不同放牧与围封高寒灌丛草地土壤微生物群落结构PLFA分析
2018-08-09韦应莉曹文侠李建宏张爱梅李小龙
韦应莉,曹文侠,*,李建宏,张爱梅,李小龙
1 甘肃农业大学草业学院,草业生态系统教育部重点实验室,兰州 730070 2 西北师范大学生命科学学院,兰州 730070
土壤微生物是微生物界的重要成员,对土壤肥力的形成和转化起着积极作用。土壤微生物作为生态系统能量流动与物质循环的重要角色,受土壤环境和地上生物量的影响,对预测土壤有机质乃至土壤环境的变化过程敏感[1]。土壤微生物群落结构和功能的改变,可以进一步反映外界环境的变化,对于土壤生态环境的动态监测具有重要的指示作用[2- 5]。由于土壤微生物的数量巨大,组成极为复杂,用传统的土壤微生物研究方法往往会低估土壤微生物真实状况,给准确了解微生物群落结构带来困难[6]。磷脂脂肪酸(phospholipid fatty acids,PLFA)是活体微生物细胞膜恒定组分,它对环境因素比较敏感、能在生物体外迅速降解,因此特定菌群PLFA数量改变可反映原位土壤真菌、细菌活体生物量与菌群结构[7]。磷脂脂肪酸(PLFA)法具有对试验条件要求低、测试功能多和稳定性好等优点[8],被广泛应用于微生物动态检测中。王曙光等[9]主要对磷脂脂肪酸(PLFA)方法在土壤微生物分析中的应用做一剖析。颜慧等[10]将常用的研究微生物多样性的几种方法进行了比较,通过一些实例证明PLFA方法的特色或独到之处。
放牧是人类利用草地资源的主要方式,而不合理的放牧将使草地植被退化,导致土壤养分循环恶化[11- 12]。另外,不同强度放牧不仅对土壤微生物各类群数量产生影响,还影响微生物类群的组成[13]。围封作为退化草地生态系统恢复的重要手段,其对土壤微生物多样性及群落结构变化的影响逐渐成为近年来研究的热点之一。李春莉等[14]研究发现,适度的放牧使壤微生物数量增加,而不放牧和过度放牧不利于土壤微生物的生长。张成霞和南志标[15]对不同放牧强度下陇东天然草地土壤微生物三大类群的动态特征研究表明,围栏封育和适度放牧均有助于土壤微生物的恢复,使草地土壤微生物的数量增加,但过度放牧降低了土壤微生物的活性,导致微生物数量下降。祁连山作为青藏高原和北部内陆荒漠地区重要的分水岭,地处青藏、蒙新、黄土三大高原交汇地带,形成了独具特色的生物多样性和成土条件[16]。高寒杜鹃灌丛草地作为东祁连山主要地带性植被之一,其生态系统脆弱,退化程度日益加重,且恢复周期长[17]。放牧对高寒灌丛草地产生的影响如何,过度放牧后的围封恢复在土壤微生物方面有何响应并不清楚。本研究以高寒杜鹃灌丛草地为研究对象,旨在探讨不同强度放牧和围封对高寒灌丛草地土壤微生物群落及其结构特征的影响,从而揭示了微生物对灌丛草地围封和放牧后的变化规律和响应,为祁连山高寒杜鹃灌丛草地的合理利用提供有效措施和理论支撑。
1 材料与方法
1.1 研究区自然概况
试验地选在青藏髙原东部的天祝县金强河地区(37°11′ N,102°47′ E),平均海拔3200 m,年均温-0.1℃,最冷月(1月)平均气温为-18.3℃,最热月(7月)平均气温为12.7℃;≥0℃和≥10℃的年积温分别为1581℃和1026℃;年降水量416 mm,多为地形雨(其中66%集中在7—9月);年蒸发量1592 mm。无绝对无霜期,每年5月下旬植被进入返青期,10月上旬枯黄,生长季长120—l40 d。土壤为高山灌丛草甸土,显微碱性,土层较薄,为40—80 cm。植被分布受地形特征和气候差异等因素的影响,垂直分异明显,沿海拔梯度向上依次分布着珠芽蓼草甸、金露梅灌丛、杜鹃灌丛等。试验设在杜鹃灌丛草地,坡向西北。灌下生长着以问荆(Equisetumarvense)、珠芽蓼(Polygonumviviparum)、嵩草(Kobresiaspp.)、苔草(Carexspp.)等为优势种的草层植物。
1.2 样地设置与样品采集
2014年6月,选取坡向和海拔一致围栏高寒杜鹃灌丛草地作为研究样地,以相邻禁牧杜鹃灌丛为未放牧利用的参照草地,根据农业部放牧利用草地退化分级标准,综合考虑灌下可饲用草本植物利用率、可饲用灌木被采食率,灌丛地境土壤基况及家畜践踏程度[18],将样地划分为:重度放牧(HG)、中度放牧(MG)、轻度放牧(LG)。分别在不同放牧强度灌丛草地内沿海拔方向各设置3个100 m×30 m的围栏,共9个围栏,实施全年禁牧,作为恢复样地,继续观测,以围栏外的自由放牧围封灌丛草地为对照,放牧家畜为藏羊、牦牛。
土壤样品于2016年8月中旬采集,鉴于围封时间较短,而且草地土壤中微生物数量的垂直分布一般是上层大于下层[19],本试验取0—10 cm土壤,按照S型曲线选择40个点,去除表面植被,用直径为3.5 cm的土钻取样,最后将40份土样混匀过筛,去除根系和土壤入侵物,采用“四分法”选取1 kg土样冷藏带回实验室保存于-70 ℃超低温冰箱,用于测定土壤微生物PLFA分析;一份带回实验室风干,用于测定土壤有机质和全氮。
1.3 测定方法
1.3.1 PLFA的测定
土壤中脂肪酸的提取和检测主要分为4步:提取、分离、酯化、GC-MS分析[1- 2]。
主要试剂:37种脂肪酸混标(FAME)(Supelco,USA)和26种脂肪酸混标(BAME)(Supelco,USA)。
1.3.2 土壤有机质和全氮分别采用重铬酸钾-外加热法和半微量凯氏法。
1.3.3 数据分析
脂肪酸采用Frostegård方法命名[20]。PLFA可以作为微生物生物量和群落结构变化的生物标记分子[20-21]。根据已有的研究结果,指示特定微生物的PLFA生物标记物如下表(表1)。微生物生物量用各PLFA含量加和表示[25],另外,常用一些相关生理比值来分析磷脂脂肪酸(PLFA)数据,如(cyc17:0+cyc19:0)/(16:1ω7c+18:1ω7c)表示压力指数[26]、细菌/真菌、革兰氏阳性菌/革兰氏阴性菌(G+/G-)。用SPSS 19.0对试验数据进行方差分析、差异显著性比较(α=0.05)、主成分分析和相关性分析;采用Excel作图。

表1 不同微生物种群的磷脂脂肪酸标记
2 结果与分析
2.1 不同强度放牧和围封对高寒杜鹃灌丛草地土壤有机碳、全氮含量的影响
方差分析结果表明(表2),高寒杜鹃灌丛草地土壤有机碳和全氮含量随着放牧强度的增强而降低,且差异显著(P<0.05)。LG灌丛草地土壤有机碳含量比MG高44.0%左右,全氮含量比MG高25.0%左右;MG灌丛草地土壤有机碳比HG高38.0%左右,全氮含量比HG高22%左右。各个放牧强度下围封处理后的土壤有机碳和全氮含量略高于放牧处理,但均未达到显著水平。

表2 不同放牧强度下土壤有机碳、全氮含量/(g/kg)
同行不同小写字母表示差异显著(P<0.05)
2.2 不同强度放牧和围封对高寒杜鹃灌丛草地土壤磷脂脂肪酸数量的影响
如图1,各个样地均检测出18种脂肪酸(14:0,i15:0,a15:0,15:0,16:0,i16:0,i17:0,16:1ω7c,cy17:0,17:0,10Me16:0,10Me18:0,18:2ω6,18:1ω9c,18:1ω9t,18:0,cy19:0,20:0),其中都以16:0和10Me16:0的含量最高,i15:0,16:1ω7c,10Me16:0,16:0和18:1ω9c的总和占脂肪酸总量的64%—68%。由表3可知,构成脂肪酸的主要是6种支链饱和脂肪酸(BFA),绝对含量占总量的42.25%—47.10%,其中主要是i15:0和10Me16:0;其次是6种直链饱和脂肪酸(SFA),绝对含量占总量的23.18%—27.81%,其中主要是16:0和18:0;3种直链单不饱和脂肪酸(MFA),绝对含量占总量的21.18%—22.23%,其中主要是16:1ω7c和18:1ω9c;多不饱和脂肪酸(PFA)1种,绝对含量占总量的1.44%—2.12%;环丙烷脂肪酸2种,绝对含量占总量的4.61%—6.22%。
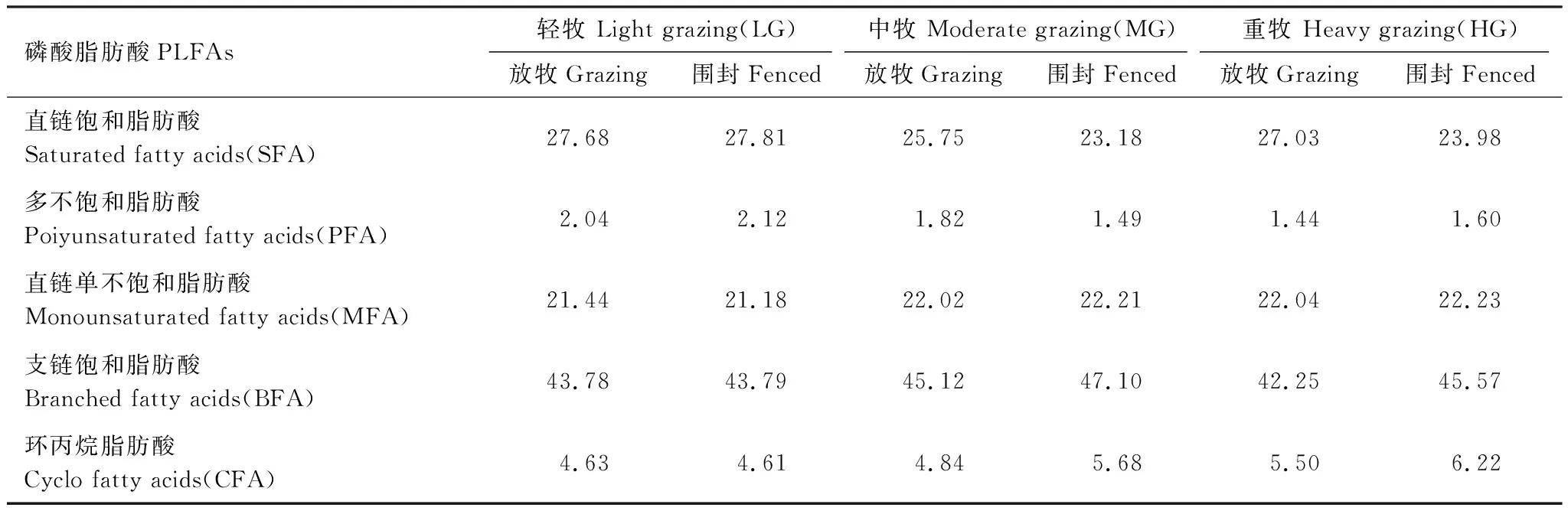
表3 不同样地土壤磷酸脂肪酸类型及其比例/%
由图1可知,脂肪酸14:0、15:0、16:0、17:0、18:0、18:2ω6,9、18:1ω9c、18:1ω9t、10Me16:0和10Me18:0的含量随着放牧强度的增强而降低,且达到显著水平(P<0.05)。围封处理下,i15:0、i16:0、i17:0含量在HG和MG间无显著差异,但显著高于LG灌丛草地;16:1ω7c(好氧细菌)含量在MG出现峰值,且差异显著(P<0.05),在LG和HG间无显著差异;(厌氧细菌)cy17:0和cy19:0的含量在各个放牧强度间存在显著差异(P<0.05),且cy17:0随着放牧强度的增强而升高;a15:0和20:0的含量在各个放牧强度间无显著差异;放牧处理下,脂肪酸i15:0、a15:0、、i16:0、i17:0、cy17:0和cy19:0的含量在各个放牧强度下随着放牧强度的增大而降低,且差异显著(P<0.05);16:1ω7c含量在MG灌丛草地出现最低值,且显著低于LG灌丛草地(P<0.05),但与HG差异不显著;20:0的含量在各个放牧强度间均无显著差异。HG强度下,围封处理后的18:0和20:0含量低于放牧处理,但差异不显著,而其他脂肪酸的含量都高于放牧处理,且都达到显著水平(P<0.05);MG强度下,围封处理后的14:0、15:0、16:1ω7c,、i15:0、i16:0、i17、10Me16:0、cy17:0和cy19:0的含量都高于放牧处理,16:0、17:0、10Me18:0、18:1ω9c、18:1ω9t 、18:2ω6,9和18:0的含量都低于放牧处理;LG强度下,围封处理后的18:1ω9c的含量高于放牧处理,且差异显著(P<0.05),而其他脂肪酸的含量与放牧处理无显著差异。
2.3 不同强度放牧和围封对高寒杜鹃灌丛草地土壤微生物群落结构的影响
磷脂脂肪酸的组成可以反映土壤微生物群落的细微变化,如生物量和分类结构的改变[27]。土壤微生物的生物量可通过直接从土壤中提取的磷酯脂肪酸的量来准确地表达。
2.3.1 不同强度放牧和围封对度高寒杜鹃灌丛草地土壤微生物PLFA总量的影响
由图2可以看出,方差分析表明,土壤微生物PLFA总量随着放牧强度的增强而降低,且达到显著水平(P<0.05)。围封处理下,LG灌丛草地土壤微生物PLFA总量分别比MG和HG高4.1%和13.8%,MG比HG高9.2%;放牧处理下,LG土壤微生物PLFA总量分别比MG和HG高12.4%和37.3%,MG比HG高22.2%。HG和MG强度下,围封处理后的灌丛草地土壤微生物总量分别比放牧处理高8.1%和20.9%,且达到显著水平(P<0.05);LG强度下,围封处理后的灌丛草地土壤微生物PLFA总量略高于放牧处理,但未达到显著水平。说明随着放牧强度的增大,土壤微生物的生物总量不断减少,各个放牧强度下围封处理后的微生物的生物总量都高于放牧处理,但在HG强度下围封处理对微生物总生物的提高作用比较明显。
2.3.2 不同强度放牧和围封对高寒杜鹃灌丛草地细菌生物量的影响
如图2,土壤细菌生物量随着放牧强度的增强而降低,且达到显著水平(P<0.05)。围封处理下,LG灌丛草地土壤细菌生物量分别比MG和HG高1.2%和6.1%,MG比HG高4.9%,在LG、MG和HG强度下,细菌生物量占微生物总量的百分数分别为59.7%、61.4%和64.0%;放牧处理下,LG土壤细菌生物量分别比MG和HG高15.3%和29.9%,MG比HG高12.7%,在LG、MG和HG强度下,细菌生物量占微生物总量分的百分数别为59.5%、58.0%和62.9%。HG和MG强度下,围封处理后的土壤细菌生物量分别比放牧处理高23.1%和14.6%,且差异显著(P<0.05);LG强度下,围封处理后的土壤细菌生物量略高于放牧处理,但差异不显著。可以看出,围封处理和放牧处理在不同放牧强度下土壤微生物细菌的生物量变化与微生物总生物量变化大致相同,这也证明了土壤微生物是以细菌为主体的群落结构。
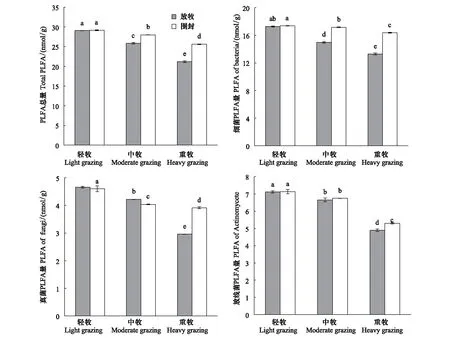
图2 不同样地土壤微生物各菌群生物量Fig.2 PLFA of each group of soil microorganisms in different sample sites相同字母表示差异不显著(P>0.5)
2.3.3 不同强度放牧和围封对高寒杜鹃灌丛草地土壤真菌生物量的影响
见图2,土壤真菌生物量随着放牧强度的增强而降低,且差异显著(P<0.05)。围封处理下,LG灌丛草地土壤细菌生物量分别比MG和HG高14.0%和17.5%,MG比HG高12.1%;放牧处理下,LG土壤细菌生物量分别比MG和HG高10.5%和57.4%,MG比HG高42.4%。HG强度下,围封处理后的土壤真菌生物量比放牧处理高32.2%,且差异显著(P<0.05);MG强度下,围封处理后的土壤真菌生物量显著比放牧处理低4.3%,且达到显著水平(P<0.05);LG强度下,围封处理后的土壤真菌生物量略高于放牧处理,但未达到显著水平。可以看出,围封处理在LG和MG强度下使土壤真菌生物量降低,而在HG强度下又使土壤真菌生物量显著增加,总的来看,不管是放牧处理还是围封处理,土壤真菌生物量都随着放牧强度的增大而降低,且各个放牧强度下围封处理对土壤真菌生物量影响作用各不相同。
2.3.4 不同强度放牧和围封对高寒杜鹃灌丛草地土壤放线菌生物量的影响
图2,土壤放线菌生物量随着放牧强度的增强而降低,且差异显著(P<0.05)。围封处理下,LG土壤放线菌生物量分别比MG和HG高5.7%和34.7%,MG比HG高27.4%;放牧处理下,LG土壤放线菌生物量分别比MG和HG高7.0和45.2%,MG比HG高35.7%。HG强度下,围封处理后的土壤放线菌生物量比放牧处理高8.1%,且差异显著(P<0.05);LG和MG强度下,围封处理后的土壤放线菌生物量略高于放牧处理,但未达到显著水平。结果表明,土壤放线菌生物量都随着放牧强度的增大而降低;围封处理能使灌丛草地土壤放线菌生物量增加。
2.3.5不同强度放牧和围封对高寒杜鹃灌丛草地土壤G+/G-的影响
由图3可知,围封处理下,土壤革兰氏阳性菌和革兰氏阴性菌的PLFA含量比值(G+/G-)在各个放牧强度间差异不显著;放牧处理下,G+/G-随着放牧强度的增强而降低,且在LG和MG强度间差异不显著,但显著高于HG强度。在HG强度下,围封处理后的G+/G-比放牧处理高32.2%,且差异显著(P<0.05);MG和LG强度下,围封处理后的G+/G-略高于放牧处理,但均未达到显著水平。各个强度放牧和围封的灌丛草地土壤中革兰氏阳性菌含量均高于革兰氏阴性菌含量,而HG围封处理后的灌丛草地G+/G-显著高于放牧处理,主要是由围封处理后的土壤中革兰氏阳性菌含量明显升高造成的。围封处理后的土壤G+/G-在各个放牧强度间较接近,不存在显著性差异,这说明围封处理可以维持土壤微生物群落结构。
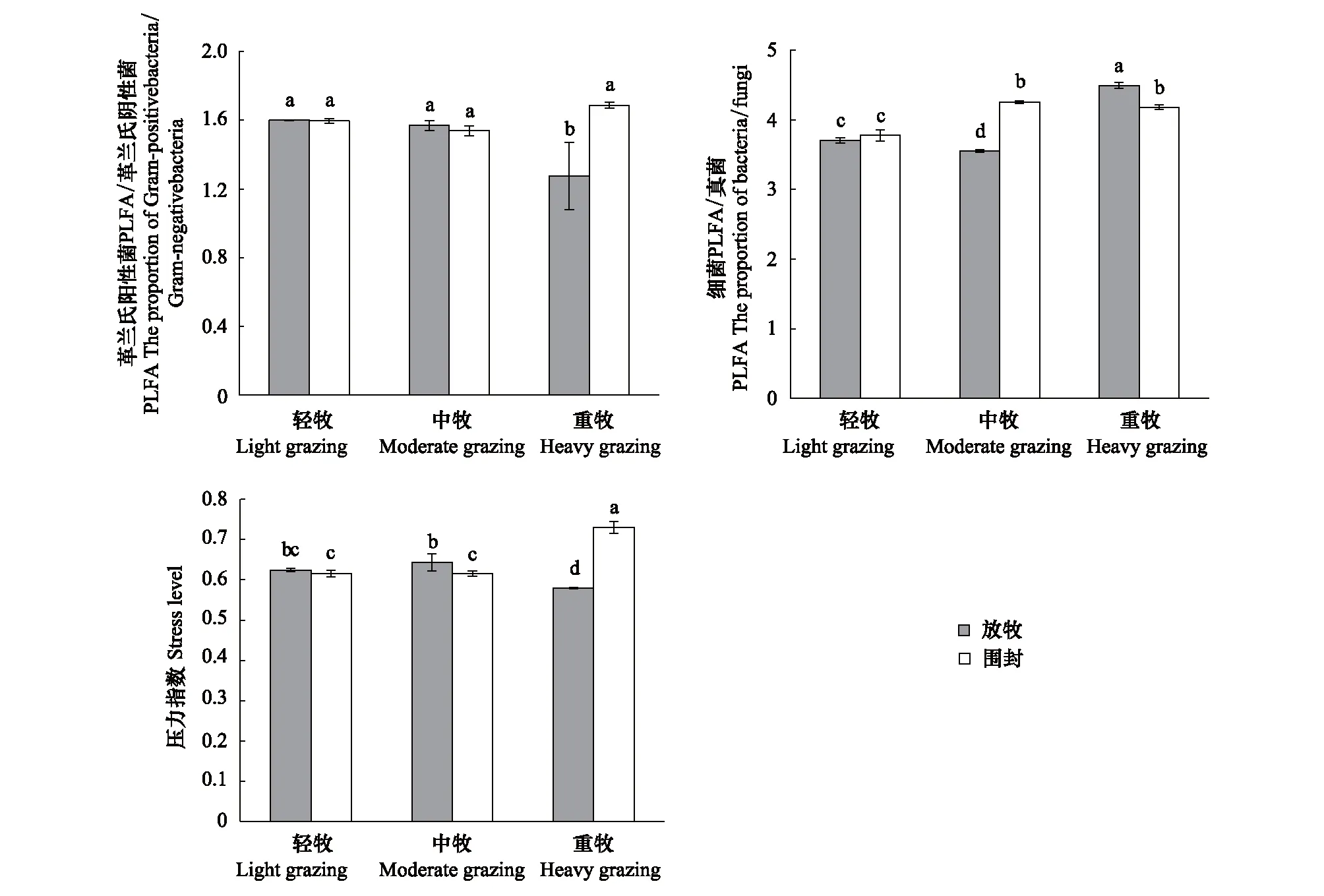
图3 不同样地土壤微生物各菌群生物量比值Fig.3 Ratios of each group of soil microorganisms in different sample sites各个菌群比值的相同字母表示差异不显著(P>0.5)
2.3.6 不同强度放牧和围封对高寒杜鹃灌丛草地土壤细菌/真菌生物量比值的影响
一般认为细菌/真菌比值可判断土壤生态系统稳定性和两个种群的相对丰富程度[28]。由图3可看出,围封处理下,细菌/真菌比值开始随着放牧强度的增强而升高,后趋于平稳;放牧处理下,细菌/真菌比值在各个放牧强度间差异显著(P<0.05),在HG强度下最高,在MG强度下最低。HG强度下,围封处理后的细菌/真菌比值比放牧处理低6.9%,且达到显著水平(P<0.05);MG强度下,围封处理后的细菌/真菌比值比放牧处理高19.8%,且差异显著(P<0.05);LG强度下,围封处理后的细菌/真菌比值略高于放牧处理,但未达到显著水平。说明不同强度放牧和围封对土壤生态系统稳定性及两个种群的相对丰富程度及有一定的影响。
2.3.7 不同强度放牧和围封对高寒杜鹃灌丛草地土壤微生物压力指数的影响
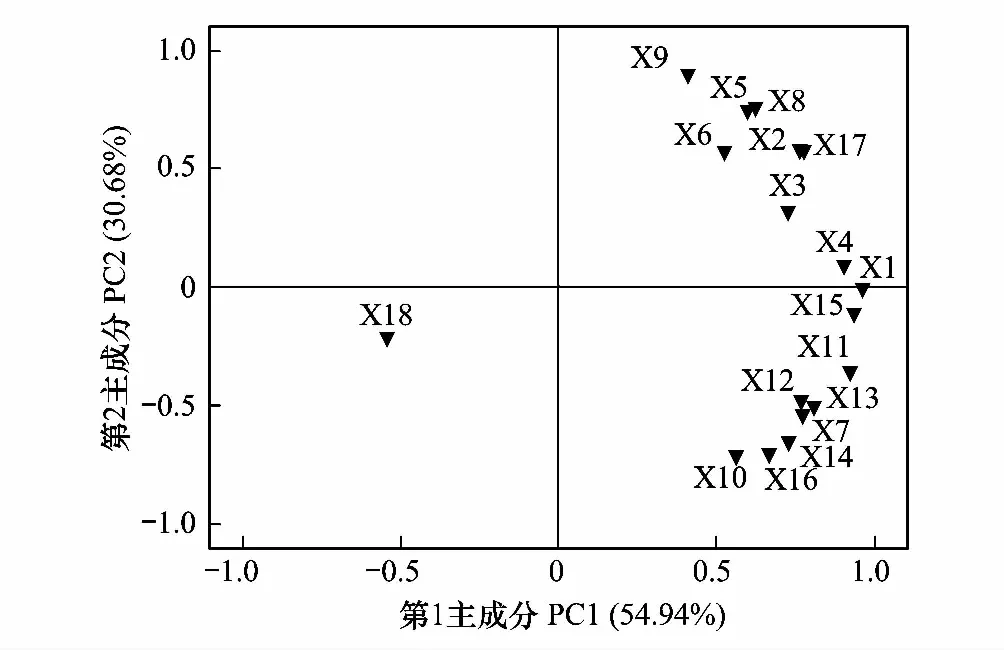
图4 不同样地PLFA主成分分析 Fig.4 Principal component analysis of PLFAs of different sample sitesX1:14:0;X2:i15:0;X3:a15:0;X4:15:0;X5:i16:0;X6:16:1ω7c;X7:16:0;X8:i17:0;X9:cy17:0;X10:17:0;X11:10Me16:0;X12:10Me18:0;X13:18:2ω6,9;X14:18:1ω9t;X15:18:1ω9c;X16:18:0;X17:cy 19:0;X18:20:0
微生物压力指数(cyc17:0+cyc19:0)/(16:1ω7c+18:1ω7c)作为评价土壤微生物群落对环境适应程度的具体指标。由图3看出,围封处理下,微生物压力指数随着放牧强度增强开始无显著差异而后又升高,且差异显著(P<0.05);放牧处理下,微生物压力指数随着放牧强度增强开始无显著差异而后又降低,且差异显著(P<0.05)。HG强度下,围封处理后的压力指数显著比放牧处理高26.0%,且差异显著;MG强度下,围封处理后的压力指数比放牧处理低4.5%,且达到显著水平(P<0.05);LG强度下,围封处理后的微生物压力指数比放牧处理略高1.3%,但未达到显著水平。
2.4 总PLFA主成分分析
供试土壤的18种脂肪酸进行主成分分析,PC1、PC2和PC3三个主成分合计达到土壤微生物群落结构组成的92.61%,其余7.39%为其他种群微生物。PC1和PC2总的方差贡献率为85.62%,因此PC1和PC2基本可以全面反映研究区域微生物群落结构特征,18种脂肪酸可以分成两个主要类型(图4)。其中中PC1主要由直链饱和脂肪酸、多不饱和脂肪酸、直链单不饱和脂肪酸、支链饱和脂肪酸环丙烷脂肪酸组成,占微生物群落结构变异的50.94%;其中14:0、15:0、10Me16:0和18:1ω9c的含量较高,占第一主成分的53.68%。PC2包括直链饱和脂肪酸、直链单不饱和脂肪酸、环丙烷脂肪酸和支链饱和脂肪酸,解释微生物群落结构组成变异的30.68%,主要由i16:0、16:1ω7c、i17:0、cy17:0、17:0、18:1ω9t、18:0和cy19:0脂肪酸组成,占第二主成分的51.34%。由特征向量与标准化后的数据相乘得出主成分表达式及公式:F=54.94%·FPC1+30.68%·FPC2+0.0699·FPC3,计算不同强度放牧和围封的总和得分F值(土壤微生物群落结构影响程度指数),并进行排序得到MG围封>HG围封>LG围封>LG放牧>MG放牧>HG放牧,即放牧和围封处理对高寒杜鹃灌丛草地土壤微生物群落结构的影响程度是围封处理大于放牧处理。由此可知,土壤微生物群落结构影响程度指数的排序结果与前面的论证结果一致,故主成分分析结果能较好地反映土壤微生物群落结构。
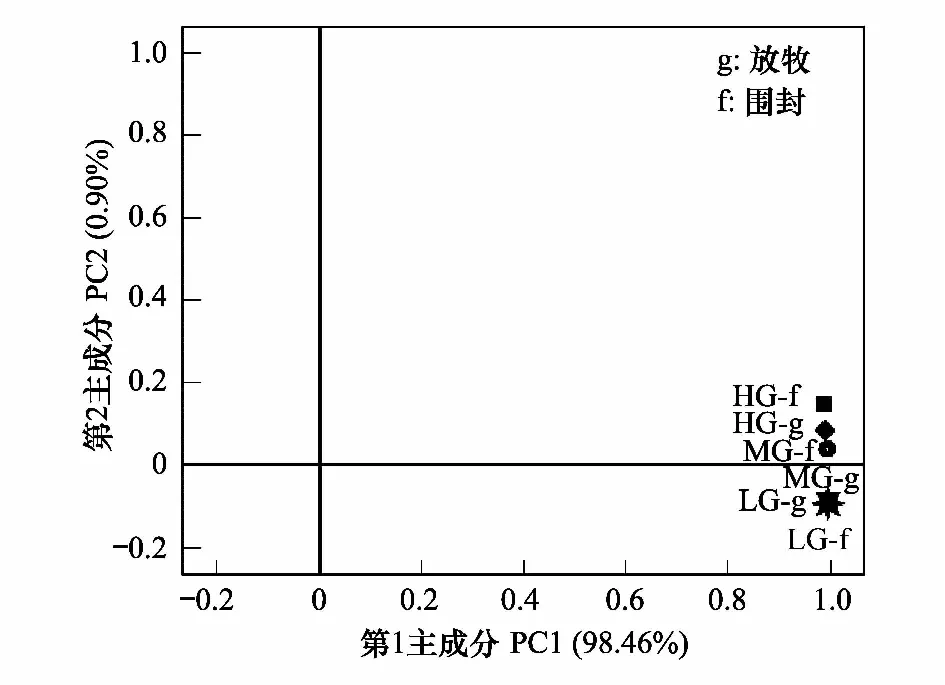
图5 样地间脂肪酸主成分分析 Fig.5 Principalcomponent analysis of PLFA structure among sites
不同强度放牧和围封地土壤磷脂脂肪酸的主成分分析(图5),结果表明各处理与第一主成分表现出正相关,HG围封、HG放牧和MG围封与第二主成分表示出正相关,其他处理与第二主成分表现为负相关。6种处理样地相距较近,且MG放牧、LG放牧和LG围封土壤的因子载荷十分接近,三者几乎重叠,但部分处理样地明显分开。说明不同强度放牧和围封改变了土壤微生物群落结构。
2.5 各菌群PLFA与土壤有机碳、全氮的相关性分析
本试验对不同放牧强度杜鹃灌丛草地各菌群PLFA与土壤有机碳、全氮进行相关性分析表明(表4),土壤总PLFA量、细菌、真菌和放线菌的PLFA与土壤有机碳、全氮均呈极显著正相关,细菌/真菌比值与有机碳、全氮呈极显著负相关。可用其来表征土壤这两项重要的肥力属性,也说明它们可用作评价土壤肥力与健康的生物指标,同时在这一地区土壤碳、氮含量也是影响土壤微生物数量和种类的最重要养分因素。
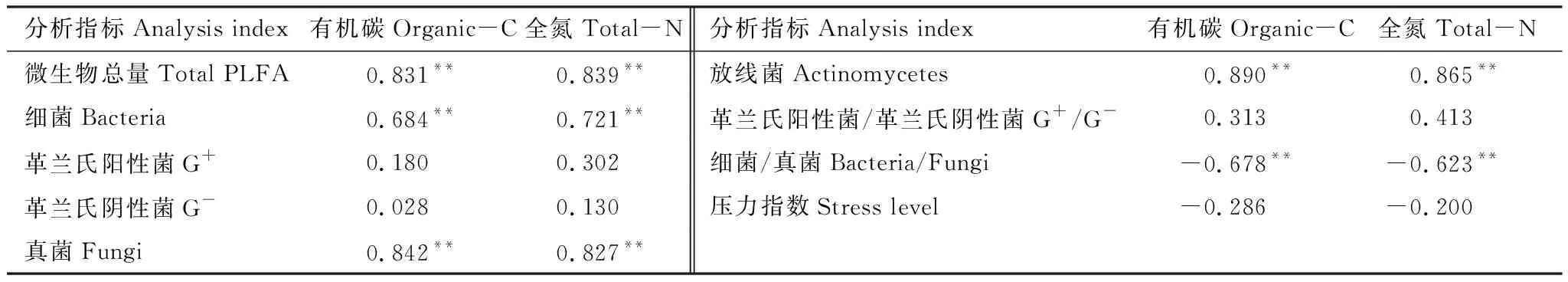
表4 微生物种群与土壤有机碳、全氮相关性分析
** 两尾检验,P<0.01
3 讨论
土壤微生物群落多样性可以检测系统受干扰后的细微变化,描述微生物群落变化和自然或人为干扰对群落的影响。高寒杜鹃灌丛草地土壤真菌生物量最少,占微生物总量的14.00%—16.32%,细菌生物量占微生物总量的57.97%—63.98%,放线菌生物量占微生物总量的20.47%—24.49%。以往的研究表明,在pH值较高的碱性土壤中细菌占据优势地位,反之则真菌占据优势地位[29]。姚拓和龙瑞军[30]采用平板表面涂抹法和稀释法对天祝高寒草地不同干扰生境土壤三大类微生物数量的时空动态进行了测定,结果表明三大类微生物中,细菌数量最大,放线菌次之,真菌最小,分别占微生物总数的82.29%—86.09%,13.91%—17.71%和0.017%—0.023%。无论是从微生物菌落的数量,还是从整体的PLFA群落结构分析(PCA),本研究都表明了放牧强度和围封对土壤微生物群落有显著的影响。
3.1 不同强度放牧对高寒杜鹃灌丛草地土壤微生物群落结构的影响
不同强度放牧对土壤结构的破坏程度有所不同,即高寒杜鹃灌丛草地围封区和放牧区因其放牧强度的不同结果不尽相同。围封处理下,土壤总PLFA量、细菌生物量、真菌生物量和放线菌生物量随着放牧强度的增大而降低,且差异显著(P<0.05);G+/G-PLFA量比值在各个放牧强度间无显著差异;细菌/真菌比值开始随着放牧强度的增强开始显著升高后无显著差异;微生物压力指数随着放牧强度增强开始无显著差异后又升高,且差异显著(P<0.05),主要是因为放牧强度增强,土壤的压实程度加重,从而降低土壤的孔隙度,扩散到土壤中的氧气的含量就比较少,好养细菌(16:1ω7c)的含量显著降低。放牧处理下,土壤总PLFA量、细菌生物量、真菌生物量和放线菌生物量随着放牧强度的增强而显著降低;G+/G-PLFA量比值随着放牧强度的增强而降低;细菌/真菌生物量比值在各个放牧强度间差异显著(P<0.05),在HG强度下比值达到最高,MG强度下比值最低,说明土壤生态系统在适度放牧下最稳定,过度放牧将低了土壤生态系统的稳定性;微生物压力指数随着放牧强度增强开始不变而后又降低,且差异显著(P<0.05),主要是由放牧强度增强,牲畜的践踏使土壤变得紧实,通气性下降,从厌氧细菌(cy17:0和cy19:0)的含量显著降低造成。萨如拉等[31]研究放牧强度对典型草原土壤微生物特征的影响,表明土壤微生物总数量随着放牧强度的增强而降低。但Wang和Sheng[32]对松嫩平原农牧交错带天然草地的研究发现,土壤细菌、放线菌和真菌数量随放牧强度的增加呈现先增多后减少的趋势,中牧地的土壤微生物数量最多。张成霞和南志标[15]对陇东黄土高原天然草地土壤微生物研究发现:土壤微生物总数和细菌数量随放牧强度的增大呈先减少后增多的趋势,而真菌和放线菌数量随放牧强度的增大而减少。谭红妍等[33]在温性草甸草原研究发现,土壤总微生物量、细菌和革兰氏阴性菌生物量均随着放牧强度的增加表现为先减少后增加的趋势,而革兰氏阳性菌、放线菌生物量则随着放牧强度的增加而增加。土壤微生物各菌落生物量因研究区域不同而有差异,且不同强度放牧使灌丛草地地上部分生物量的差异使其根系的分泌物明显不同,放牧强度不同其枯落物的质量也不同,以上两个方面均会影响杜鹃灌丛草地土壤微生物生长和代谢,从而影响微生物的数量和群论结构。另外,放牧牲畜的排泄物对土壤微生物也会造成一定的影响。
3.2 不同强度围封对高寒杜鹃灌丛草地土壤微生物群落结构的影响
HG强度下,围封处理的高寒杜鹃灌丛草地土壤总PLFA量、细菌生物量、真菌生物量、放线菌生物量、G+/G-PLFA和压力指数高于放牧处理,而围封处理的细菌/真菌比显著低于放牧处理,且均差异显著(P<0.05)。围封和放牧处理的HG灌丛草地土壤中G+PLFA量均高于G-PLFA量,且围封处理的HG灌丛草地土壤G+PLFA量和G-PLFA量均增加,但G+PLFA量增加的百分数大于G-PLFA量增加的百分数,导致G+/G-PLFA比显著高于放牧处理。土壤细菌与真菌比值越低,土壤生态系统越稳定[34-36]。此HG围封处理后的灌丛草地土壤细菌/真菌比值降低,说明生态系统稳定性升高。可能是因为围封处理的灌丛草地地上凋落物中含有更多的木质素和多酚类物质,这些物质能够诱导某些真菌生长,亦可能是围封没有牲畜的践踏,保证了真菌菌丝体的完整性使真菌菌丝体结构完整性的破坏减少,真菌生物量持续稳定增长,从而降低了细菌与真菌的脂肪酸比值。围封处理后的HG灌丛草地土壤厌氧细菌(cy17:0和cy19:0)和好氧细菌(16:1ω7c)均增加,但厌氧细菌增加的百分比大于好氧细菌,导致压力指数升高。MG强度下,围封处理后的土壤总PLFA量、细菌生物量、和细菌/真菌比值高于放牧处理,真菌生物量和压力指数低于放牧处理,且均差异显著(P<0.05)。围封处理的MG灌丛草地土壤细菌/真菌增大,可能因为围封处理后使动物排泄物减少,真菌吸收的营养减少,不利于真菌生长,导致真菌生物量有所降低,而细菌生物量增大,导致土壤生态系统的稳定性下降,微生物群落结构发生改变。围封处理的MG灌丛草地土壤厌氧细菌(cy17:0和cy19:0)和好氧细菌(16:1ω7c)均增加,但好氧细菌增加的百分比大于厌氧细菌,导致压力指数降低。LG强度下,围封处理后的土壤生物量和生物量比值均无显著差异。原因可能是由放牧强度较轻,围封作用不明显,围封内外土壤稳定结构没有显著差异所造成。赵帅等[37]在内蒙古针茅草原上的研究表明,围栏条件下,土壤细菌脂肪酸与总PLFA含量均显著高于放牧草地;放牧导致各类型草原革兰氏阳性细菌PLFA/革兰氏阴性细菌PLFA(G+/G-)比值显著降低,而除了克氏针茅草原,细菌/真菌PLFA则显著升高。究其原因可能为两地围封管理方式,放牧强度和草地类型不同所致,再者,放牧强度增加,牲畜践踏对土壤稳定结构的破坏程度增加,使土壤微生物生物量降低。张莉等[38]以放牧为对照,应用PLFA法分析研究围封后高寒草甸土壤微生物群落结构的变化,结果表明,围封处理的微生物总量、土壤细菌、G+、G+/G-和真菌均小于放牧处理,但其细菌/真菌和放线菌生物量高于放牧处理,围封处理降低了微生物的土壤生态系统的稳定性。由于牲畜践踏改变了土壤的紧实度,使土壤孔隙度和水稳性团聚体减少,引起透水性、透气性和水导率下降,土壤微环境改变,土壤微生物的繁殖代谢受到强烈的干扰,造成围封样地的土壤微生物PLFAs量显著高于放牧样地。Bardgett等[39]的研究发现:长期围封地表层土壤真菌丰富度和真菌/细菌PLFA值显著低于放牧地。此研究结果与本研究中MG杜鹃灌丛草地土壤真菌PLFA和细菌/真菌PLFA值研究结果一致。Bardgeett等[28]使用PLFA技术对不同放牧管理程度的草原土壤以及不同草原类型土壤微生物群落结构开展研究,结果显示放牧显著降低了土壤微生物PLFAs生物量,这与本研究结果一致。高凤等[40]对藏北古露高寒草地生态系统研究表明,短期围封使土壤微生物生物量(细菌、放线菌、真菌)均呈显著增加,能维持土壤微生物多样性。
主成分分析表明,不同的放牧强度和不同的利用方式(围封和放牧)对高寒杜鹃灌丛草地土壤微生物群落结构产生影响,且围封处理的影响程度大于放牧处理。说明利用方式和放牧强度对于微生物群落的影响不仅在于微生物生物量的变化,也影响微生物群落结构。
相关性分析表明,不同放牧强度下土壤结构及微生物可利用营养元素输入的变化是引起微生物群落结构变化的可能原因之一。韩世忠等[41]利用磷脂脂肪酸(PLFA)生物标记法在中亚热带地区罗浮栲天然林和相邻的杉木人工林研究表明,土壤微生物主要类群PLFAs含量与总氮、有机碳、C/N和铵态氮均呈显著正相关。吴则焰等[42]研究结果表明,真菌类群与土壤养分因子都呈负相关,且与总有机碳、全氮之间呈显著负相关,表明真菌比其他微生物类型更能适应养分贫瘠的条件。由于放牧强度的增强,在一定程度上限制了土壤微生物的活动,进而影响了生态系统中土壤养分和土壤结构等的变化。在轻度放牧灌丛草地,自然植被良好,地上生物量相对较大,土壤微生物数量大,生态系统的稳定性高。因此在当地独特的气候条件影响下,随着放牧强度的加重,植被种类结构趋向简单化,植被和微生物总量减少,土壤理化性质恶化,导致土壤全氮和有机质含量下降。
尽管PLFA技术常被用于研究土壤微生物群落结构特征,但该技术本身也有局限性。PLFA能定量描述环境样品中的微生物群体,而不能在菌种和菌株的水平精确的描述环境中微生物的种类,因此,要全面解析土壤微生物群落及其结构特征,还需结合BIOLOG鉴定系统和分子生物学等其他的研究手段。
