白及GMP基因cDNA全长克隆及生物信息学分析
2018-08-04石云平韦绍龙苏祖祥胡一凤李小泉
许 娟,石云平,韦绍龙,苏祖祥,林 茜,桂 杰,胡一凤,李小泉*
(1. 广西农业科学院生物技术研究所,广西 南宁 530007;2. 广西作物遗传改良生物技术重点开放实验室,广西 南宁 530007)
【研究意义】白及Bletillastriata(Thunb.)Reichb.f.为兰科多年生草本植物,具有上千年药用历史,在本草经集注、本草图经及本草纲目中都有记载[1-2]。白及以块茎(假鳞茎)入药,块茎组织中含40 %~50 %天然水溶性多糖。目前已从白及中分离出20多种糖苷类化合物,其中白及多糖是报道最多的化合物,也是目前最有开发前景的药物。白及多糖具有促进人类脐静脉内皮细胞生长及血管内皮细胞生长因子的表达[3]、缓和炎症反应[4]等功能。近期有临床研究报道白及胶可作为外源性重组人表皮生长因子载体,能促进创面表皮细胞DNA合成,加速伤口愈合等[5]。因此,研究白及多糖合成的分子机制对培育富含多糖的白及新品种具有重要意义。【前人研究进展】白及多糖以甘露聚糖家族的葡萄甘露聚糖为主,GDP-甘露糖的合成是白及多糖合成的第一步,GDP-甘露糖由GDP-甘露糖焦磷酸化酶(GMPase)催化甘露糖-1-磷酸而成,GMPase是甘露聚糖合成的关键酶[6]。1956年,Munch-Petersen[7]首次从brewer酵母中分离到一种可以催化甘露糖-1-磷酸和GTP生成GDP-甘露糖的酶,1964年其正式被命名为GDP-甘露糖焦磷酸化酶。1996年,酵母中的GMP基因首次被克隆到[8],随后,其它植物中的GMP基因也被克隆出来,如拟南芥[9]、马铃薯[10]、番木瓜[11]、水稻[12]和番茄[13]等。何春梅[14]克隆了2个铁皮石斛GMP,分别命名为DoGMP1和DoGMP2,DoGMP1的cDNA全长为1528 bp,开放性阅读框(ORF)1086 bp,编码362个氨基酸的蛋白;DoGMP2的cDNA全长为1492 bp,开放性阅读框(ORF)1248 bp,编码416个氨基酸的蛋白。【本研究切入点】目前有关白及多糖合成的研究较少,白及GMP基因的克隆也未见报道,白及GMP基因的序列及结构等分子生物学特性尚不明确。【拟解决的关键问题】以白及叶片为材料,通过同源基因比对,设计兼并引物,利用RACE-PCR方法从白及中克隆GMP基因的cDNA全长,并进行生物信息学分析,为该基因在白及中的功能研究奠定基础,为培育富含多糖的白及新品种提供理论依据。
1 材料与方法
1.1 供试材料
1.1.1 试验材料 采用“桂及1号”白及为试验品种,采取广西农业科学院生物技术研究所种植基地生长1年以上植株的新鲜叶片,用水清洗处理并擦干,迅速放入液氮中速冻,并装袋于-80 ℃超低温冰箱中贮存备用。
1.1.2 主要试剂 RNAprep Pure多糖多酚植物总RNA提取试剂盒,购自天根生化科技(北京)有限公司;pMD19-T载体、dCTP和M-MLV逆转录酶,购自TaKaRa公司;dNTP、TaqDNA聚合酶和氨苄青霉素(Amp),购自上海生工生物工程有限公司;SMARTerTMRACE扩增试剂盒,购自Clontech公司;目的条带胶回收试剂盒,购自QIAGEN 公司;Tris-HCl、EDTA、琼脂糖等试剂为市销售进口或国产分析纯。引物合成及DNA测序由生工生物工程(上海)股份有限公司完成。
1.1.3 引物设计 采用Primer 5和Oligo 6.0设计基因克隆所用引物。具体引物序列见表1。
1.2 试验方法
1.2.1 总RNA的提取及cDNA合成 用1.2 %琼脂糖凝胶电泳检测总RNA完整性,并利用NANODROP 1000核酸蛋白检测仪测定其浓度和纯度。参照逆转录酶M-MLV说明书合成cDNA,引物为Oligo(dT)18。
1.2.2 白及GMP基因中间片段的PCR扩增 根据NCBI已登录的其它植物GMP基因序列的保守区设计简并引物GMP-F和GMP-R(表1)。PCR扩增反应总体系为25 μl:cDNA模板1 μl,dNTP(10 mmol/L)0.5 μl,GMP-F,GMP-R(10 mmol/L)各0.5 μl,Taq酶0.2 μl,缓冲液2.5 μl,双蒸水19.8 μl。扩增程序:94 ℃,4 min;94 ℃,30 s;55 ℃,30 s;72 ℃,1 min,36个循环;72 ℃,8 min。取5 μl PCR产物进行琼脂糖凝胶电泳检测。
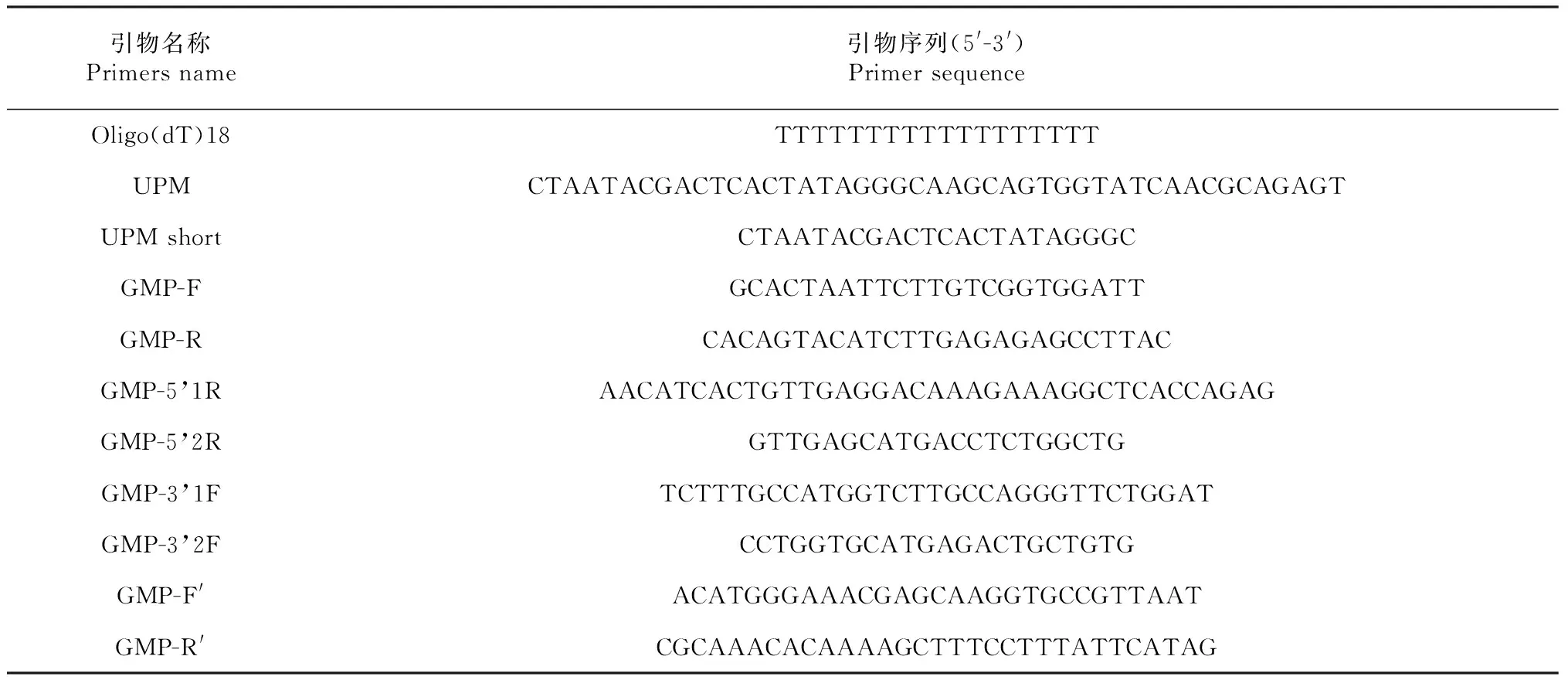
表1 引物序列
1.2.3 中间片段回收、测序 将PCR产物用QIAGEN PCR纯化试剂盒回收,与pMD19-T载体连接,转化大肠杆菌感受态DH5α,涂在含有100 μg/mL Amp的LB固体培养基上,37 ℃倒置培养过夜,挑取单菌落,在含Amp的液体LB培养基上37 ℃摇菌4 h,经菌液PCR检测,将阳性克隆送上海生工公司测序。
1.2.4 白及GMP基因5′与3′末端RACE扩增 根据获得的白及GMP基因的中间序列,分别设计5′端和3′端特异性巢式PCR引物GMP-5′1R、2R和和3′端特异性巢式PCR引物GMP-3′1F、2F(表1),以白及叶片cDNA为模板,经2轮巢式PCR扩增,分别扩增GMP基因的5′端和3′端序列,具体按照Clontech公司SMARTerTMRACE扩增试剂盒说明书进行,将目的片段进行回收、连接、转化和鉴定后,送阳性克隆到上海生工进行测序。
1.2.5 白及GMP基因cDNA全长的获得 根据中间片段、5′与3′末端的测序结果,用Vector-NTI对序列进行比对和拼接,最后得到cDNA全长,并设计引物GMP-F′和GMP-R′(表1)扩增其全长序列,并测序进行验证。
1.3 白及GDP-甘露糖焦磷酸化酶同源基因生物信息学分析
用Premier 5和Oligo 6.0软件进行引物设计,用DNAmanV6软件进行序列拼接和序列分析。利用ORF Finder在线工具对获得的cDNA序列进行开放阅读框(ORF)查找分析;用BLAST在线工具(http://www.ncbi.nlm.nih.gov/Blast.cgi/)搜索GenBank的同源序列;用BioXM 2.6对氨基酸序列进行预测;用TMpred分析该基因编码氨基酸组成、理论分子量和等电点等理化性质;通过Softberry服务器分析白及GDP-甘露糖焦磷酸化酶蛋白的亚细胞定位;运用NCBI的Conserved Domain Database(CDD)在线工具进行功能结构域分析;使用SWISS-MODEL在线工具预测蛋白质的三维结构;利用MEGA 5.0软件构建进化树。
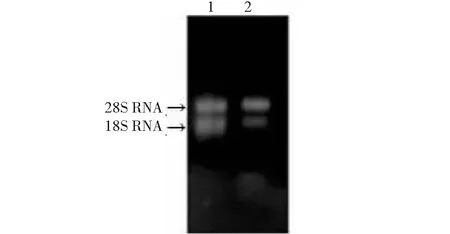
1~2: 提取RNA样品1-2: Extracted RNA samples图1 白及“桂及1号”叶片总RNA琼脂糖凝胶电泳结果Fig.1 Agarose gel electrophoresis of total RNA from Bletilla striata leaves
2 结果与分析
2.1 总RNA的提取及检测
以生长1年以上“桂及1号”白及植株的新鲜叶片为材料,进行叶片总RNA提取,经1.2 %琼脂糖凝胶电泳检测(图1),可见28S rRNA和18S rRNA条带明亮清晰,说明所提取的RNA完整性较好。经NANODROP 1000核酸蛋白检测仪测得OD260/OD280为2.05,OD260/OD230为1.84,表明RNA纯度较高,可以用于RT-PCR扩增实验。
2.2 白及GMP基因全长的克隆
以白及叶片总RNA反转录所得到的cDNA为模板,进行PCR扩增。扩增产物经凝胶电泳检测发现约870 bp的条带(图2),与预期片断的大小一致,目的基因片段经过连接、转化及菌液PCR鉴定,得到阳性克隆(图3),经测序,目的基因大小为873 bp,经BLAST比对分析,发现该序列与铁皮石斛、蝴蝶兰的GMP基因同源性很高。根据获得的白及GMP基因中间片段的序列,分别设计5′端和3′端特异性巢式PCR引物,以cDNA为模板,经2轮巢式PCR扩增3′端和5′端基因序列,得到3′端基因序列长度为683 bp的片段和5′端基因序列长度为483 bp的片段(图4),将其与中间片段序列进行拼接,得到1523 bp的cDNA全长,对其进行扩增(图5),所得条带与预期片段大小一致,测序结果表明拼接无误。将本试验获得的白及GMP基因命名为BsGMP。

M: DL2000DNA marker; 1:扩增产物M: DL2000 DNA marker; 1: The fragment of RT-PCR图2 白及GMP基因中间片段扩增结果Fig.2 Amplification results of partial GMP gene of Bletilla striata
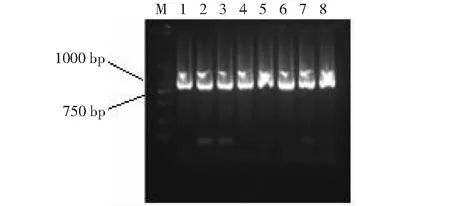
M: DL2000 DNA marker; 1~8:阳性克隆M: DL2000 DNA marker; 1-8: Positive clones图3 菌液PCR结果Fig.3 PCR results of bacteria solution

M: DL2000 DNA marker; 1:3′ 端序列扩增产物;2:5′ 端序列扩增产物M: DL2000 DNA marker; 1: Amplification products of 3′ RACE; 2: Amplification products of 5′ RACE图4 白及GMP基因两端部分序列扩增结果Fig.4 Amplification results of two ends of GMP gene
2.3 BsGMP同源基因的生物信息学分析
在NCBI的ORF finder软件上查找BsGMP的编码区,发现该基因的ORF为1086 bp,该基因所编码的蛋白含有361个氨基酸(图6),分子量为39.35 kDa,等电点为6.03,含量最大的3种氨基酸为:Val(11.9 %)、Leu(9.7 %)和Gly(9.1 %);亚细胞定位分析表明,BsGMP编码的蛋白定位于细胞质,在线粒体中也有分布。Tmpred分析显示该蛋白有2个可能的内部到外部跨膜螺旋区和2个可能的外部到内部跨膜螺旋区。利用NCBI的CDD在线工具进行BsGMP蛋白功能保守结构域分析,结果表明该蛋白含有醛酮还原酶的保守结构域(第1~24个氨基酸)和LbetaH家族保守区(第256~335个氨基酸)(图7)。
将氨基酸序列导入SWISS-MODEL在线分析软件中进行比对建模,用RasMol 2.7.5软件对BsGMP蛋白的三级结构进行显示(图8),可以看出,BsGMP蛋白具有11个典型的α螺旋,33个β折叠,同时具有46个β转角。其中α螺旋用深红色表示,β折叠用黄色表示,转角用淡蓝色表示,其他残基用白色表示。

M: DL2000 DNA marker; 1:基因全长序列M: DL2000 DNA marker; 1: Gene full-length sequence图5 白及GMP基因cDNA全长序列扩增(1523 bp)Fig.5 Amplification results of Bletilla striata GMP gene full-length(1523 bp)
2.4 BsGMP进化分析
将BsGMP序列与NCBI上其他物种的GMP基因序列进行比较,发现相似系数为64.0 %~95.0 %,其中与铁皮石斛和蝴蝶兰的相似系数最高达95.0 %。用MEGA 5.0软件对不同物种GMP基因编码的氨基酸序列构建进化树(图9),发现BsGMP与已报道的铁皮石斛和蝴蝶兰同源性最近,因而最先聚在一起,草莓、甜樱桃、金丝枣、核桃、胡杨和马铃薯也聚为一类,紫花苜蓿和拟南芥则被单独聚为一类,说明BsGMP基因与拟南芥亲缘性最远,与兰科植物GMP基因的亲缘性较近。
3 讨 论
GMP基因具有较高的保守性,GMPase催化所得产物 GDP-甘露糖,参与植物体内多种代谢途径,如蛋白质糖基化作用、抗坏血酸、细胞壁和甘露聚糖的合成等。研究结果表明GMP与植物的逆境胁迫响应有关:李超汉等[15]发现番茄GMPase基因在马铃薯中的超表达,可使马铃薯抵御温度胁迫能力明显提高;张琳[16]对番茄中GMPase基因进行功能分析,转正义T1代植株,GMPase酶活性和光合性能明显增加,果实中可溶性糖的含量和糖酸比显著增加,有机酸的含量显著降低;Wang等[17]研究发现转基因烟草GMP低水平表达时,对高温或低温更敏感;何春梅[14]对DoGMP1进行功能验证,发现DoGMP1在拟南芥中的过表达不仅可以提高拟南芥的抗坏血酸含量和多糖含量,还可以提高其种子在盐胁迫下的萌发率和植株对盐胁迫的耐受性。国内外关于GMPase基因的研究还不多,只在少数作物如拟南芥[9]、马铃薯[10]、番木瓜[11]和水稻[12]等中克隆到了GMPase基因。Zou等[18]从番茄中克隆了GMPase基因的cDNA全长,共1535 bp,包含1086 bp的ORF,编码361个氨基酸残基,该基因推导的氨基酸序列经BLAST比对发现,其与马铃薯GMPase基因的同源性最高,达99 %,与烟草、紫花苜蓿和拟南芥的GMPase基因的同源性分别为97 %、91 %和89 %。本试验利用RACE方法,首次从白及中克隆到一个GDP-甘露糖焦磷酸化酶基因(BsGMP),其cDNA全长1523 bp,其中包含1086 bp的完整ORF,编码361个氨基酸的多肽,理论蛋白分子量为39.05 kDa,等电点为6.03,呈酸性。BLAST比对结果表明BsGMP氨基酸序列与其他物种氨基酸序列相似性为64.0 %~95.0 %,氨基酸同源性数据分析结果表明BsGMP基因分化后在不同物种内仍执行着相近的功能,与铁皮石斛、蝴蝶兰序列的相似性最高,达90.0 %以上,并聚为一支,说明这3个物种为一个科,同属兰科植物,与所对比其他物种亲缘关系较远。

图6 BsGMP序列及其推导氨基酸序列Fig.6 Bletilla striata GMP gene sequence and its deduced amino acid sequence
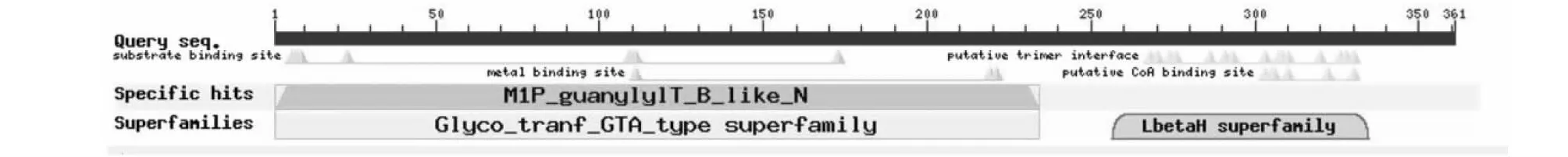
图7 BsGMP蛋白结构域预测Fig.7 Conserved domains of BsGMP
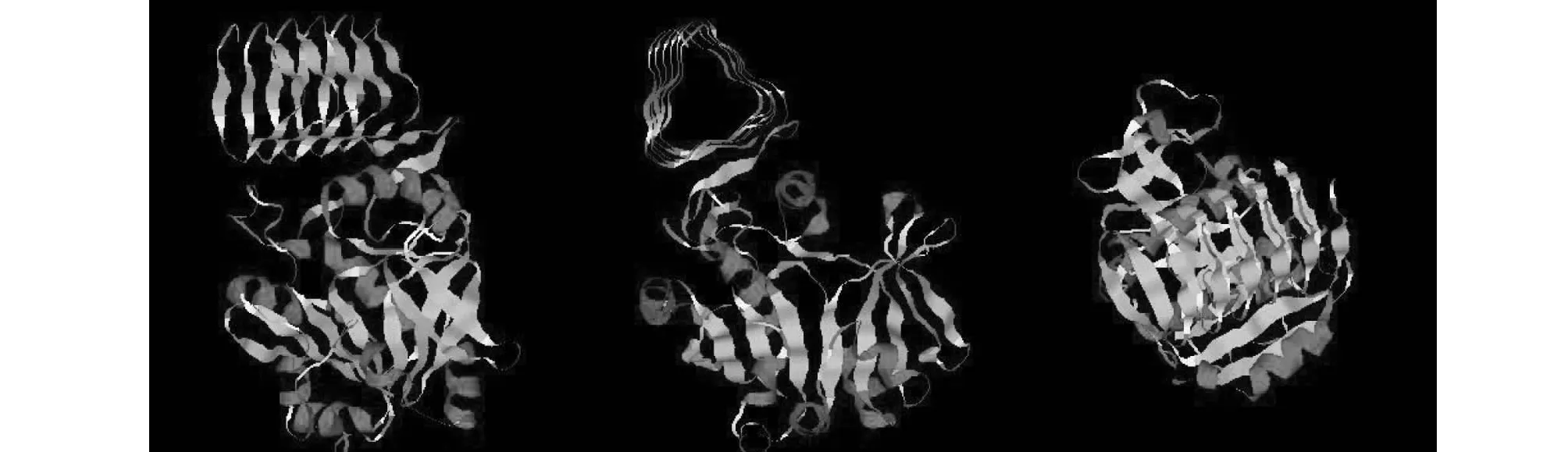
图8 白及GMP蛋白三维结构Fig.8 Three dimensional structure of GMP protein from Bletilla striata
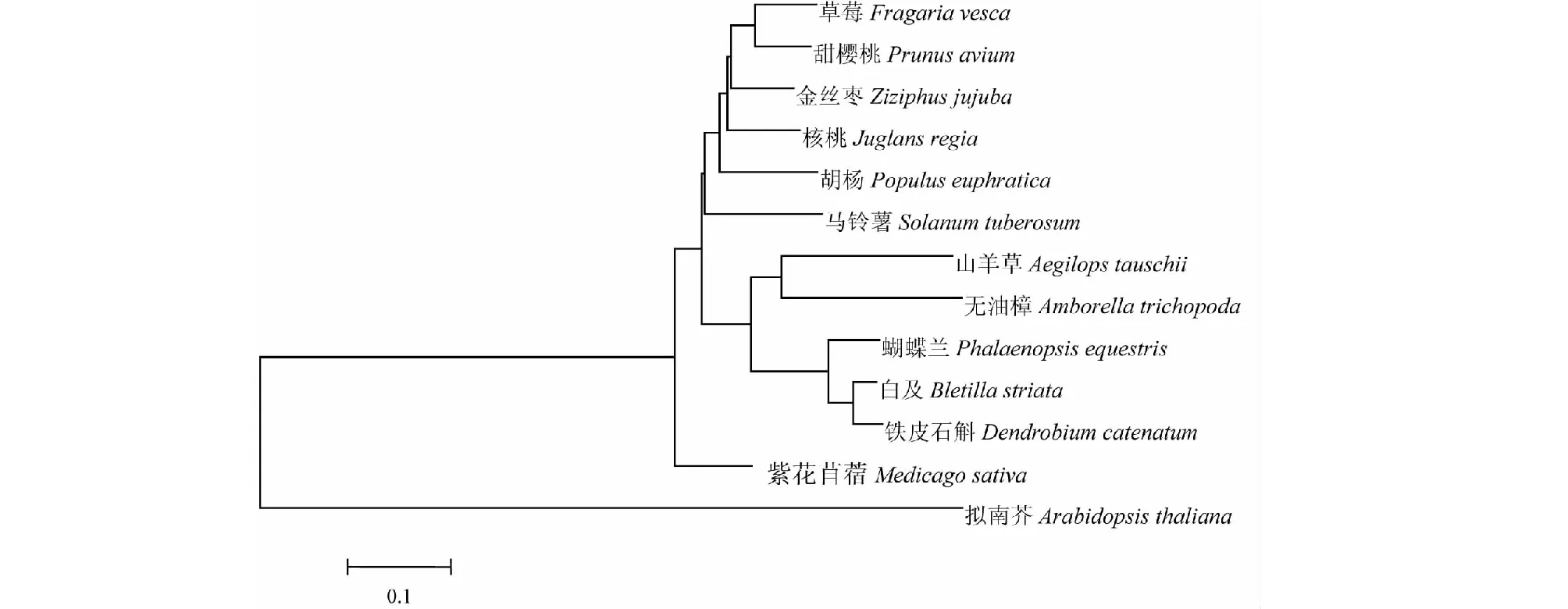
图9 基于GMP基因同源性的系统进化树Fig.9 Phylogenetic analysis of GMP gene from Bletilla striata
生物体中存在2种类型的GDP-甘露糖焦磷酸化酶基因,分为A型和B型。何春梅[15]克隆了3个铁皮石斛GMP基因,分别命名DoGMP1、DoGMP2和DoGMP3,其中DoGMP1 和DoGMP3属于B 型,而DoGMP2属于A型,但3者都具有较高的同源性;对DoGMP1蛋白进行亚细胞定位,发现该蛋白与其它植物GMP家族的成员相似,主要分布在细胞质中。根据本试验结果对蛋白结构进行预测及分析,发现该蛋白有2个可能的内部到外部跨膜螺旋区和2个可能的外部到内部跨膜螺旋区,BsGMP编码的蛋白定位于细胞质,在线粒体中也有分布。BsGMP蛋白功能保守结构域的分析表明,该蛋白含有醛酮还原酶的保守结构域(第1~24个氨基酸)和LbetaH家族保守区(第256~335个氨基酸),这些保守区在细胞壁多糖合成、蛋白糖基化及蛋白转录后调控等过程中具有重要作用[19-20]。特定蛋白有不同的螺旋数与转角数,结构的不同直接影响基因功能的不同,因此,下一步需要通过转基因方法来验证其功能,这对于进一步了解白及甘露糖合成关键酶基因GMP的功能及多糖合成分子机理具有重要意义。
随着生物技术的迅速发展,RACE技术为基因克隆提供了方便简洁的方法。操作中尤其要对逆转录产物进行纯化,以及在目的产物扩增中设计好特异引物,避开发夹结构和引物二聚体,防止产生大量非特异性或截短的产物。基因克隆技术的成熟完善,我们可以较易地通过克隆目的基因,并进一步研究基因的功能结构,今后可利用转基因技术提高白及GMPase的活性,通过增加多糖生物合成前体GDP-甘露糖的供应,达到大幅度提高白及块茎多糖含量的目的,为培育富含多糖的白及新品种提供理论依据。
4 结 论
从白及叶片中克隆到1个GMP基因的全长cDNA,命名为BsGMP,其长度为1523 bp,包含1086 bp的完整开放阅读框,编码361个氨基酸。BsGMP氨基酸序列与其他物种氨基酸序列相似性为64.0 %~95.0 %。预测BsGMP编码的多肽分子量为39.05 kDa,含量最大的3种氨基酸为:Val、Leu和Gly,等电点为6.03,定位于细胞质,为不稳定蛋白,在线粒体中也有分布,并有2个可能的内部到外部跨膜螺旋区和2个可能的外部到内部跨膜螺旋区。预测BsGMP蛋白有11个α螺旋,33个β折叠,46个β转角,从第1~24个氨基酸含有醛酮还原酶的保守结构域,第256~335个氨基酸含有LbetaH家族保守区,这些保守区是BsGMP蛋白酶活的结合区域,主要参与细胞壁多糖合成、蛋白糖基化及蛋白转录后调控等。
