1 株携带mcr-1的食品动物源多药耐药奇异变形杆菌耐药性状分析
2018-07-24张德福赵禹宗刘雪飞杨立娜白凤翎周冬生李钰金励建荣
张德福,安 慧,张 健,赵禹宗,张 蕊,刘雪飞,杨立娜,白凤翎,李 春,周冬生,李钰金,殷 喆,励建荣,*
(1.渤海大学食品科学与工程学院,生鲜农产品贮藏加工及安全控制技术国家地方联合工程研究中心,辽宁省食品安全重点实验室,辽宁省高等学校生鲜食品产业技术研究院,辽宁 锦州 121013;2.军事科学院军事医学研究院微生物流行病研究所,病原微生物生物安全国家重点实验室,北京 100071;3.渤海大学数理学院,辽宁 锦州 121013;4.荣成泰祥食品股份有限公司,山东 荣成 264309)
耐药细菌的出现是21世纪人类健康面临的最严峻的挑战之一,而食物链在人接触耐药细菌的途径中居重要地位[1-3],如人类大多数耐药沙门菌感染就是因为食用了污染的动物性食品而引起的[3-7]。在美国,每年可能有一百万起食源性沙门菌喹诺酮耐药性病例,其中有上千起死亡病例,几乎都源于动物性食品[8]。人和动物的肠道是耐药菌的储存库,而且由于畜禽集约化养殖的迅猛发展,畜禽粪便经常没有任何净化处理任意排放,污染了土壤和水源[3,7,9-10];粪便经雨水冲刷、地表径流等最终进入湖泊和海洋[11-12],造成蔬菜[13-14]、水产品[13,15-18]、畜禽等动物性食品[6,9,19]及地下水[7,10-11]等出现耐药菌。耐药菌最终通过被污染的作物、耐药菌感染的畜禽肉和蔬菜、暴露在耐药菌污染的水中的水产品等途径进入人体,从而引起人类的感染及耐药菌株的传播[3]。
奇异变形杆菌(Proteus mirabilis)是肠杆菌科、变形杆菌属的一种革兰氏阴性条件致病菌,广泛存在于水、土壤、腐败的有机物以及人和动物的肠道中,与临床关系较密切,能够引起慢性中耳炎、创伤感染等继发感染,也可引起膀胱炎、婴儿腹泻、食物中毒等,是仅次于大肠埃希菌的可引起泌尿系统感染的主要病原菌[20]。奇异变形杆菌主要污染动物性食品,特别是肉类,受污染的食品在一定条件下会引起人的感染。我国公布的细菌性食物中毒病原中,奇异变形杆菌仅次于副溶血弧菌(Vibrio parahaemolyticus)、沙门菌(Salmonella)和金黄色葡萄球菌(Staphylococcus aureus),是引起食物中毒的第四大主要病原菌[21]。
多黏菌素是从多黏杆菌培养液中提取的一种多肽类抗菌药物,主要为多黏菌素B和多黏菌素E(黏菌素)两种。由于多黏菌素具有肾毒性和神经毒性及广谱抗菌药物的研发与使用,在临床上已较少使用;但是由于20世纪90年代末多药耐药革兰氏阴性菌的出现,尤其是耐碳青霉烯类肠杆菌的出现,多黏菌素又被重新应用于这类细菌的治疗,并被视为对抗多药耐药革兰氏阴性细菌感染的最后一道防线[22]。因此,多黏菌素耐药问题受到了国际上的广泛关注。2015年11月,华南农业大学刘健华教授和中国农业大学沈建忠院士的团队首次从猪体内分离得到一株对黏菌素耐药的大肠杆菌SHP45,进而报道了肠杆菌科细菌质粒介导的多黏菌素耐药基因mcr-1对多黏菌素的耐药机理,该基因是国际上首次发现的质粒介导的黏菌素耐药基因[23]。mcr-1耐药基因多出现在大肠埃希菌中,此外,肺炎克雷伯菌(Klebsiella pneumoniae)、产气肠杆菌(Enterobacter aerogenes)、阴沟肠杆菌(E. cloacae)、沙门菌(Salmonella)、克吕沃菌(Kluyvera ascorbata)、阪崎克罗诺杆菌(Cronobacter sakazakii)等肠杆菌科细菌也有关于mcr-1耐药基因的报道[22],但尚鲜有研究表明奇异变形杆菌携带mcr-1耐药基因的研究报道。本课题组在对辽宁锦州附近的养鸡场、养猪场、屠宰场等地的食品动物源性多药耐药菌的调查中,在某养鸡场分离得到一株携带mcr-1等耐药基因的广泛耐药分离株F160211,经鉴定为奇异变形杆菌,并对该菌的耐药谱、耐药基因、携带耐药基因的质粒复制类型等进行了进一步的研究。
1 材料与方法
1.1 材料与试剂
麦康凯(MacConkey,MAC)琼脂培养基、伊红美蓝(eosin methylene blue,EMB)琼脂培养基、牛脑心浸液(bovine brain heart infusion,BHI)培养基、Mueller-Hinton肉汤(MH)琼脂培养基 美国BD公司;革兰氏阴性需氧菌药敏检测板 上海星佰生物技术有限公司;Taq-聚合酶链式反应(polymerase chain reaction,PCR)Master Mix、细菌基因组DNA提取试剂盒 生工生物工程(上海)股份有限公司;琼脂糖 美国Sigma公司;Goldview核酸染料 北京博迈德公司;2000 DNA Marker 北京鸿跃公司;甘油 美国MP Biomedicals公司;所有引物全部由生工生物工程(上海)股份有限公司合成。
药敏纸片:头孢唑啉(30 μg/片)、头孢他啶(10 μg/片)、亚胺培南(10 μg/片)、环丙沙星(5 μg/片)、阿米卡星(30 μg/片)、阿奇霉素(15 μ g/片)、四环素(1 0 μ g/片)、磷霉素(200 μg/片)、氯霉素(30 μg/片)、复方新诺明(25 μg/片)、替加环素(15 μg/片)和多黏菌素E(10 μg/片) 英国Oxoid公司。
1.2 仪器与设备
DYY-8C型电泳仪 北京市六一仪器厂;LRH系列生化培养箱 上海一恒科学仪器有限公司;Mastercycler pro梯度PCR仪、5424R冷冻离心机 德国Eppendorf公司;Cheimd Doc XRS型凝胶成像仪 美国Bio-Rad公司;NanoDrop 2000微量分光光度计、II级生物安全柜、纸片分配器 美国Thermo公司;比浊仪 法国梅里埃公司;HWS恒温恒湿培养箱 宁波市科技园区新江南仪器有限公司。
1.3 方法
1.3.1 细菌的分离与纯化
自养鸡场采集病鸡、自屠宰场采集血液样品、自养猪场用棉签蘸取粪便(或者肛门拭子)和鼻拭样品,样品放入采样管或采样袋中,编号后放入冰盒内带回实验室。将病鸡解剖后取发病部位组织约50 g,匀浆后取20 μL涂布MAC琼脂平板,猪粪便直接涂板,抗凝血取100 μL涂板。平板37 ℃培养过夜后挑取单克隆划线接种至EMB琼脂平板上37 ℃培养24 h,再挑单克隆菌落涂布BHI琼脂平板上37 ℃培养12 h左右,刮下菌苔,置于含30%甘油的BHI肉汤中-80℃保存备用。上述培养基均添加200 μg/mL氨苄西林。
1.3.2 细菌DNA模板的制备
根据细菌基因组DNA提取试剂盒说明书提取细菌基因组DNA,稀释至质量浓度为10 ng/μL备用。
1.3.3 16S rRNA基因扩增
以保存的细菌基因组DNA为模板,以参考文献[24]中的27F/1492R引物和反应条件扩增细菌16S rRNA基因。PCR产物经琼脂糖凝胶电泳观察条带大小无误后送到生工生物工程(上海)股份有限公司测序。
1.3.4 纸片扩散法鉴定耐药谱
参照美国临床和实验室标准协会推荐方法[25]及SN/T 1944—2016 《动物及其制品中细菌耐药性的测定 纸片扩散法》鉴定细菌的耐药谱。
1.3.5 最小抑菌浓度(minimum inhibitory concentration,MIC)的测定
采用肉汤稀释法,根据试剂盒说明书,挑取单克隆菌落用0.45%灭菌的生理盐水2~3 mL配制成0.5麦氏浊度的菌悬液后取60 μL加入12 mL BHI培养基摇匀,加入至革兰氏阴性需氧菌药敏检测板中,放置37 ℃恒温培养16~20 h观察结果。
1.3.6 耐药基因筛查
以实验室建立的方法,用PCR筛查超广谱β-内酰胺酶、碳青霉烯酶和喹诺酮类、大环内酯类、多黏菌素类等常见抗菌药物的耐药基因。超广谱β-内酰胺酶、碳青霉烯酶扩增引物等见参考文献[26],其他引物序列与退火温度等见表1。
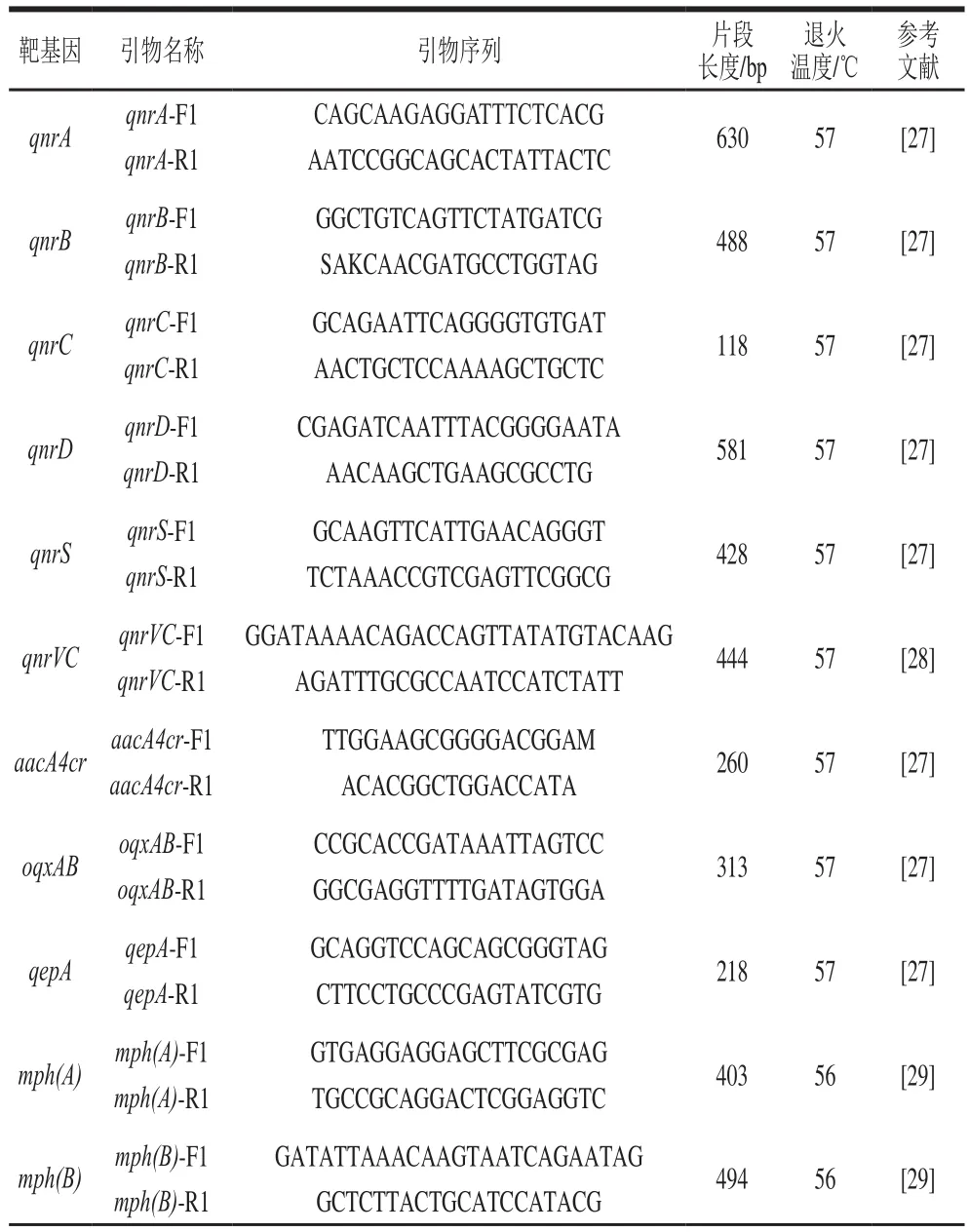
表1 耐药基因筛查用引物Table1 Primers used for drug-resistant gene screening
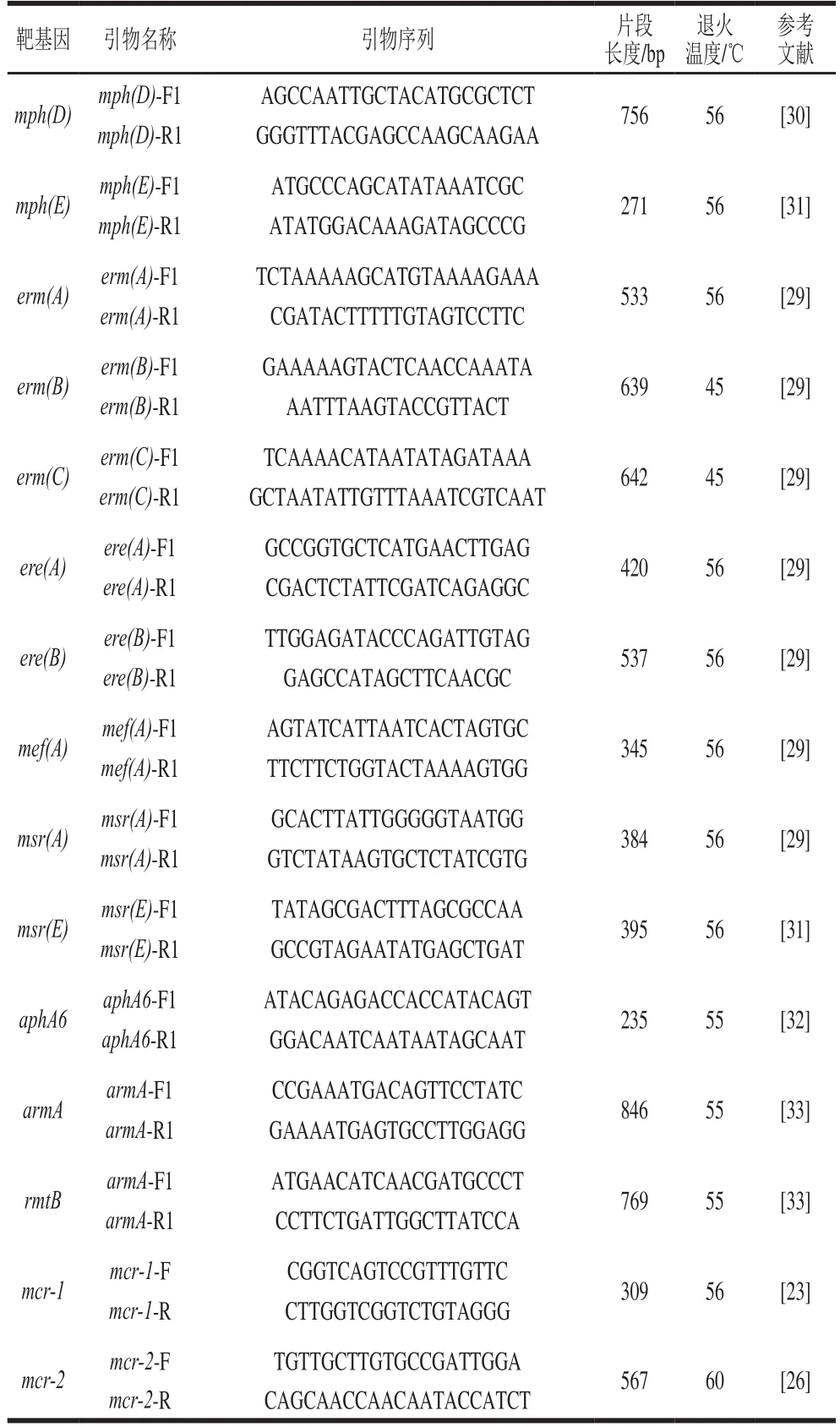
续表1
1.3.7 质粒复制类型分析
根据p H N S H P4 5序列(G e n B a n k登录号:KP347127)的复制起始蛋白基因repA和质粒分离蛋白parA基因设计引物进行扩增,检测携带mcr-1的质粒复制类型。PCR程序为:95 ℃预变性5 min;94 ℃变性40 s,58 ℃退火40 s,72 ℃延伸1 min,30 个循环;最终72 ℃延伸5 min。引物序列见表2。
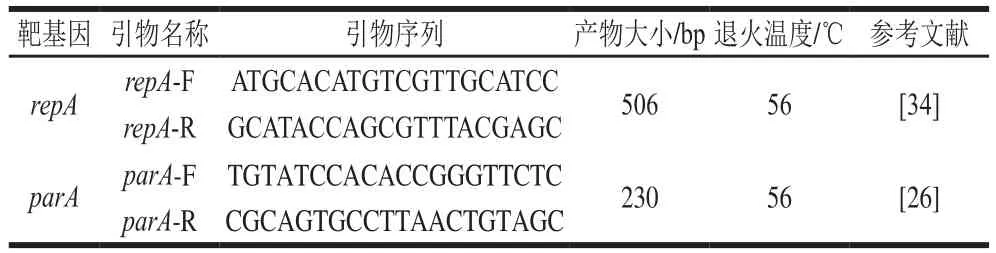
表2 质粒复制类型PCR检测用引物Table2 Primers used for detection of plasmid replicon type
2 结果与分析
2.1 细菌的分离与纯化
课题组在辽宁省内多家养鸡场、养猪场、屠宰场等共采集样品500余份,分离到多药耐药菌361 株,其中一株编号为F160211的菌株初步鉴定为对多黏菌素等药物耐药,因此进行了进一步的研究。
2.2 16S rRNA菌种扩增结果
以细菌F160211的基因组DNA为模板,以27F/1492R引物扩增16S rRNA基因后的测序结果进行BLAST分析表明,分离株F160211为奇异变形杆菌。
2.3 纸片扩散法鉴定耐药谱
纸片扩散法对奇异变形杆菌F160211的耐药谱鉴定表明(表3),该菌对头孢唑啉、头孢他啶、环丙沙星、阿米卡星、阿奇霉素、四环素、磷霉素、氯霉素、复方新诺明和多黏菌素耐药,对亚胺培南和替加环素敏感。

表3 纸片扩散法鉴定奇异变形杆菌F160211耐药谱Table3 Drug-resistance pro fi le of P. mirabilis F160211 using disk-based test
2.4 肉汤稀释法测MIC结果

表4 奇异变形杆菌F160211的MICTable4 MIC of P. mirabilis F160211
利用肉汤稀释法对奇异变形杆菌F160211的耐药情况进一步详细分析(表4),结果表明,该菌对青霉素类、第1代和第2代头孢菌素类、大环内酯类、氨基糖苷类、酰胺醇类、四环素类、磺胺类、利福霉素类和黏菌素类等耐药,对呋喃类、多肽类、喹诺酮类中介耐药,对第3代和第4代头孢菌素类、单环β-内酰胺类、碳青霉烯类、甘氨酰环素类敏感。
2.5 耐药基因筛查结果
以文献报道的、实验室已合成的多种耐药基因引物对奇异变形杆菌F160211的基因组DNA进行PCR扩增,结果表明,F160211携带blaCTX-M-1、blaCTX-M-9、blaTEM、blaOXA-1、mph(A)、qnrS和mcr-1等多种耐药基因(图1)。
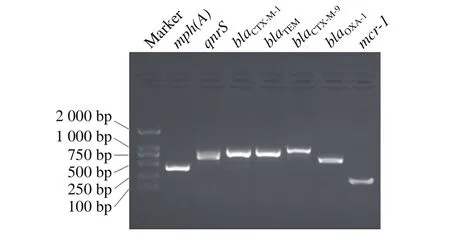
图1 奇异变形杆菌F160211耐药基因PCR筛查Fig.1 PCR screening of drug-resistance genes from P. mirabilis F160211
2.6 质粒复制类型分析
通过对奇异变形杆菌F160211携带mcr-1基因的质粒repA和parA基因进行PCR验证结果表明,养殖场分离得到的分离株奇异变形杆菌F160211的mcr-1基因存在于pHNSHP45类似的IncI2型质粒上(图2)。
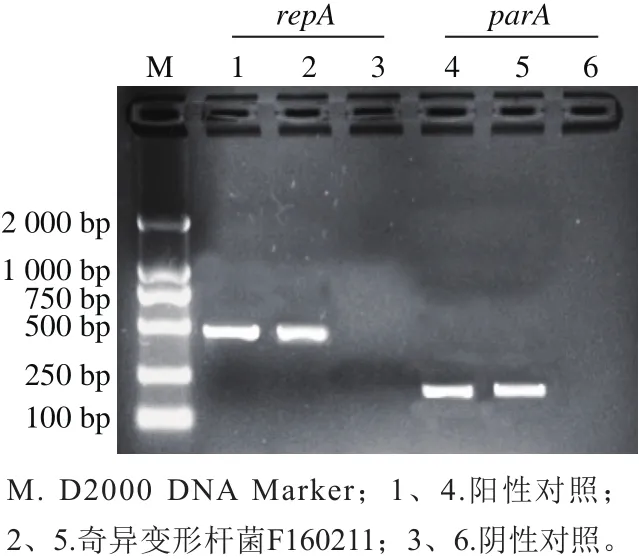
图2 质粒复制类型PCR检测结果Fig.2 PCR detection of plasmid replicon type
3 讨 论
MIC和耐药基因PCR扩增结果表明,分离自病鸡肉中的奇异变形杆菌F160211为多药耐药株,携带有多种耐药基因,耐药性非常严重,除了对第3代和第4代头孢菌素类、单环β-内酰胺类、碳青霉烯类、甘氨酰环素类敏感外,对其他药物均为耐药或中介耐药;且其耐药基因位于质粒上,具有水平转移给其他细菌并播散开来的风险,给消费者的食品安全和身体健康带来隐患。
多黏菌素对大多数革兰氏阴性菌具有较好的杀菌作用,一直被养殖业者广泛应用于促进动物生长和疾病的预治等;在临床上用作对碳青霉烯类耐药的肺炎克雷伯菌、鲍曼不动杆菌和铜绿假单胞菌等革兰氏阴性多药耐药菌治疗的最后一道防线;虽然也有耐药菌出现,但一般都是染色体介导的,其耐药基因的传播也是通过病人之间的克隆株的传播,耐药率较低,危害较小[35]。但Liu Yiyun等[23]在鸡、猪、零售的肉类和病人体内发现了质粒携带的、可在细菌间接合转移的多黏菌素耐药基因mcr-1,且该质粒在鸡肉中存在的比例高达28%,引起世界范围的震惊。
本研究中,奇异变形杆菌F160211携带有mcr-1耐药基因,但其耐药性为敏感,这也与Liu Yiyun[23]、易灵娴[22]、Skov[36]等报道mcr-1介导宿主菌低水平的黏菌素耐药相符;携带mcr-1的奇异变形杆菌质粒与Liu Yiyun等[23]报道的属于同一类型不相容性质粒,宿主范围也由报道的大肠杆菌、沙门菌等进一步扩大至奇异变形杆菌,这说明该质粒已在不同菌种间发生了水平转移,形势更加严峻。随着时间的推移,如果没有合理的应对措施,将可能有更多的菌种产生mcr-1耐药基因,这无论对食品安全还是临床感染的治疗都将是严峻的挑战,同时也对人类滥用抗菌药物的行为敲响了警钟。
