异育银鲫源嗜水气单胞菌对磺胺类耐药性分析
2018-07-24万夕和王李宝史文军范贤平张建明
乔 毅,沈 辉,万夕和,王李宝,史文军,黎 慧,蒋 葛,范贤平,张建明
( 1.江苏省海洋水产研究所,江苏 南通 226007; 2.江苏省海洋渔业指挥部,江苏 南通 226006 )
异育银鲫(Carassiusauratusgibelio)是中国科学院水生生物研究所的鱼类育种专家于1976—1981年成功选育的一种鲫鱼养殖新种,因其具有生长快、个体大、抗逆性强等优点,成为我国重要的淡水水产养殖鱼类。近些年,随着异育银鲫养殖技术的成熟,养殖户不断提高放养密度,投喂高蛋白优质饵料,获得较好的经济效益。但是在追求高产的同时,异育银鲫的病害频频爆发,鳃出血病[1-3]、黏孢子虫病[4]和常见细菌病害[5-7]给该产业造成巨大的损失,严重制约异育银鲫养殖业的健康发展。
嗜水气单胞菌(Aeromonashydrophila)属弧菌科,气单胞菌属,广泛分布于自然界的各种水体中,是多种水生动物的原发性致病菌[8-9],为条件致病菌,也是典型的人—兽—渔共患病病原菌[10]。目前在淡水养殖过程中,大多数品种均有嗜水气单胞菌引起病害的报道,其主要症状是爆发性出血,由于嗜水气单胞菌的血清型较多,所以感染不同对象表现的症状各异。目前,以抗菌药物治疗水生动物细菌性病害是水产养殖过程中最常用的方法,由于抗菌药物的长时间乱用和滥用,水产病原菌在应对药物反复刺激的压力下形成的耐药性日趋严重,使得水产源病原菌引起的疾病药物治疗效果越来越差。磺胺类药物因其具有抗菌谱广、性质稳定、吸收迅速、使用方便和价格低廉等优点,是水产养殖常见的抗菌药物之一,磺胺类药物通过阻碍细菌RNA的合成达到抑菌作用,可防治多种水产病原菌,而临床中的不合理使用,使得该类药物的耐药性日益突显。
江苏省苏北地区异育银鲫养殖面积超过六万公顷,近些年病害引起该地区异育银鲫的损失高达十余亿元。2015年,本课题研究团队从盐城市大丰地区异育银鲫病样中筛选致病菌株,经鉴定其中嗜水气单胞菌36株。笔者以分离纯化的嗜水气单胞菌为对象,分析菌株对磺胺类药物的敏感性,同时对菌株携带的磺胺类耐药基因进行PCR扩增,旨在为临床用药以及嗜水气单胞菌对磺胺类药物产生耐药的机制作一定参考。
1 材料与方法
1.1 菌株
36株嗜水气单胞菌株,为2015年3—9月大丰发病异育银鲫体内分离得到,依次编号为1、2、3、……、36;嗜水气单胞菌标准菌株ATCC7966购自苏州达麦迪生物医学科技有限公司;大肠杆菌(Escherichiacoli)标准菌株ATCC25922购自杭州滨和微生物试剂有限公司。
1.2 主要试剂
磺胺嘧啶(CAS:68-35-9)、磺胺甲噁唑(CAS:723-46-6)、磺胺异噁唑(CAS:127-69-5)、磺胺二甲基嘧啶(CAS:57-68-1)标准品购自上海阿拉丁生化科技股份有限公司;0.5麦氏单位比浊管和水解酪蛋白琼脂培养基购自杭州滨和微生物试剂有限公司;细菌基因组DNA提取试剂盒(离心柱型)、质粒小提试剂盒(离心柱型)购自天根生化科技(北京)有限公司;Agarose B、Low EEO、SanPrep柱式DNA胶回收试剂盒、DEPC水购自生工生物工程(上海)股份有限公司;大肠杆菌DH5α感受态细胞、Premix TaqTM(TaKaRa TaqTMVersion 2.0 plus dye)、T-Vector pMDTM19 (Simple)、Ribonuclease A (RNase A)等分子生物试剂购自宝生物工程 (大连) 有限公司;药敏纸片自制,含量均为300 μg/片;其他化学试剂均为分析纯。
1.3 菌株药物敏感性试验
在无菌条件下,接种低温冻存的菌株在LB琼脂培养基上复苏细菌。根据美国临床实验室标准化委员会推荐的K-B纸片扩散法,挑取纯培养菌落,悬于3 mL的无菌生理盐水试管中,混匀后调整浊度与0.5麦氏单位比浊管一致。用无菌棉拭子蘸取菌液,均匀涂布整个水解酪蛋白琼脂培养基表面。待平板上的水分被琼脂完全吸收后,用无菌镊子取药敏纸片贴在平板表面,纸片贴上后不可再拿起,用镊子轻压纸片中央,保证纸片与琼脂表面紧密贴合,每张纸片中心间距不少于24 mm,纸片中心距平皿边缘不少于15 mm,在细菌接种后15 min内贴完纸片。将平板倒置,28 ℃培养18~20 h,用游标卡尺准确测量抑菌圈直径。参照标准判读结果(表1)。每批药敏试验必须用大肠杆菌ATCC25922做质控。只有当质控菌株的抑菌圈直径在允许范围内,其他菌株的结果才可以报告。
1.4 Sul基因扩增及分析
按照天根试剂盒操作说明分别提取嗜水气单胞菌基因组DNA和质粒DNA,经RNase A酶去除DNA制备过程中的RNA,-20 ℃保存备用。磺胺类药物耐药基因Sul1、Sul2和Sul3引物参照文献[11]的报道(表2)。引物委托南京金斯瑞生物科技有限公司合成。分别以嗜水气单胞菌基因组DNA和质粒DNA为模板,常规PCR扩增磺胺类耐药基因Sul1、Sul2和Sul3。扩增采用50 μL体系,包括25 μL Premix Taq酶,上、下游引物各2 μL,DEPC处理水19 μL,模板2 μL。3个基因的扩增程序一致:94 ℃ 预变性5 min,94 ℃变性1 min,55 ℃退火1 min,72 ℃延伸1 min,共30个循环,72 ℃终延伸7 min。PCR扩增产物经1.5%琼脂糖凝胶电泳检测,对出现与目的片段大小相符的条带进行切胶回收,回收产物按操作说明与pMDTM19 载体连接后导入工程菌DH5α 中构建重组质粒,提取重组质粒DNA用相应基因引物进行扩增,扩增阳性的质粒外送测序。序列结果利用DNAMAN软件进行序列间相似性分析,再通过美国国立生物技术信息中心基因库中Blast比对,并利用Clustal X 3.0和Mega 6.0软件构建耐药基因系统发育树。
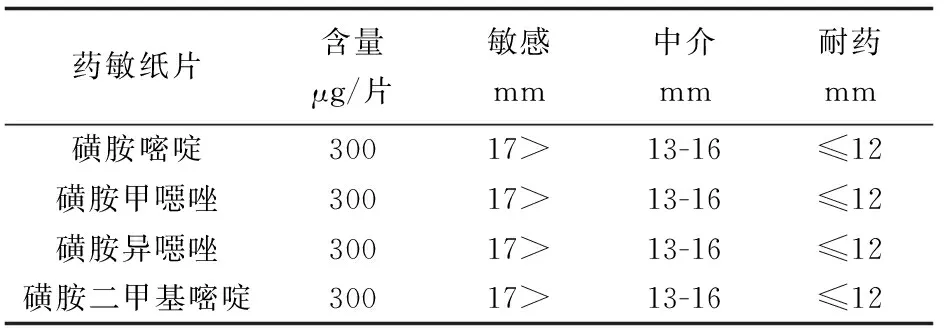
表1 嗜水气单胞菌抑菌圈直径解释标准
2 结 果
2.1 药物敏感性试验结果
采用纸片扩散法和肉汤稀释法测定36株异育银鲫源嗜水气单胞菌对4种磺胺类药物敏感性,参照美国临床和实验室标准协会标准,以纸片周围形成的抑菌圈大小作为评价嗜水气单胞菌药物敏感性的主要依据,结合描述的方法,在进行药物敏感性的最终判定时,对纸片法显示为中介而最小抑菌质量浓度值较大的菌株判定为耐药,对抑菌圈远大于标准而最小抑菌质量浓度值偏大的菌株判定为敏感。其药物敏感性结果见表3。
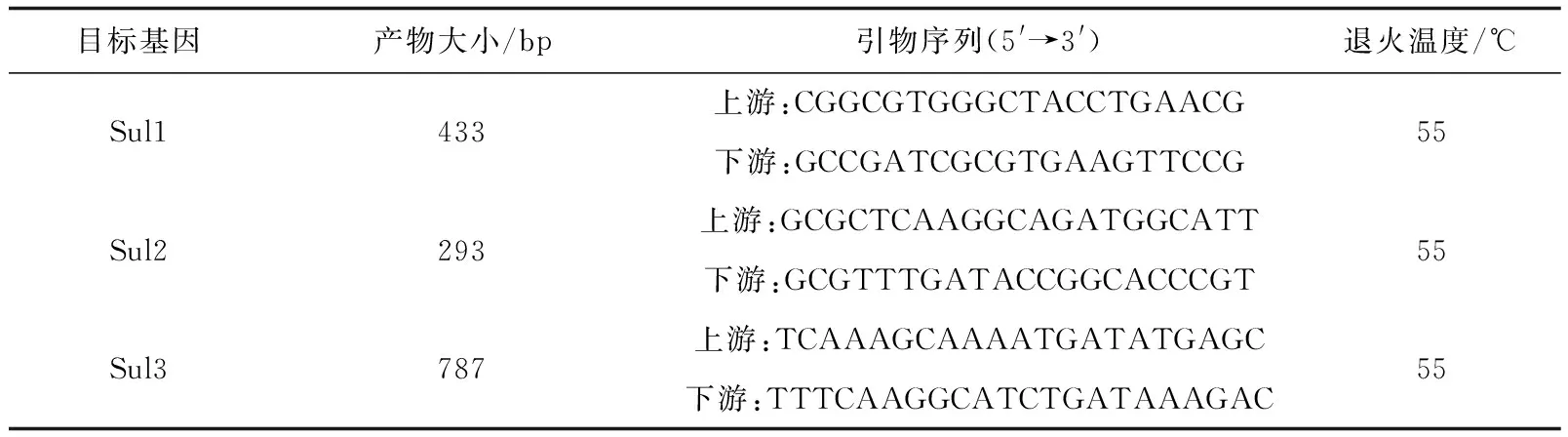
表2 磺胺类耐药基因扩增引物
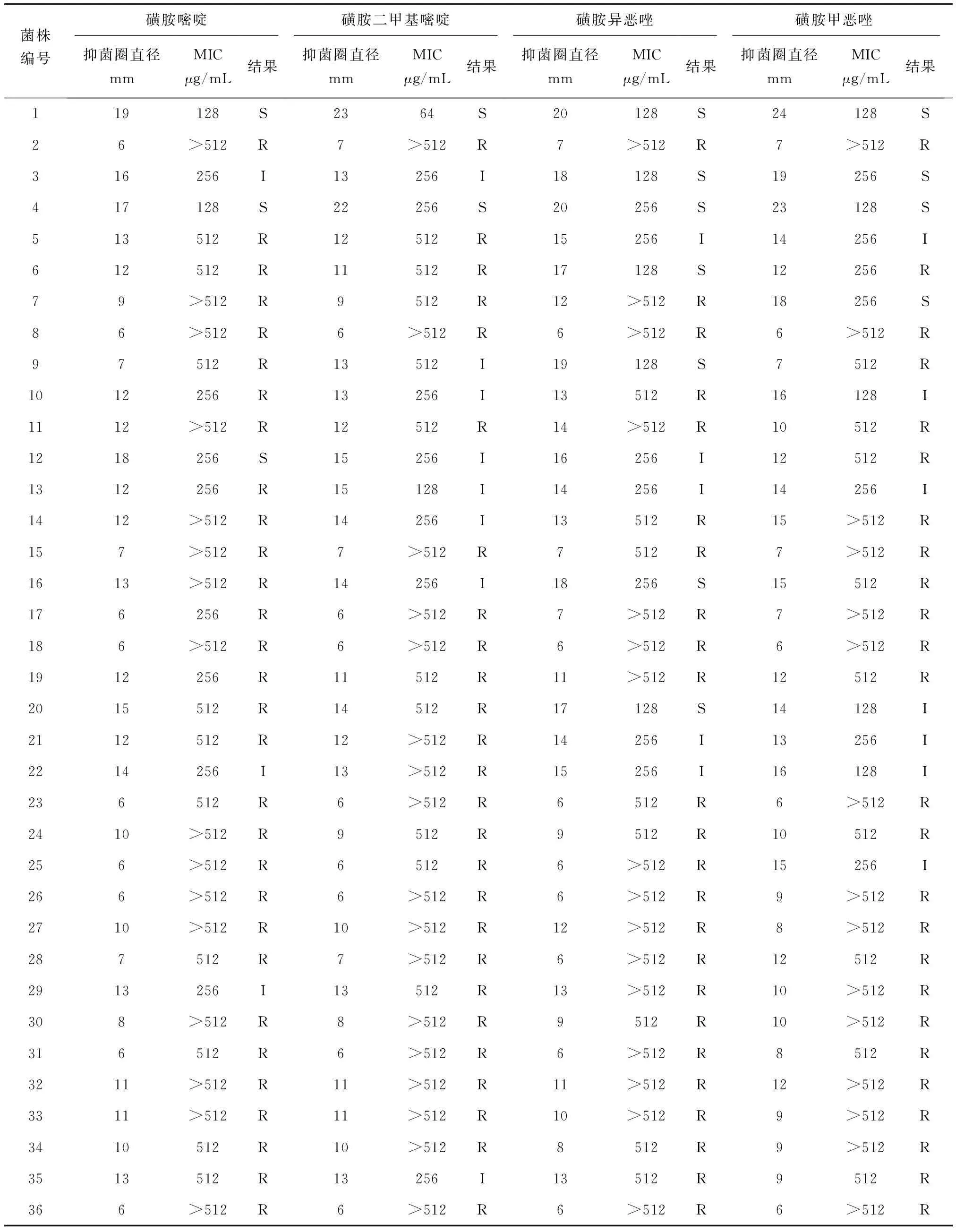
表3 嗜水气单胞菌药物敏感性试验结果
注:R,耐药;I,中介;S,敏感;MIC,最小抑菌质量浓度.
统计发现,36株嗜水气单胞菌对磺胺嘧啶、磺胺二甲嘧啶、磺胺异噁唑和磺胺甲噁唑的耐药率分别为83.34%、72.22%、66.66%和69.45%,其具体耐药菌株号见表4。
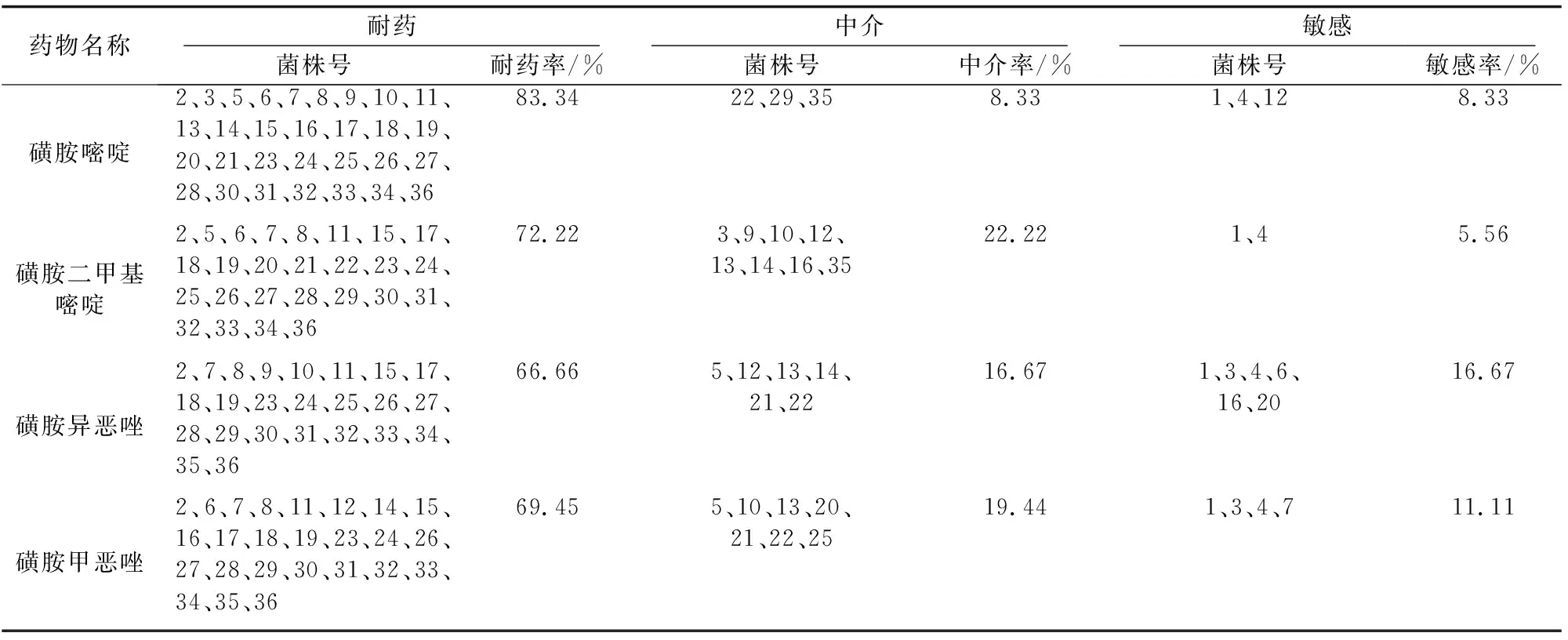
表4 嗜水气单胞菌对磺胺类药物的耐药率、中介率和敏感率
2.2 耐药基因扩增结果
本研究通过常规PCR方法对36株异育银鲫源嗜水气单胞菌磺胺类药物耐药基因进行扩增,其电泳结果见图1~图6,其中Sul1耐药基因在细菌染色体中检测为阳性的有11株,质粒中阳性的有10株;Sul2耐药基因在细菌染色体中检测为阳性的有30株,质粒中阳性的有33株;Sul3耐药基因在36株菌的染色体和质粒中均未检出。

图1 染色体Sul1耐药基因电泳结果
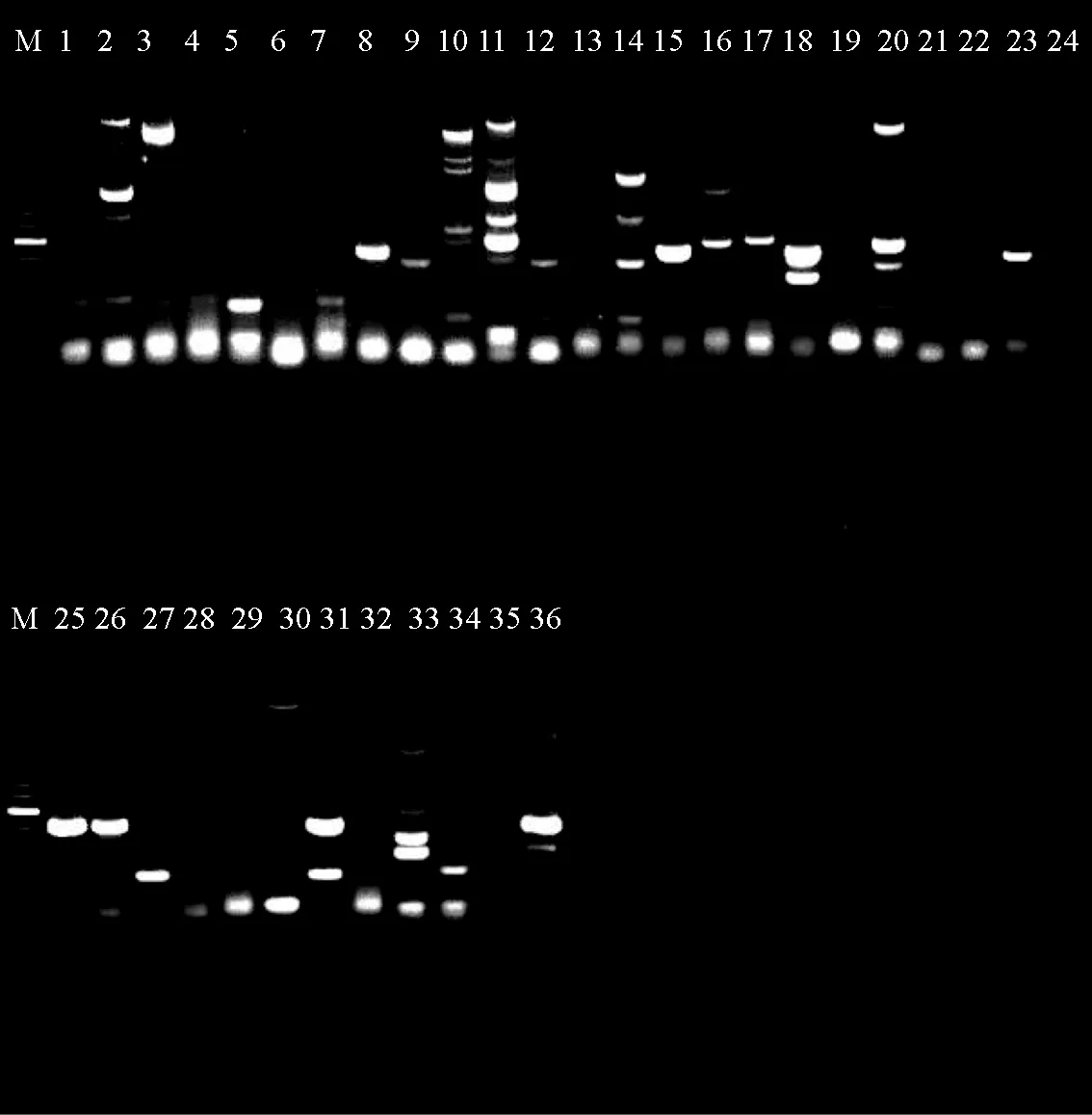
图2 质粒Sul1耐药基因电泳结果

图3 染色体Sul2耐药基因电泳结果

图4 质粒Sul2耐药基因电泳结果
图5 染色体Sul3耐药基因电泳结果

图6 质粒Sul3耐药基因电泳结果
分别统计对4种磺胺类药物耐药的嗜水气单胞菌中耐药基因的携带情况(表6),Sul1耐药基因在4种药物耐药菌的染色体和质粒上携带率显著;Sul2耐药基因在4种药物耐药菌的染色体上携带率为83.33%~88.00%,在质粒上携带率为91.67%~93.33%。
2.3 基因克隆及分析
耐药基因电泳结果除具有目的片段大小的条带外还有其他非特异扩增(图1~图4),本研究将目的片段大小的条带进行切胶回收,通过克隆后外送测序。序列大小与预期结果基本相符,通过DNAMAN软件分别对Sul1和Sul2基因片段进行多序列比对分析,结果显示序列一致性为100%。将序列输入美国国立生物技术信息中心数据库中进行Blast同源性比对,结果显示,Sul1和Sul2基因序列分别与数据库中多个基因片段同源性达到100%。
在GenBank分别下载多个Sul1和Sul2基因,通过Mega 6.0构建系统发育树。结果显示,本研究中的嗜水气单胞菌磺胺类药物耐药基因Sul1与不动杆菌属(Acinetobacter,登录号:EU375649.1)的Sul1基因聚为一支(图7);Sul2耐药基因与假交替单胞菌属(Pseudoalteromonas,登录号:EF492006.1)的Sul2基因聚为一支(图8)。

表5 嗜水气单胞菌耐药基因检测结果
注:“+”示结果阳性,但条带较弱;“++”示结果阳性,条带较亮;“-”示结果阴性.
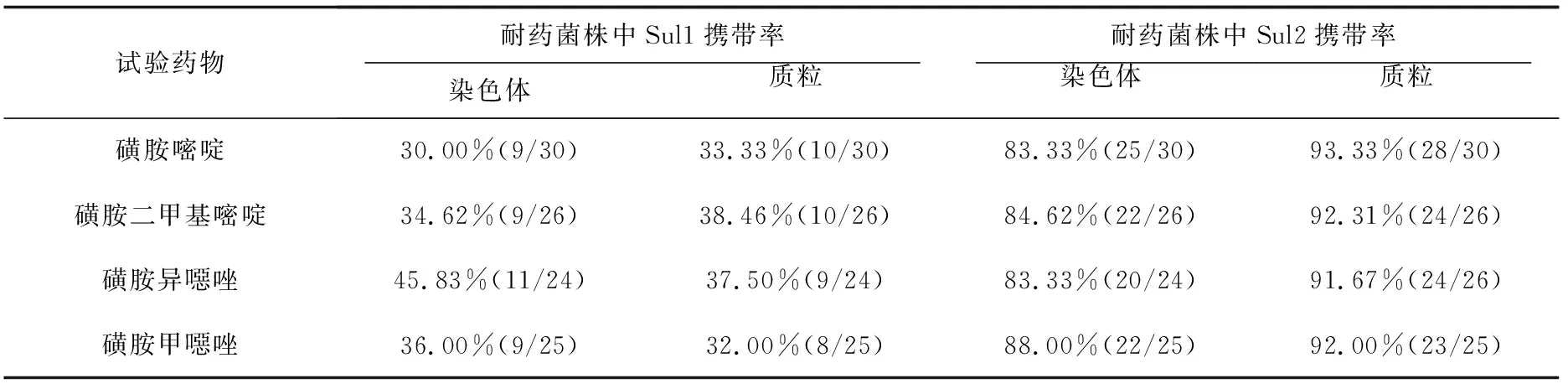
表6 磺胺类药物耐药嗜水气单胞菌染色体和质粒上耐药基因的携带率
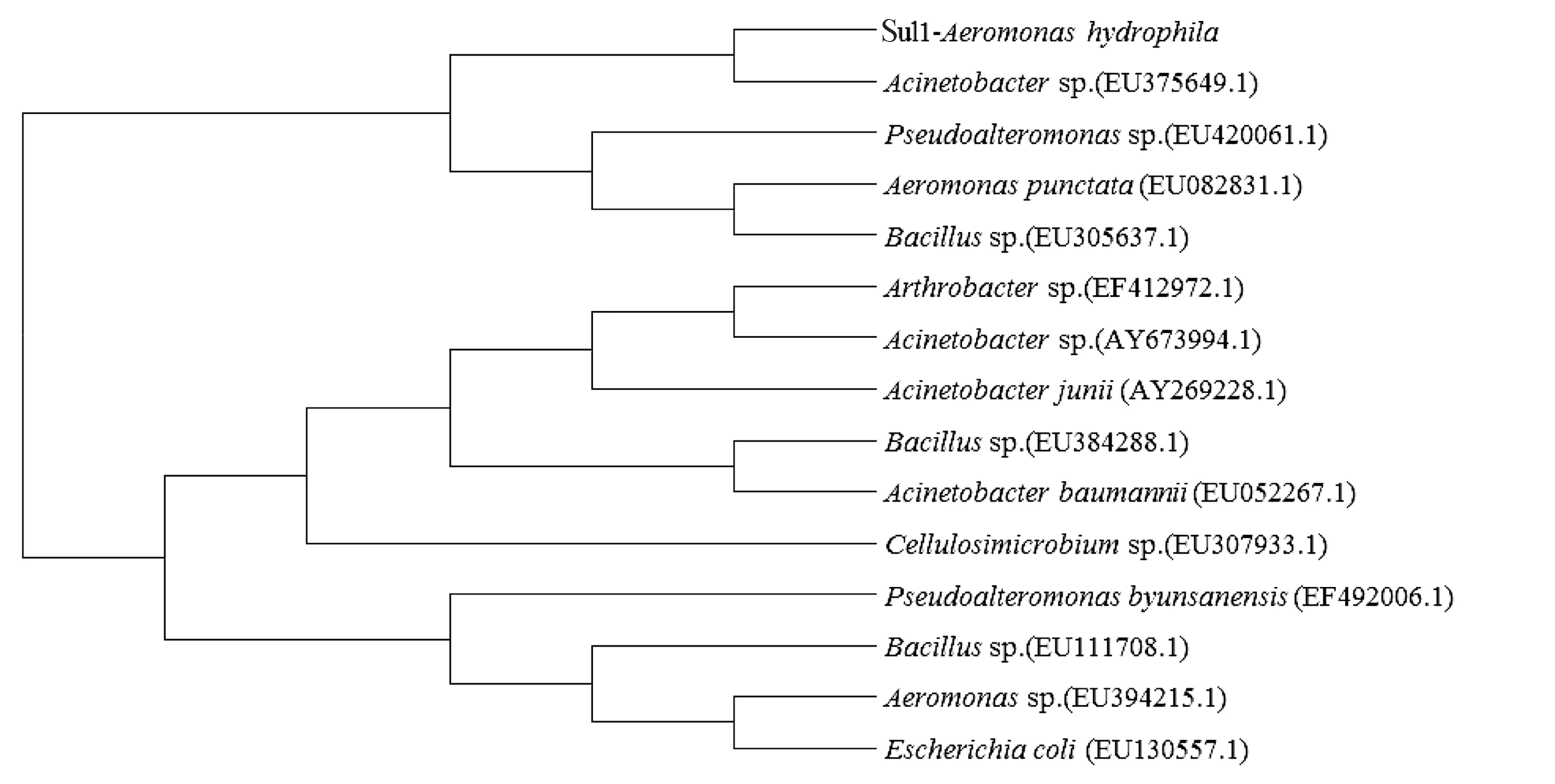
图7 嗜水气单胞菌Sul1耐药基因系统发育树
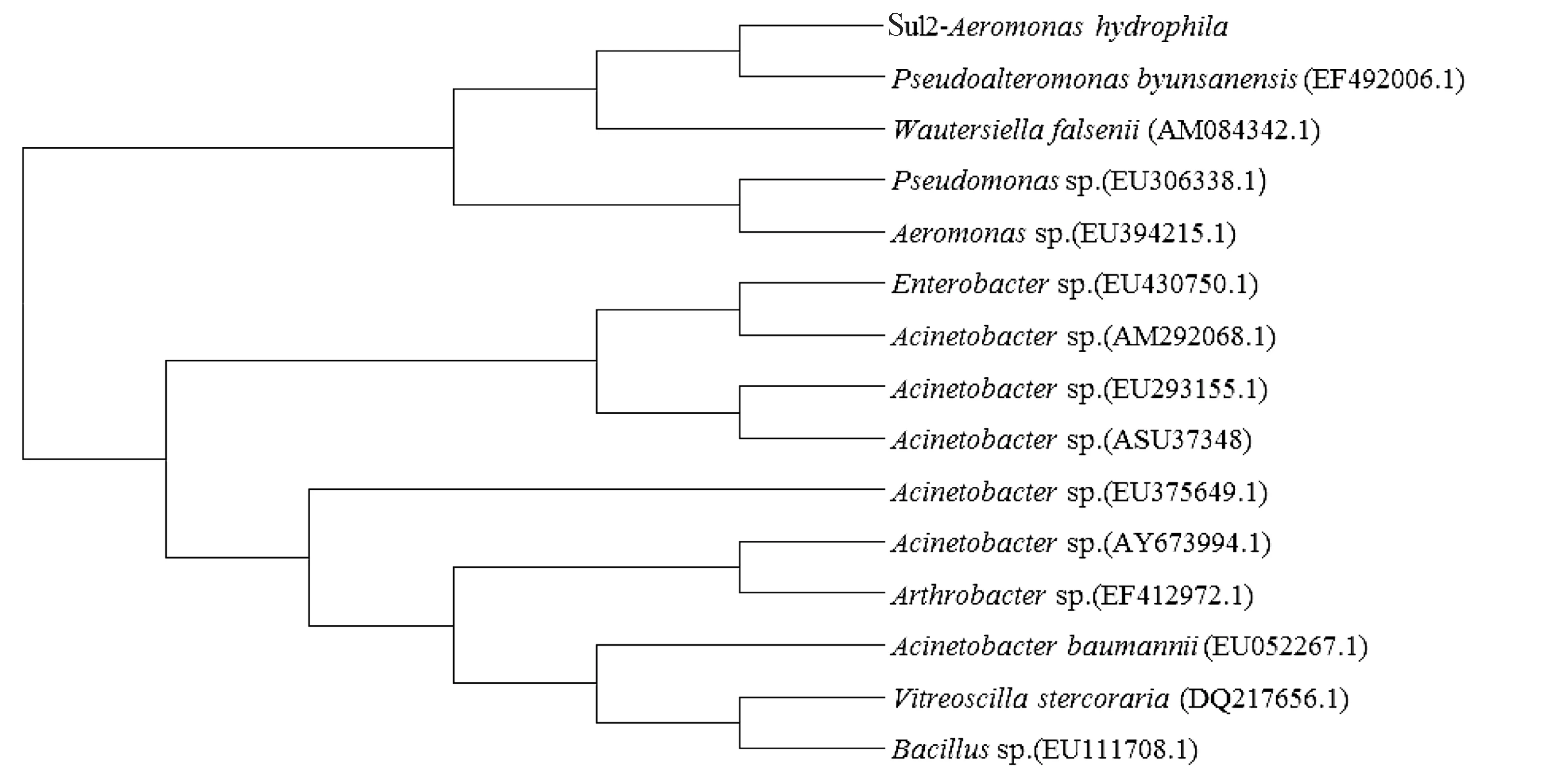
图8 嗜水气单胞菌Sul2耐药基因系统发育树
3 讨 论
采用纸片扩散法和最小抑菌质量浓度法测试了36株异育银鲫源嗜水气单胞菌对4种磺胺类药物的敏感性,2种方法的试验结果并不完全一致,为了能准确反映细菌的药物敏感性,结果参考文献[12]的方法进行判定,以纸片扩散法抑菌圈的大小作为主要依据,辅以最小抑菌质量浓度评价药物敏感性,对于纸片法中介,最小抑菌质量浓度值较大的菌株判定为耐药,对于抑菌圈远大于标准而最小抑菌质量浓度值偏大的菌株判定敏感。试验中最小抑菌质量浓度法的肉汤中磺胺类药物配制的最高质量浓度为512 μg/mL,当最高质量浓度的试管中依然有细菌生长时最小抑菌质量浓度值记为>512 μg/mL。最终试验结果表明36株菌对磺胺嘧啶、磺胺二甲基嘧啶、磺胺甲噁唑和磺胺异噁唑耐药率依次为83.34%、72.22%、66.66%和69.45%,这与彭彬等[13-16]的研究结论相似,由此可见,水产养殖源嗜水气单胞菌对磺胺类药物的耐性达到非常严重的水平。
对磺胺类药物敏感的细菌不能直接利用周围环境中的叶酸,只能利用氨苯甲酸和二氢蝶啶在细菌体内经二氢叶酸合成酶的催化合成二氢叶酸,再经过二氢叶酸还原酶的作用形成四氢叶酸。四氢叶酸的活化型是一碳单位的传递体,在嘌呤和嘧啶核苷酸形成过程中起着重要的传递作用。磺胺类药物的结构跟氨苯甲酸相似,因而可与氨苯甲酸竞争二氢叶酸合成酶,阻碍二氢叶酸的合成,从而影响核酸的合成,抑制细菌的生长繁殖。细菌额外获得二氢叶酸合成酶(Sul基因编码)和二氢叶酸还原酶(dfrA基因编码)编码基因,降低了磺胺类药物对二氢叶酸合成酶的竞争力,同时补充了二氢叶酸还原酶,削弱磺胺类药物增效剂(如甲氧苄啶等)的作用,导致细菌对磺胺类药物出现耐药[17]。其中Sul基因在细菌对磺胺类药物耐药过程中起主要作用,目前认为Sul基因可以分为3个亚型,分别为Sul1[18]、Sul2[19]和Sul3[20],其中Sul1基因由Ⅰ类整合子介导,可在不同菌株间水平传播,Sul2基因既可存在大的质粒上又可由染色体介导,Sul3是Perreten等[20]在大肠杆菌中新发现的一个Sul基因。本研究中在36株嗜水气单胞菌的染色体和质粒中均检测到Sul1和Sul2基因,Sul1基因在染色体和质粒中的检出率分别为30.56%和25.64%,Sul2基因在染色体和质粒中的检出率分别为83.33%和91.67%,Sul3基因在染色体和质粒中均未检出,并且在对4种磺胺类药物耐药嗜水气单胞菌染色体和质粒中都有较高的携带率,可见Sul1和Sul2耐药基因与36株异育银鲫源嗜水气单胞菌磺胺类药物耐药性存在相关性,其中Sul2可能是介导耐药的重要基因。该结果与羊云飞等[21]的研究较为相似,赖海梅等[22]在肉鸡沙门氏菌(Salmonella)中检测到Sul1、Sul2和Sul3的阳性率分别为40%、100%和63.3%,认为Sul2是主要的磺胺类耐药基因,而与Antunes等[23-25]Sul1为磺胺类药物的主要耐药基因的研究结果存在差异。Huerta等[26-27]研究认为耐药基因的产生很有可能是多因子共同影响的结果,Ji等[28]就发现Sul1和Sul3与重金属Cu、Zn和Hg存在显著正相关,本研究的对象、时间和空间的差异可能是导致与其他学者研究结果不同的因素。Hoa等[29]研究表明,大多数Sul基因位于染色体基因上,本研究对嗜水气单胞菌磺胺类耐药基因在染色体DNA和质粒DNA上分布情况做了简单统计,结果发现Sul基因在嗜水气单胞菌的染色体和质粒DNA上分布差异不显著,这与王娜[30]的研究结论一致。磺胺类耐药性与耐药基因及其分布关系复杂,是开展深入研究的一个重要方向。
通过对36株异育银鲫源嗜水气单胞菌磺胺类药物的敏感性研究,发现该类细菌对磺胺类药物的抗性较强,对该类细菌引起的水生动物病害控制形成一定难度。同时,嗜水气单胞菌是一种人—兽—渔共患病菌,这对人类公共卫生、食品安全和生态环境构成严重威胁。另外,耐药基因能够通过质粒和转座子等可移动元件上在同种或不同种细菌间水平传播,因此,关于该地区异育银鲫源嗜水气单胞菌耐药基因的研究应该受到广泛关注。
