北方冬麦区小麦品种产量相关性状和幼穗分化特点研究
2018-07-27买春艳李洪杰刘宏伟于立强张宏军
买春艳,李洪杰,刘宏伟,杨 丽,于立强,周 阳,张宏军
(1.新乡矮败小麦育种技术创新中心,河南新乡 453731;2.中国农业科学院作物科学研究所/农作物基因资源与基因改良国家重大科学工程,北京 100081;3.石家庄市农林科学研究院赵县试验基地,河北赵县 051530)
小麦作为重要的粮食作物,为全世界人口提供20%的热量和蛋白质[1]。据2013年联合国粮农组织(FAO)预计,小麦产量必须以1.6%的年增长率才能满足人口不断增长的需求。因此,在维持种植面积不变的条件下,小麦产量需要从目前的每公顷6 000 kg增长到10 000 kg[2]。我国是世界上最大的小麦生产和消费国,年种植面积达2.4×109hm2,平均产量4 672 kg·hm-2[3]。随着人口的增长、可用耕地的减少,提高单位面积产量是确保我国粮食安全的必然要求[4]。因此,高产是小麦育种的长期目标。
小麦产量主要由有效穗数、穗粒数和千粒重三要素构成[5],高产育种就是创造和选择能使产量三要素得到最大乘积的遗传变异[6]。北方冬麦区包括北部冬麦区和黄淮冬麦区,是我国小麦主产区[7]。由于北方冬麦区分布范围广,生态条件差异大,不同生态区间小麦产量的构成因素具有不同特点,如北部冬麦区小麦产量的提高与穗粒数和千粒重提高密切相关[8];黄淮冬麦区北片的山东省和河北省小麦产量提高主要是因为有效穗数和穗粒数的提高[9-10],而黄淮冬麦区南片的河南省小麦产量的提高主要通过提高千粒重的途径实现[11]。目前,对北方冬麦区不同生态区小麦产量相关性状已有较多研究[7-11],但在相同环境下对来自不同生态区的代表性品种进行产量相关性状解析和产量潜力评价则鲜有报道。
小麦幼穗分化进程对穗粒数有重要影响[12],与品种的适应性关系密切。大面积推广的品种一般都具有与其所处生态区光温条件相适应的幼穗分化特点。通过调整品种穗分化时间和节奏可使其能够更好地利用光温资源,从而实现高产,如繁6较同时期其他品种穗分化提前,穗分化持续时间延长,穗粒数增加,其产量显著提高[13]。因此,了解北方冬麦区不同生态区代表性品种幼穗分化的特点,对于合理选配亲本,实现幼穗分化不同阶段的最佳配置,进一步提高该麦区的小麦产量具有重要意义。
本研究以北方冬麦区10个大面积推广的代表性品种为材料,对其产量相关性状和幼穗分化特点进行研究和分析,以明确该麦区不同生态区代表性小麦品种的产量形成特点和幼穗分化进程的差异,为高产育种提供理论依据。
1 材料与方法
1.1 试验材料
10个供试小麦品种均来自北方冬麦区(表1),包括黄淮冬麦区南片6个品种,其中,矮抗58是该麦区种植面积最大的品种之一;周麦18和偃展4110分别是该麦区国家冬水组和春水组区域试验的对照品种;周麦16不仅丰产性突出,而且是该麦区最重要的骨干亲本之一;小偃22和淮麦20分别是陕西省关中灌区的当家品种和江苏、安徽淮北麦区的大面积推广品种,也是这两个地区小麦品种区域试验的对照品种。黄淮冬麦区北片品种3个,其中,济麦22是20世纪80年代以来我国种植面积最大的冬小麦品种;烟农19是审定省份最多、种植地区跨度最大的品种,覆盖黄淮冬麦区南片、黄淮冬麦区北片和北部冬麦区3个生态区,先后通过安徽、江苏、山东、山西、河南、北京6个省(市)审(认)定[14];石4185年最大种植面积达到5.71×105hm2(2003年),曾是冀中南水地组和国家黄淮冬麦区北片水地组区域试验的对照品种。轮选987是北部冬麦区的大面积推广品种。

表1 供试材料基本信息Table 1 General information of wheat cultivars tested
*:数据来自国家农业技术推广服务中心。
*:The statistic data originated from the National Agricultural Technology Extension and Service Center.SYHWZ:Southern part of the Yellow and Huang River Valley Winter Wheat Zone;NYHWZ:Northern part of the Yellow and Huang River Valley Winter Wheat Zone;NWWZ:Northern Winter Wheat Zone.
1.2 试验设计与田间管理
2013-2014年,试验在河南省新乡市新乡县七里营镇的中国农业科学院作物科学研究所新乡试验站(北纬35°09′,东经113°48′)进行。采取随机区组设计,4次重复,前3次重复用于产量及其相关性状表型评价,重复4用于幼穗观察。6行区,行长9.0 m,行距20.0 cm,小区间距40.0 cm,小区面积13.5 m2。使用防倒伏网,网眼20 cm2,出苗后将网覆盖在地面上,以后随着植株的生长逐渐抬高网子,使网眼始终处于植株中上部,防止发生倒伏。试验地周围设1.0 m宽的保护行。2013年10月11日播种,按基本苗2.4 × 106株·hm-2计算播种量。播种前施复合肥,折合N 200 kg·hm-2,P2O5191 kg·hm-2,K2O 41 kg·hm-2。拔节期追施尿素,折合纯N 104 kg·hm-2。浇越冬水、拔节水和灌浆水。及时采取药剂防治,避免病虫草害发生。
1.3 测定项目与方法
成熟期每小区随机取10株小麦测量株高;每小区选取整齐均匀的1.0 m双行计数有效穗数;每小区随机选取20个穗测穗粒数;各小区单独机收、脱粒,自然晾干测产量、千粒重。有效分蘖率(成穗率)=有效穗数/最高总茎数 × 100%。
1.4 幼穗分化观察
幼穗分化观察参考文献[15]。幼穗分化形成8个时期(伸长期、单棱期、二棱期、护颖分化期、小花分化期、雌雄蕊分化期、药隔形成期和四分体形成期)的具体划分方法参考文献[16]。幼苗越冬期开始利用体视显微镜(Olympus SZH-ILLD,日本东京)进行幼穗分化观察,返青期后每3 d观察一次,直至旗叶完全展开,整个过程共取样13次,持续时间从2014年1月11日- 4月9日。每个品种每次观察5个主茎,以至少4个主茎穗中部的小穗或者小穗基部两侧的小花到达某一时期来确定具体幼穗分化时期。记载叶龄。
1.5 春化基因和光周期基因分子标记检测
利用DNA快速提取试剂盒(北京天根生化)提取小麦叶片基因组DNA。春化基因主要检测基因座包括 Vrn-A1[17]、 Vrn-B1[17]、 Vrn-D1[15,18]和 Vrn-B3[19],光周期基因检测主要基因座为 Ppd-D1[20]。PCR反应在Biometra Thermocycler(Germany)上进行,总反应体系为20 μL,包括2×TaqPCR Master Mix 10 μL,正反引物(10 μ mol·L-1)各1 μL,DNA模板(50~100 ng·μL-1)2 μL。PCR反应程序同所用引物相应的文献。PCR扩增产物用2%琼脂糖凝胶电泳分离,EB染色后用凝胶成像系统观察电泳结果,读取基因型。
1.6 数据分析
利用SAS 9.2对产量及其相关性状进行方差分析和相关分析。多重比较采用LSD法。
2 结果与分析
2.1 产量及其相关性状分析
由表2可知,8个被测指标在10个品种间存在不同程度差异。就抽穗期和成熟期而言,轮选987最晚,与其他品种差异显著;其次是烟农19和济麦22,石4185和黄淮南片大部分品种相对较早。轮选987、烟农19、淮麦20和小偃22的株高均大于90 cm,其中,轮选987最高,达到97.8 cm,显著高于其他9个品种;偃展4110、济麦22、周麦18和石4185株高85 cm左右;矮抗58和周麦16的株高(75 cm左右)显著低于其他品种。在产量构成三要素上,有效穗数最高的是石4185,达到810.0·m-2,其次为济麦22(797.6·m-2)和轮选987(772.6·m-2),最少的为周麦16(585.3·m-2);穗粒数以济麦22、烟农19最多(35.4),偃展4110最少,与前二者差异显著;周麦16(51.7 g)和周麦18(50.0 g)的千粒重显著高于其他品种。不同生态区间品种的产量没有明显差异。产量达到9 000 kg·hm-2以上的品种有5个,包括小偃22、矮抗58、周麦18、济麦22、轮选987,其中,济麦22和小偃22显著高于其他品种,其余5个品种产量也均达到8 000 kg·hm-2以上,偃展4110的产量显著低于其他品种。

表2 供试小麦品种的产量及其相关性状Table 2 Yield and yield related traits of the tested wheat cultivars
同列数据后不同字母表示品种间差异显著(P<0.05)。图1同。
Different letters following values mean significant difference among cultivars at 0.05 level.HD:Heading period;MD:Maturity period;PH:Plant height;SN:Spikes number per square meter;SR:Spike rate of tiller;KN:Kernel number per spike;TKW:Thousand kernel weight;GY:Grain yield per hectare.The same in Fig.1.
2.2 产量及其相关性状的相关分析
由图1可知,除株高外,其他6个性状均与产量呈正相关,相关系数为0.08(千粒重)~0.45(穗粒数)。有效穗数、穗粒数和千粒重与产量的相关系数分别为0.28、0.45和0.08。抽穗期与成熟期呈极显著正相关,相关系数达到0.98(P<0.01);抽穗期和成熟期与株高呈正相关,相关系数分别为0.61和0.55。有效穗数和成穗率呈显著正相关(r= 0.76,P<0.05)。千粒重与有效穗数和成穗率呈负相关,其中,与成穗率呈显著负相关(r= -0.73,P<0.05)。

图1 产量相关性状的相关分析
2.3 不同小麦品种幼穗分化特点
不同生态区间小麦品种幼穗分化进程存在明显差异。黄淮冬麦区南片品种前期发育较快,后期发育较慢;而轮选987和黄淮冬麦区北片特别是山东省的品种前期持续时间长而后期发育较快。观察幼穗分化进程发现,矮抗58、周麦18、偃展4110、小偃22、淮麦20和石4185穗分化较快,这些品种进入二棱期后,其他4个品种轮选987、济麦22、烟农19和周麦16仍处于单棱期,其中,轮选987穗分化最慢(图2A)。偃展4110、小偃22和石4185较早进入护颖分化期,当这些品种接近小花分化期时,其他大部分品种仍处于护颖分化期,轮选987仍处于二棱期(图2B)。相比周麦16、济麦22、烟农19和轮选987,其他品种较早进入雌雄蕊分化期(图2C)和药隔形成期(图2D)。偃展4110和石4185较早到达四分体形成期,轮选987最晚(图2E)。由表3可知,供试10小麦品种间二棱期结束时间最长相差11 d,叶龄相差1.9。在护颖原基分化期,幼穗分化持续时间为3~8 d,叶龄差为0.2~1.6。其中,烟农19在此时期持续时间最短,为3 d;偃展4110和石4185持续时间最长,为8 d。在小花原基分化期,幼穗分化持续时间为4(矮抗58)~8 d(济麦22);叶龄差为0.5(周麦18)~1.3(矮抗58、周麦18、偃展4110、小偃22和烟农19)。在雌雄蕊原基分化期,幼穗分化持续时间为5(轮选987)~8 d(烟农19);叶龄差为0.8(周麦16)~1.8 (小偃22)。在药隔形成期,幼穗分化持续时间为9~12 d;叶龄差为1.3~3.0。

A~E:分别表示2014年1月11日、3月10日、3月19日、3月28日和4月9日取样10个品种幼穗分化情况。品种顺序从左到右依次为矮抗58、周麦18、周麦16、偃展4110、小偃22、淮麦20、济麦22、烟农19、石4185和轮选987。
A-E:Spike differentiation of ten wheat cultivars on Jan.11,Mar.10,Mar.19,Mar.28 and Apr.9,2014,respectively.The order of the ten cultivars from left to right is as follows:Aikang 58,Zhoumai 18,Zhoumai 16,Yanzan 4110,Xiaoyan 22,Huaimai 20,Jimai 22,Yannong 19,Shi 4185 and Lunxuan 987.
图210个供试小麦品种的幼穗分化进程
Fig.2Spikedifferentiationoftenwheatcultivarstested
2.4 春化基因和光周期基因分子标记检测
除了 Vrn-D1,10个供试品种在其他3个春化基因座 Vrn-A1、 Vrn-B1和 Vrn-B3上全部含有隐性等位变异 vrn-A1、 vrn-B1和 vrn-B3(图3)。在 Vrn-D1基因座,小偃22、石4185和偃展4110含有显性等位变异 Vrn-D1,而其他7个品种含有隐性等位变异 vrn-D1(图3);进一步分析发现,小偃22和石4185含有与半冬性生长习性相关的等位变异 Vrn-D1b,而偃展4110含有与春性生长习性相关的等位变异 Vrn-D1a(数据未列出)。对于光周期基因 Ppd-D1,供试品种均含有不敏感等位变异 Ppd-D1a。

表3 10个供试小麦品种幼穗分化各时期开始时间和叶龄Table 3 Beginning time(BT) of spike differentiation at different stages and leaf ages of ten wheat cultivars
GPDS:护颖原基分化期;FPDS:小花原基分化期;SPDS:雌雄蕊原基分化期;APFS:药隔形成期:TFS:四分体形成期;BT:开始时间。
AK58:Aikang 58;ZM18:Zhoumai 18;ZM16:Zhoumai 16;YZ4110:Yanzhan 4110;XY22:Xiaoyan 22;HM20:Huaimai 20;JM22:Jimai 22;YN19:Yannong 19;S4185:Shi 4185;LX987:Lunxuan 987.GPDS:Glume primordium differentiation stage;FPDS:Floret primordium differentiation stage;SPDS:Stamen primordium differentiation stage;APFS:Anther and pistil forming stage;TFS:Tetrad forming stage;BT:Beginning time.
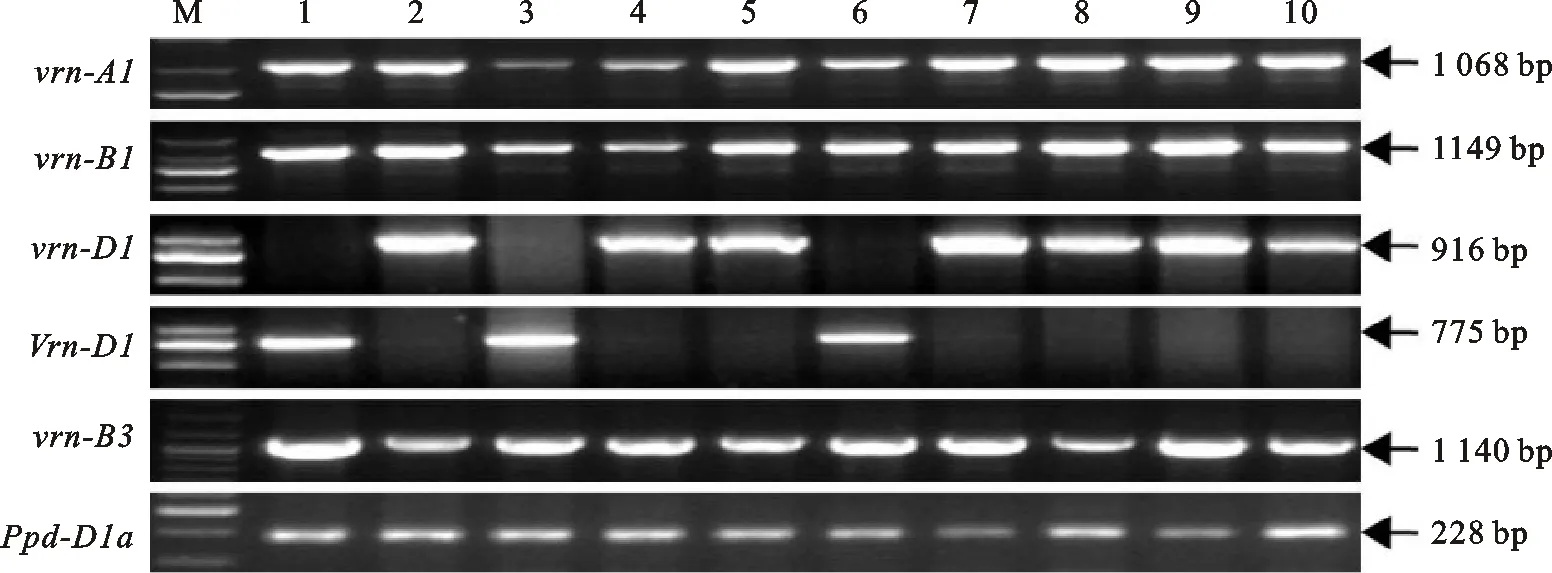
M:DNA标记;1:小偃22;2:矮抗58;3:石4185;4:周麦18;5:周麦16;6:偃展4110;7:济麦22;8:烟农19;9:淮麦20;10:轮选987。
M:DNA marker;1:Xiaoyan 22;2:Aikang 58;3:Shi 4185;4:Zhoumai 18;5:Zhoumai 16;6:Yanzhan 4110;7:Jimai 22;8:Yannong 19;9:Huaimai 20;10:Lunxuan 987.
图310个小麦品种的春化基因和光周期基因
Fig.3Detectionofvernalizationgenesandphotoperiodgenesoftenwheatcultivars
3 讨 论
本研究田间试验是在水肥充足、采取防倒伏和药物防病虫草害条件下进行的,有利于品种产量潜力的充分表现。10个供试品种中,有5个品种产量达到9 000 kg·hm-2,其中,黄淮冬麦区北片的济麦22产量最高,但与黄淮南片的小偃22产量差异不显著,北部冬麦区的轮选987与黄淮冬黄淮冬麦区南片的周麦18和矮抗58产量差异不显著。由此可见,北方冬麦区不同生态区间小麦品种产量潜力差异较小,在适宜的生产条件下均可实现较高的产量。一些品种之所以没能够在河南省大面积种植可能与株高有关。例如,本试验中,小偃22、烟农19、淮麦20和轮选987株高均超过90 cm,如果不采取防倒伏措施,发生倒伏的风险较大。此外,生育期和抗病性也可能限制其跨区种植。
通过对全国各地产量7 500 kg·hm-2小麦产量及其构成因素分析,小麦品种可分为3类:多穗型(750万~900万穗·hm-2),以北部冬麦区品种为主;大穗型(每穗粒重不少于1.3~1.6 g),以南方麦区为主;中间型,以黄淮麦区品种为主[21]。随着生产条件和气候条件的变化,品种类型也发生变化,比如北部冬麦区品种开始由多穗型转向中间型[21]。本研究中,5个产量达到9 000 kg·hm-2品种的有效穗数均属于中间偏多型(600万~750万穗·hm-2),这与前人对黄淮麦区小麦品种产量构成因素分析结果一致[22]。小麦高产的实现需要三要素协调发展,一味地改善某一要素而忽略其他两个要素很难达到高产[23]。例如,石4185在10个品种中有效穗数最多(810.0·m-2),但其千粒重却最低(39.6 g);偃展4110同样具有较高的有效穗数(723.9·m-2),但穗粒数(30.4)最低,结果导致这两个品种产量低于其他品种。
本研究对产量及其构成三要素相关分析结果显示,三者与产量间相关性均未达显著水平,其主要原因可能为:一是本研究供试品种相对较少,尽管他们之间相关系数比较大,但不显著,比如产量与穗粒数间;二是虽然不同品种间产量构成要素相同,但三要素的侧重点不同,所有品种放在一起进行相关分析会相互抵消,导致相关性不显著。
冬性和半冬性小麦品种必须经历一定时间的低温春化处理才能够完成茎生长锥到幼穗分化的转变[16]。春化基因检测结果表明,偃展4110为春性,石4185和小偃22为半冬性,而其他7个品种为冬性。幼穗分化观察发现,相比冬性品种,春性品种偃展4110和半冬性品种石4185、小偃22单棱期持续时间明显缩短。尽管其他7个品种均为冬性,但这些品种间的幼穗分化进程不尽相同,冬性越强的品种单棱期持续的时间越长[16]。小麦穗分化各个时期均受光照长度的控制[24]。对光周期主要基因 Ppd-D1不同等位变异检测发现,供试的品种均含有光周期不敏感型等位变异 Ppd-D1a,表明10个品种在 Ppd-D1基因座上没有差异。品种间在光周期基因座 Ppd-A1和 Ppd-B1上是否存在不同的等位基因有待进一步研究。
本研究中,相比黄淮冬麦区南片的品种,北片的小麦品种普遍前期发育慢,而后期(小花分化期-四分体形成期)发育快,与前人研究结论一致[16]。在对抽穗期和成熟期进行改良时,应注意品种间穗分化不同阶段的互补。例如,烟农19在进入小花原基分化期以前,穗分化速度较慢,进入小花原基分化期以后,穗分化速度加快,但进入药隔形成期和四分体形成期时,穗分化速度变慢,而石4185在进入雌雄蕊原基分化期、药隔形成期以及四分体形成期时,穗分化速度较快。如果要对烟农19的成熟期进行改良,使其成熟期适当提早,通过与石4185进行杂交,后代比较容易选择出比烟农19成熟期和抽穗期更早的材料。
不同冬春性类型(春性、冬性和半冬性)小麦品种间抗寒能力不尽相同,而且同一生长习性类型品种间也存在明显差异[16,25-26]。多年生产实践表明,黄淮北片育成品种(济麦22和烟农19)抗春季低温冻害的能力较强,而黄淮南片育成品种周麦18和周麦16抗春季低温冻害的能力较弱。一般认为,幼穗分化进入雌雄蕊分化和药隔形成期,品种对春季低温最敏感[27]。本研究中,周麦18和周麦16进入这两个分化时期的时间与济麦22和烟农19基本一致。由此可见,单纯从穗分化进程上很难解释黄淮北片品种与黄淮南片品种抗春季冻害能力的差异。品种间抗春季冻害能力的差异可能存在其他的遗传机制。
