内陆盐沼芦苇根系形态及生物量分配对土壤盐分因子的响应
2018-08-09王继伟赵成章赵连春王小鹏
王继伟,赵成章,赵连春,王小鹏,李 群
西北师范大学地理与环境科学学院,甘肃省湿地资源保护与产业发展工程研究中心,兰州 730070
植物重要的生态策略和权衡维度,要通过植物内部与外部性状的精细协调来实现[1- 2],植物性状的变异格局以及性状间的“投资-收益”权衡则要靠植物自身的资源优化配置和补偿机制来调控[3]。根系是植物吸收水分、养分的重要器官,因直接与土壤环境接触,也是植物感受逆境胁迫的首要部位,更易受到土壤环境变化的影响。根系在逆境下的形态塑性和生物量分配格局是植物有效吸收和利用土壤养分最直接的适应特征之一,对植物的耐盐能力至关重要[4- 5]。根平均直径、根分叉数以及根冠比等性状体现了植物在空间上对土壤资源的占据和根的“觅食”能力,其变化影响着根系吸收代谢能力的发挥;比根长、根总长度及根深大小主要反映了根系拓展能力和利用土壤资源的集中程度,其变化影响着根系吸收效率的高低[6- 7]。有研究表明,盐渍化条件下,耐性植物通常会增加比根长、根冠比等主动拓展根系分布范围以适应逆境条件;而敏感植物则主要采取规避机制,通过减小根总长度、根深,增加根平均直径,来缩小根系分布范围、发展通气组织限制盐离子的过分吸收与缓解盐胁迫带来的缺氧损害[8- 10]。因此,它们都是反映根系吸收能力的可靠指标,对于揭示植物应对逆境胁迫的适应机制具有重要意义[11- 12]。内陆盐沼蒸发/降水比大,土壤盐渍化特点显著,土壤盐分等条件性因子往往呈梯度分布[13],制约着盐沼植物群落的空间组成与结构[14]。盐渍化环境可以改变土壤水分和离子的热力学平衡,通过影响植株有效吸收资源的数量,迫使植株地上、地下生物量分配格局发生改变,从而引起根系形态发生变化[8, 15- 16];植物能够感应外界胁迫,通过自身调节系统有效权衡比根长、根总长度、根分叉数、根平均直径和根冠比等重要根系性状的最佳收益/代价关系,构建与生境相匹配的根系吸收系统,以增强在胁迫条件下的生存机会。因此,从根系可塑性角度探究干旱区盐沼植被对不同土壤盐分环境的生态响应特征和规律,可为揭示盐沼植物环境适应策略具有重要意义。
近年来,国内外针对盐分胁迫下植物根系形态和分布[9, 17- 19]、根系活力[5, 20]、根系生理过程[21- 22]、根系结构和功能[23- 24]等方面已做了大量研究,然而,以往的研究多数为盐介质均匀的条件下(如水培、沙培)进行,仅考虑不同盐分梯度对植物根系的影响,忽略了自然界中植物常常经受多种非生物因素(如资源分布、环境条件等)的作用和影响,很难通过室内控制实验来准确判断植物根系的响应方式和适应特征。另外,以往的研究对象多集中于一些粮食作物和模式植物[25],而对广泛分布于干旱区盐沼湿地中的草本植物研究鲜见报道,此外,对长期遭受盐胁迫的草本植物是否存在生物量分配可塑性也不甚清楚。芦苇(Phragmitesaustralis)是一种典型的根茎型无性系禾本科植物,其地下部分具粗壮的匍匐根状茎,能够通过渗透调节作用适应盐渍化生境,并随背景条件的差异在异质生境中形成相适应的形态结构[26- 27]。目前,有关盐分胁迫下芦苇地上部分的生态响应特征[28]、适应机制[27]、生理生化[29]等方面已有一些报道。但是关于盐分胁迫下芦苇地下根系的适应机制,特别是不同盐分梯度下根系性状的变异规律和生物量分配策略尚不明晰。鉴于此,本文以秦王川国家湿地公园盐沼湿地的芦苇为研究对象,通过分析不同盐分梯度下生长芦苇的根系形态指标和生物量的变化,试图明晰(1)不同盐分梯度下芦苇根系性状和生物量分配模式如何变化?(2)芦苇的生态适应策略是什么?旨在为进一步认识湿地植物根系形态的环境可塑性机制提供理论依据。
1 材料和方法
1.1 研究区域与样地概况
研究区位于甘肃省兰州新区的秦王川国家湿地公园,地理位置介于103°35′38″—108°38′37″E,36°23′59″—36°27′56″N之间,东西宽约1.8 km,南北长约7.6 km,面积约274 hm2,海拔1895—1897 m。地势北高南低,属大陆性季风气候,年均气温6.9 ℃,年均降水量约285 mm,主要集中在6—9月,年日照时数为2700 h,无霜期为126 d,蒸发量高达1880 mm。由于地处乌鞘岭褶皱山岭南侧的边缘低山区,四周山地环绕,中心地带地势平坦开阔,形成盆地,盆地南部广泛分布第四系松散层孔隙潜水,含水层为砂碎石及中西砂层,受常年或季节性水淹的影响,形成以淡灰钙土为主的自然土壤,发育了水生和湿生植物群落。主要植物有:芦苇、水蒿(Artemisiaselengensis)、碱蓬(Suaedasalsa)、莎草(Cyperusrotundus)、珍珠猪毛菜(Salsolapasserina)、盐爪爪(Kalidiumfoliatum)、红砂(Reaumuriasongarica)等。
1.2 实验设置
秦王川国家湿地公园位于秦王川盆地南部,海拔较低,水资源丰富,是盆地自然降水、地下水潜流溢出、泉水溢流和引大入秦灌溉用水的主要汇水区,盆地蒸发量远大于降雨量,经过长期的水文过程,逐渐形成了全国罕见的陇中黄土高原半咸水内陆盐沼湿地。依照前期对土壤盐分分布规律的观测以及距离沙河河岸远近形成的天然盐分梯度的基础上,选择一处从沙河岸边至芦苇群落末端,面积约150 m×200 m且芦苇为单一优势种群的盐沼湿地进行实验。首先进行土壤盐分梯度的划分,垂直于沙河河岸,间隔20 m依次布置3条50 m×50 m的平行样带,进行土壤盐分的测定(在每条样带上从近河岸开始每间隔10 m采集表层土壤,测定土壤电导率)。根据土壤盐分变化规律,在3条平行样带上从河岸边向芦苇群落边缘依次设置3个20 m×20 m的样地(Ⅰ、Ⅱ和Ⅲ),分别为:(1)样地Ⅰ位于近水区湿地边缘,芦苇生长在岸边的沼泽地,该地段季节性淹水,进入生长旺季后,群落盖度在85%以上,土壤电导率介于1.9—2.2 ms/cm;(2)样地Ⅱ位于中水区过度地带,芦苇生长在沼泽地与盐碱土交接的生境中,土壤表层有轻微盐碱累积,植被稀疏,盖度在50%左右,土壤电导率介于2.5—3.6 ms/cm;(3)样地Ⅲ位于远水区芦苇群落末端,土壤表层盐碱累积,芦苇生长于盐碱土上,多成零星分布,分株矮小,盖度在20%左右,土壤电导率介于4.1—4.6 ms/cm。根据相关土壤盐渍化分级标准[30],土壤含盐量:4—8 ms/cm,属重盐渍土;2—4 ms/cm,属中盐渍土;1.8—2 ms/cm,属弱盐渍土。
1.3 野外采样
鉴于植被生长季节性和干旱区盐分的表聚性特点,采样时间为2015年8月,采样前8 d无降雨,土壤中水分、盐分状况相对稳定,利于研究工作开展。每个样地设置3个5 m×5 m的样方,现场记录所有植物的数量、株高、盖度等生态学指标并采集地上生物量。每个样方沿对角线法选取10株长势基本一致、生长良好的成年芦苇10株进行标记,参照Cornelissen的方法离植株基部直径约30 cm的范围用平板利铲挖取完整根系,将挖取的土柱置于塑料布上,保持根系的原始状态,用卷尺测量最大根深;然后用孔径为0.25 mm的尼龙网孔筛在附近河边把土柱的泥土全部冲洗干净,整株挑出,剪下地上部分与根系分别装入自封袋中迅速带回实验室4 ℃下保持。同步采集样方内表层土壤样品,在每个样方内沿对角线选取4个样点,分别用土钻(直径4 cm)每间隔10 cm分3层采集0—30 cm表层土样,重复3次,剔除所有样品中明显的植物根段和枯落物等杂质,现场称鲜重后装入编号的铝盒中,带回实验室。
1.4 实验方法
1.4.1 根系形态测定
在根系扫描仪根盘中放入少量蒸馏水,将根样置于根盘中浸泡,洗净粘附在根表面的土壤及杂质,并用细毛刷认真梳理根系使其在水中分布均匀,之后用根系扫描仪对根系进行扫描,应用Win-RHIZO(Pro 2009a, Regent Instruments, Quebec, Canada)软件进行数字化处理,定量分析根总长度、根平均直径、根分叉数。
1.4.2 芦苇生物量和比根长测定
芦苇地上部分自然风干,放入85 ℃恒温的烘箱内,直至重量恒重。根系形态分析完成以后,将植株根系于105 ℃杀青30 min,之后在入85 ℃恒温的烘箱内,烘至恒重,并根据以下公式计算根冠比和比根长:
根冠比=根干重/地上部干重
比根长=根总长度/根干重
1.4.3 土壤水分和盐分测定
带回实验室的土壤样品采用烘干法测定土壤含水量,电导率仪测定土壤含盐量(水土比为5∶1)。
1.5 数据处理方法
实验数据用SPSS 18.0进行统计分析,Origin 9.0软件进行绘图。不同样地的土壤盐分含量及芦苇生长特征参数通过单因素方差分析(one-way ANOVA)的Tukev法进行显著性检验。土壤盐分与生长特征参数的相关性分析采用SPSS 18.0的Person相关分析方法。
2 结果分析
2.1 湿地群落生物学特征与土壤水分因子统计学分析
各样地群落生物学特征与土壤含水量呈现规律性梯度变化,表现出不同程度的差异(表1)。不同样地群落的密度、高度、盖度、地上生物量差异性均显著(P< 0.05),表现为样地Ⅰ>样地Ⅱ>样地Ⅲ,由河岸至湿地末端呈递减趋势;土壤含水量差异显著,呈降低的趋势,由样地Ⅰ的50.23%减小到样地Ⅲ的30.27%,由于受地势和浅地下水埋深条件下的影响,土壤含水量总体很高,均大于30%,植被生长不存在水分短缺的限制。

表1 群落特征及水分因子的统计学参数(平均值±标准误差)
同列不同小写字母表示样地间差异显著(P< 0.05)
2.2 研究区土壤盐分状况
2.2.1 研究区土壤盐分含量分析
由表2可以看出,土壤含盐量的平均值为3.15 ms/cm,中值为2.26 ms/cm,二者差异较大,且平均值大于中值,说明土壤含盐量呈左偏态分布,因此采用中值作为衡量研究区土壤盐渍化指标较为适宜,研究区土壤属中度盐渍化水平。变异系数(CV)是描述变量空间变异程度的主要指标,土壤含盐量变异系数为1.05,表现为在水平方向上呈强变异性(根据变异系数的划分等级:CV > 1为强变异性,0.1 < CV < 1为中等变异性,CV < 0.1为弱变异性)。

表2 研究区土壤盐分统计特征
2.2.2 各样地土壤盐分含量及垂直分布特征分析
由表3可以看出,3个样地土壤盐分含量(0—30 cm土层)均值波动范围为2.11—4.18 ms/cm,样地Ⅲ土壤盐分含量均值为4.18 ms/cm,样地Ⅰ土壤盐分含量均值为2.11 ms/cm,极差达2.07 ms/cm,其变化趋势为:样地Ⅰ<样地Ⅱ<样地Ⅲ,样地间差异显著(P< 0.05),与盐渍化生境有关。各样地不同土层间的盐分含量呈现不同的差异性,样地Ⅲ不同土层间的盐分含量均存在显著差异(P< 0.05);样地Ⅱ中20—30 cm土层与其他土层间差异显著(P< 0.05),样地Ⅰ不同土层间的盐分含量差异均不显著。
各样地土壤含盐量随土层深度的增加总体呈降低的趋势(表4)。从变异系数来看,样地Ⅲ各土层深度土壤含盐量的变异系数为1.03、1.06、1.40,均达到强变异性,并随着土壤深度的增加则不断上升,说明土壤在垂直方向上变异性逐渐趋强;样地Ⅱ和样地Ⅰ各土层土壤含盐量均呈中等变异性,但随着土层深度的增加,样地Ⅱ变异系数有增强趋势,而样地Ⅰ变异系数逐渐趋弱。

表3 各样地土壤盐分含量特征
同行不同小写字母表示样地内差异显著(P< 0.05),同列不同大写字母表示样地间差异显著(P< 0.05)
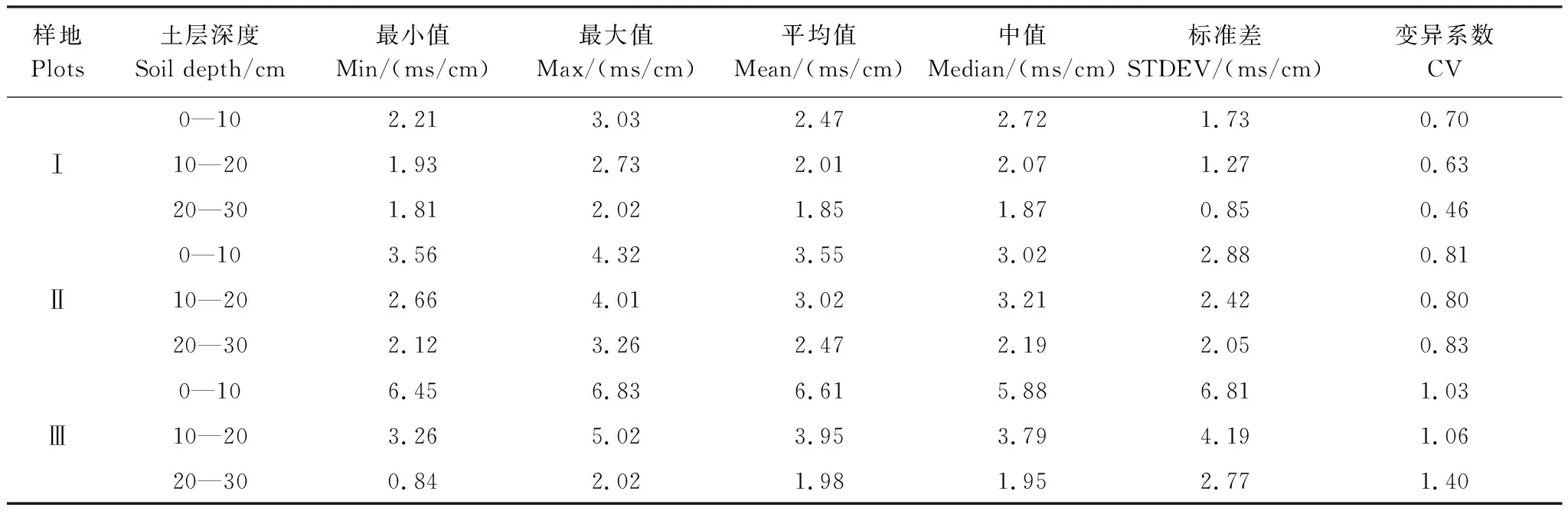
表4 各样地不同深度土壤盐分含量统计特征
2.3 不同盐分梯度下芦苇根系形态和生物量的变化
2.3.1 不同盐分梯度下芦苇根系生物量的变化
从图1可以看出,土壤盐分显著抑制了芦苇地上部和根系的生长(P< 0.05),抑制率分别达29%和42%,根系生物量减小相对地上部分更为明显,达到显著水平(P< 0.05),说明根系对土壤盐分的敏感程度高于地上部分。从样地Ⅰ到样地Ⅲ,随着土壤盐分含量的不断上升,根冠比总体呈下降趋势,但下降的速率有所减缓,在样地Ⅱ和样地Ⅲ间差异不显著,与土壤盐分的强变异性有关。
2.3.2 不同盐分梯度下芦苇根系形态的变化
从根系的几项形态指标来看,土壤盐分对芦苇根系的根总长度、比根长、根分叉数、根平均直径都有影响,且样地间差异显著(P< 0.05)(图2,图3)。在样地Ⅱ,根总长度、比根长和根分叉数参数比样地Ⅰ减小的较明显,均达到显著水平(P< 0.05),说明土壤盐分含量达到3.01 ms/cm对芦苇根系有强烈的抑制作用,可能达到了耐受阈值,其中最明显的变化是根总长度和根分叉数的减小,因为根总长度和根分叉数包括了所有侧根和分支根在盐胁迫下的变化。从样地Ⅰ到样地Ⅲ,随着土壤盐分含量的加剧,根总长度、比根长、根分叉数指标呈先下降后上升的趋势,而根平均直径呈先上升后下降的趋势。

图1 不同样地芦苇生物量的变化(平均值±标准误差)Fig.1 Change in biomass of Phragmites australis under different plot (mean±SE)

图2 不同样地芦苇根总长度和根深的变化(平均值±标准误差)Fig.2 Change in total roots length and root depth of Phragmites australis under different plot (mean±SE)

图3 不同样地芦苇根分叉数、根平均直径、比根长和根冠比的变化(平均值±标准误差)Fig.3 Change in root forks, root average diameter, specific root length and root-shoot ratio of Phragmites australis under different plot(mean±SE)
2.3.3 芦苇根系性状与土壤盐分相关性分析
表5数据表明,根总长度与土壤含盐量相关性达到极显著水平(P< 0.05),根冠比、根分叉数、比根长与土壤含盐量呈显著相关(P< 0.05),而根平均直径与土壤含盐量相关性不显著。

表5 土壤盐分与芦苇根系生长特征的相关性分析
*,在0.05水平显著相关;**,在0.01水平显著相关
3 讨论
植物性状特征是遗传因素和外界环境条件共同作用的结果[31],环境选择压力会促使植物性状通过权衡机制发生变化,进而调整自身性状间的功能组合和资源配置模式,形成相应的生态适应策略[32]。根系是植物一生中感受最敏感、调节最强的生长发育中心,特别是对逆境下植物的适应性生存尤为重要[33]。从河岸至湿地末端,土壤盐分含量具有很强的水平变异程度(表2),随着盐渍化程度的增加,芦苇根冠比、根分叉数呈逐渐减小的趋势,比根长、根总长度呈先下降后上升的趋势,而根平均直径呈相反的变化趋势。经Person相关性分析可知,土壤含盐量与根总长度呈极显著相关关系(P< 0.01),与比根长、根冠比、根分叉数呈显著相关(P< 0.05),而与根平均直径相关性不显著(表5),表明不同盐分梯度下芦苇根系形态和生物量分配的差异性,是植物根系构建的资源配置模式对盐分胁迫生境适应的结果。
生物量配置模式的自我调节是植物适应盐胁迫的可塑性机制之一,往往符合最优分配理论的预测[8, 34]。芦苇为典型的水生、盐生植物,对盐分的响应极为敏感,能通过不同器官生物量分配比例的变化来适应盐分环境。Mauchamp等学者研究发现芦苇通过减少根系生物量的分配以降低盐离子的过度吸收,也减少了盐分向地上部的运输,同时增加茎的生物量分配,将过多的盐分贮存在茎中[10]。本研究结果表明,离河岸愈远,土壤盐分含量逐渐增加,群落盖度、高度、地上生物量均有不同程度的下降,芦苇根冠比在各样地间存在差异(图1),并随盐分胁迫的加剧呈降低趋势,说明盐分胁迫造成了光合产物供给根系的减少,因而生长抑制程度根系大于地上部,这与Mauchamp等学者对芦苇的研究结果一致[10],而有关学者的研究结果则相反[27]。这主要是因为:(1)渗透胁迫和盐分离子毒害会导致根系吸收水分、养分能力的严重降低[17],氮、磷、钾等元素的缺乏会使碳水化合物不能顺利地运输到根、茎等部位,光合产物在细胞内大量积累,又给光合作用造成了反馈抑制,这可能是导致根冠比下降的一个重要原因;(2)有研究表明,盐分胁迫会迫使叶绿素含量降低,影响色素蛋白复合体的功能,从而降低光合速率导致光合产物减少[35],生物量向根系的分配也将减少,因此植株根生物量的减小更为显著;(3)与Mauchamp等学者研究结果一样,内陆盐沼的芦苇通过减少根生物量分配的调节机制来适应盐分胁迫环境,这种分配模式的积极意义在于降低了盐分对根系的损伤;(4)芦苇在适应盐胁迫环境中,可能分配给根状茎的生物量影响了地上部与根系的碳投入比例。进一步分析芦苇根冠比变化的速率发现:从样地Ⅱ到样地Ⅲ芦苇根冠比未随土壤盐分增加而显著降低,这与前人的研究结果略有不同[10, 36]。这主要是由于样地Ⅲ生境严酷,植被盖度、高度、密度和地上生物量最低,光照强烈使得土壤水分蒸发较快,盐分向表层迁移不断浓缩,表层土壤电导率高达6.61 ms/cm,属重盐土类型,而下层土壤电导率为1.98 ms/cm,属弱盐土类型,表层高盐会诱发植物根系发生严重的离子胁迫和渗透胁迫,从而影响植株的生长发育。在高光照和高盐等双重环境压力下,芦苇为了增加生存适合度,并没有降低光合产物供给根系的比例,而是将有限的生物量投资根系用于向深层土壤中拓展,来寻觅相对适宜的生存环境(图1),从而适应土壤盐分的高变异性(表4)。这种生物量配置策略既规避了高盐度对根系生长的抑制作用又增加了根系对深层土壤水分和养分的获取,也稀释了细胞内的盐分,从而增强了植物体在逆境胁迫下对盐渍环境的适应能力。因此,这种生物量调节机制,体现了内陆盐沼植物重要的自我保护策略。
在盐渍化条件下,植物通过根系功能的调整与权衡来适应环境的变化,其中根系形态的改变对植物的耐盐性具有重要意义[37]。大多数研究认为盐胁迫会抑制根系的生长,但也有报道表明,盐分胁迫能够促进侧根的发育,这可能与试验材料、植物发育时期及环境条件有关[9, 38- 39]。本研究结果表明,由样地Ⅰ向样地Ⅱ转变,随着土壤盐分含量的增加,芦苇根深、根分叉数、根总长度和比根长均显著下降,根平均直径则相反,其中根总长度变化最大(图2),芦苇的响应策略表现出敏感植物的特性。这主要是由于:(1)根总长度、比根长和根分叉数的减小,降低了根系对土壤空间的占据和利用量,可以限制盐离子的过多吸收,同时增加根系直径来发展通气组织能够减弱盐胁迫带来的缺氧损害[40],但这种功能权衡模式不利于根系吸收功能发挥,因此,芦苇如何通过根系形态的调整,达到既能抵抗盐胁迫又可以维持正常生理功能有待进一步研究;(2)根系的生长素平衡及分配格局会受到盐胁迫的扰乱,进而促进生长素在根伸长区的过量积累,抑制其重新循环,导致根伸长区细胞伸长受到抑制[41],限制了植物根系的扩展;(3)根总长度包括了所有不定根、侧根和分支根对盐胁迫的反应,因而在盐胁迫下变化剧烈,减小了50%。从样地Ⅱ向样地Ⅲ转变,土壤盐分含量逐渐升高,植被密度、盖度显著降低,有更多的土壤资源被释放出来供应芦苇根系的生长,芦苇在权衡资源捕获效率和生存适合度的基础上,采取增加根深、根总长度和比根长,减小根分叉数和根平均直径的优化配置模式来适应样地Ⅲ中盐分的强变异性。增大根深能够规避表层的高盐环境,促使根系延伸到土层20—30 cm处盐渍化程度较弱的空间,来降低盐分胁迫的风险;在生存环境较适宜的20—30 cm土层中,增大根总长度和比根长将有助于提高根系对土壤资源的获取能力,促进地上枝叶的生长,提高植株光合固碳效率,进而向根系分配更多的碳水化合物满足根系生长,提升抵御逆境的能力;降低根分叉数能够降低根系重叠程度,避免在狭小空间中根系内部不必要的竞争;配置较小的根直径,一定程度上将粗根转化为细根,能进一步提升芦苇吸收水分、养分的能力。因此,根系感知与响应胁迫的能力在很大程度上依赖于其对不断变化的土壤环境的适应。
4 结论
综上所述,芦苇通过表型可塑性形成了与生境相适应的根系形态和生物量分配格局,既抵抗了盐胁迫又维持了正常生存,其响应策略表现出敏感植物的特性。在生物量分配方面,随着土壤含盐量的增加,芦苇根冠比呈逐渐下降的趋势,这一分配模式对降低盐分的吸收,规避离子毒害风险具有重要意义。在根系形态方面,从样地Ⅰ到样地Ⅱ,芦苇根深、根分叉数、根总长度和比根长均显著下降,根平均直径则相反,芦苇根系的响应策略有利于限制盐离子的过度吸收和缓解缺氧损伤,从而增加生存适合度;从样地Ⅱ到样地Ⅲ,芦苇选择增加根深、根总长度和比根长,减小根分叉数和根平均直径的资源配置策略,利用根系塑性实现了空间拓展和资源吸收,最大限度的降低了逆境条件对植物的毒害,这主要与盐分高度空间变异有关。反映了植物应对胁迫生境的适应性机制,以及降低投资风险实现生存、繁衍的更新策略。芦苇地上地下生长发育是土壤水盐对其整个生长周期内的综合影响结果,今后有必要进一步研究芦苇整个生长周期内根系形态和生物量是如何对土壤水盐因子的变化产生响应。
