80%, 虽有被为害, 但受害率在0.1%~10.0%之间, 故这6个品种为高抗(HR)型绿豆。A22和晋绿7号绿豆内只有少量幼虫羽化为成虫, 成虫羽化率分别为1.39%和8.79%; 与取食高感型绿豆后的绿豆象相比, 取食高抗品种A22和晋绿7号的绿豆象发育历期延长1.0~1.5 d, 分别为25.84 d和25.46 d; 雌雄成虫体重均较轻, 雌成虫分别为5.01 mg和5.04 mg, 雄成虫分别为3.43 mg和3.71 mg。其余7个品种受害率均在90%以上, 属高感型(HS)绿豆, 取食高感型品种绿豆的成虫羽化率达59.9%~63.3%; 绿豆象发育历期较短, 雌雄成虫较重; 与取食高抗型绿豆后的绿豆象相比, 二者存在显著性差异(<0.05)。表明抗虫绿豆的抗虫性在于抗生性, 抗虫绿豆种子中含对绿豆象幼虫具有毒性作用的化学成分。

2.2 去皮抗、感绿豆不同品种对绿豆象的抗性
由表2可知, 感、抗绿豆去皮处理后, 与相应的带皮绿豆从着卵量、孵化率、成虫羽化率、发育历期及种子受害率相比, 均无显著性差异。表明绿豆皮不含有抗虫成分, 绿豆皮的有无对后续的实验并无影响, 故后续的实验所用绿豆均为带皮绿豆。
2.3 人工绿豆对绿豆象的抗性
由表3可知, 人工绿豆种子间着卵量无显著性差异, 表明绿豆种子中蛋白和淀粉含量的多少对绿豆象的产卵选择无影响。绿豆象幼虫取食含晋绿7号粗蛋白不同比例的人工绿豆后, 成虫羽化率有显著性差异, 随蛋白含量的增加成虫羽化率减小, 当感虫绿豆粉中加入25%的蛋白时, 成虫羽化率下降到30.48%; 含量达到50%时无绿豆象成虫羽化。但淀粉含量的变化对绿豆象成虫羽化率没有显著性影响。表明抗虫绿豆的抗绿豆象成分主要在绿豆种子的蛋白质部分。

表2 去皮抗、感绿豆抗虫性比较
不同小写字母表示在0.05水平差异显著。“NS”表示同列无显著性差异。
The different small letters in the same line indicate the significant difference at the 0.05 probability level. “NS” indicate no significant difference in the same column.
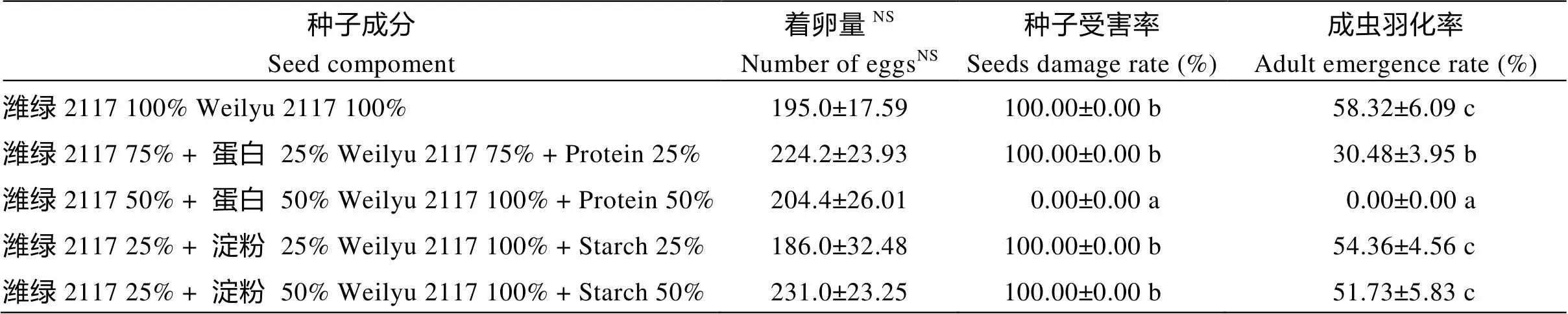
表3 不同比例抗虫绿豆蛋白和淀粉对绿豆象的影响
不同小写字母表示在0.05水平差异显著。“NS”表示同列无显著性差异。
The different small letters in the same line indicate the significant difference at the 0.05 probability level. “NS” indicate no significant difference in the same column.
3 讨论
植物的抗虫性表现为抗生性、耐害性和不选择性。本研究表明, 绿豆象在供试的13个绿豆品种上均可产卵, 表明绿豆象对不同绿豆品种之间没有表现出不选择性。因此, 抗虫绿豆对绿豆象的抗性表现为抗生性。
本试验表明, 抗虫绿豆对绿豆象的抗性与豆粒的大小没有关系, 产卵量的多少与种子的大小有关, 这与Lambrides等[10]报道的关于几种鞘翅目豆象对不同品种绿豆产卵特性的研究结果相一致。但与Johnson等[9]报道的绿豆的抗虫性与种子的大小和种皮有关的研究不同, 究其原因有待进一步探索。
种皮是绿豆象蛀蚀绿豆种子的第一道防御系统。据Janzen[11]报道, 在参试的73个物种中, 超过69%的种皮可阻止四纹豆象幼虫的入侵; 约有50%的绿豆象和四纹豆象幼虫不能穿透蚕豆种皮[12]; 菜豆象不能危害菜豆种子也与其种皮有直接的关系[13]; 此外, 研究还表明, 种皮也会使刚孵化的幼虫到达种仁的时间延长[14-16]。本研究表明, 当取食B18、B20、B23、B27、晋绿7号等抗虫品种时, 发现着卵部位下种仁部分受损, 并有死亡的低龄绿豆象幼虫, 说明绿豆象可以穿透种皮进入种仁。同时将相同品种的去皮绿豆和带皮绿豆进行绿豆象卵的孵化率、成虫羽化率、发育历期及种子受害率等试验比较, 结果显示这些指标在两者间均无显著性差异, 表明不同的抗、感绿豆品种的种皮既不对绿豆象胚胎的发育延长也不会降低卵的孵化率, 可见种皮对绿豆象的入侵没有影响, 抗绿豆象的抗虫成分在种仁而不在种皮, 绿豆种子自身的抗虫物质在害虫防御体系中起主导作用。这与Kashiwaba等[17]所报道的赤小豆种仁所含生化物质才是其抗虫的主要原因的研究结果一致。
据Talekar等[18]报道, 绿豆抗虫品种V2709和V2802中可能含有抗生物质; 赤小豆中也发现有抗豆象的化学成分[17]; Chen等[19]曾发现野生绿豆种TC1966中有一种对绿豆象幼虫有致死作用的富含半胱氨酸的蛋白。本研究表明, 随人工绿豆中抗虫绿豆蛋白比例的增加, 绿豆象幼虫的死亡率增加, 成虫羽化率下降, 说明抗虫绿豆的蛋白中含有影响绿豆象幼虫生长发育的抗虫成分, 但抗虫绿豆中到底是哪一种或哪几种蛋白质影响了绿豆象幼虫发育和成虫羽化, 还有待进一步研究。
4 结论
植物的不同品种对同种害虫的受害程度完全不同, 这就表现出品种的抗虫性。从13个参试品种中, 筛选出6个高抗绿豆象的绿豆品种B18、B20、B23、B27、A22和晋绿7号; 绿豆象产卵对抗、感绿豆品种无选择性差异; 取食抗虫绿豆的绿豆象幼虫、成虫体重均显著低于取食感虫绿豆, 且幼虫发育历期延长, 成虫羽化率降低; 绿豆种皮不含对绿豆象有毒杀作用的抗虫成分, 抗虫绿豆的抗虫成分存在于绿豆种子所含的蛋白质中。
[1] Gujar G T, Yadav T D. Feeding of(Fab.) and(Linn.) in green gram, 1978, 40: 108–112
[2] Tomooka N, Kashiwaba K, Vaughan D A, Ishimoto M, Egawa Y. The effectiveness of evaluating wild species: searching for sources of resistance to bruchid beetles in the genusCeratotropis., 2000, 115: 27–41
[3] Kitamura K, Ishmoto M, Sawa M. Inheritance of resistance to infestation with azukibean weevil inand successful in corporation to., 1988, 38: 459–464
[4] 程须珍, 王素华, 金达生, 杨又迪, 吴绍宇, 周吉红. 绿豆抗豆象遗传的初步研究. 植物遗传资源科学, 2001, 2(4): 12–15 Cheng X Z, Wang S H, Jin D S, Yang Y D, Wu S Y, Zhou J H. Preliminary study on heredity of mungbean resistance to bruchid., 2001, 2(4): 12–15 (in Chinese with English abstract)
[5] 程须珍, 王素华, 金达生, 王泮龙, 杨又迪. 绿豆抗豆象育种品系综合评价. 植物遗传资源学报, 2003, 4: 110–113Cheng X Z, Wang S H, Jin D S, Wang P L, Yang Y D. Evaluation on mungbean breeding lines for resistance to bruchid., 2003, 4: 110–113 (in Chinese with English abstract)
[6] 成珊, 沈群. 不同品种和不同产地绿豆分离蛋白功能特性的研究. 食品科技, 2009, 34(9): 148–153 Cheng S, Shen Q. Study on functional characteristics of mungbean protein isolates in different cultivars and from different areas., 2009, 34(9): 148–153 (in Chinese with English abstract)
[7] 李文浩, 谭斌, 刘宏, 张国权, 沈群. 我国9个品种绿豆淀粉的理化特性研究. 中国食品学报, 2013, 13(4): 58–64 Li W H, Tan B, Liu H, Zhang G Q, Shen Q. Physicochemical properties of starches separated from nine mung bean varieties grown in China., 2013, 13(4): 58–64 (in Chinese with English abstract)
[8] Somta P, Talekar N S, Srinives P. Characterization of(L.) resistance in(Thunb.) Ohwi and Ohashi., 2006, 42: 313–327
[9] Johnson C D. Coevolution of some seed beetles (Coleoptera: Bruchidae) and their hosts., 1974, 55: 1096–1103
[10] Lambrides C J, Imrie B C. Susceptibility of mungbean varieties to the bruchid species(F.),(Gyll.),(L.), and(Say.) (Coleoptera: Chrysomelidae)., 2000, 51: 85–89
[11] Janzen D H. How southern cowpea weevil larvae (Bruchidae:) die on non-host seeds., 1977, 58: 921–927
[12] Desroches P, Shazly E E, Mandon N, Duc G, Huignard JDevelopment of(L.) and(F.) (Coleoptera: Bruchidae) in seeds ofL. differing in their tannin, vicine and convicine contents., 1995, 31: 83–89
[13] Thiery D, Jarry M, Pouzat J. To penetrate or not to penetrate? A behavioral choice by bean beetle first-instar larvae in response toseed surface quality., 1994, 20: 1867–1875
[14] Oliveira A E A, Fernandes K V S, Souza A J, Santos P O. Influence of the soybean seed coat upon seed infestation and development oflarvae. In: Davies S, Evans G, Columbus F, eds. Soybean and Wheat Crops: Growth, Fertilization, and Yield. New YorkNova Science Publishers, 2009. pp 1–21
[15] Souza A J, Santos P O, Pinto M S T, Wermelinger T T, Ribeiro E S, Souza S C, Deus M F, Souza M C, Xavier-Filho J, Fernandes K V S, Oliveira A E A. Natural seed coats provide protection against penetration by(Coleoptera: Bruchidae) larvae., 2011, 30: 651–657
[16] Souza A J, Ferreira A T S, Perales J, Beghini D G, Fernandes K V S, Xavier-Filho J, Venancio T M, Oliveira A E A. Identification ofseed coat chitin-binding vicilins (7S globulins) with high toxicity to the larvae of the bruchid., 2012, 45: 118–124
[17] Kashiwaba K, Tomooka N, Kaga A, Han O K, Vaughan D ACharacterization of resistance to three bruchid species (spp., Coleoptera, Bruchidae) in cultivated rice bean ()., 2003, 96: 207–213
[18] Talekar N S, Lin C P. Characterization of(Coleoptera: Bruchidae) resistance in mungbean., 1992, 85: 1150–1153
[19] Chen K C, Lin C Y, Kuan C C, Sung H Y, Chen C S. A novel defensin encoded by a mungbean cDNA exhibits insecticidal activity against bruchid., 2002, 50: 7258–7263
Preliminary Study on Resistance of Different Mungbean Varieties to(L.)
WANG Hong-Min1,**, CHENG Xiao-Fang1,**, FAN Yan-Ping2, ZHENG Hai-Xia2, ZHANG Yao-Wen3, and ZHANG Xian-Hong2,*
1College of Economics and Management, Shanxi Agricultural University, Taigu 030801, Shanxi, China;2College of Agriculture, Shanxi Agricultural University, Taigu 030801, Shanxi, China;3Crop Sciences Institute, Shanxi Academy of Agricultural Sciences, Taiyuan 030031, Shanxi, China
To clarify the effective component in mungbean seeds against bruchid (L.), we identified the bruchid resistance of 13 mungbean varieties via in-house artificial feeding method, and the insect-resistant components in mungbean were investigated. Six mungbean varieties including B18, B20, B23, B27, A22, and Jinlyu 7, were found to be highly resistant to bruchid, showing seed-damage rates lower than 10%, whereas, the remaining seven varieties were highly susceptible with seed-damage rates higher than 90%. There was no significant difference in egg-hatch rate among all varieties. However, the developmental duration, the adult-emergence rate ,and the bodyweights of both male and female adults varied significantly between the resistant and susceptible varieties. Moreover, seed coat had no effect on the resistance to bruchid because samples with or without seed coat showed no significant difference in the rates of egg-hatch, adult emergence, and seed damage. The resistance test with synthetic mungbean indicated that the adult-emergence rate declined from 30.48% to 0 when the protein component of resistant mungbean varieties increased from 25% to 50%. In contrast, the increase of starch proportion had no effect on theadult emergence rate. Therefore, we conclude that seed proteins play an important role in bruchid resistance in mungbean.
mungbean; different varieties;(L.); resistance
2018-03-26;
2018-04-24.
10.3724/SP.J.1006.2018.01136
张仙红, E-mail: zxh6288@sina.com**同等贡献(Contributed equally to this work)
王宏民, E-mail: whm6970@163.com
2017-12-10;
本研究由国家现代农业产业技术体系建设专项(GARS-08-G11)资助。
This study was supported by the China Agriculture Research Systems (GARS-08-G11).
URL:http://kns.cnki.net/kcms/detail/11.1809.s.20180423.1446.002.html
