濒危植物丰都车前的繁育系统与传粉生物学研究
2018-07-18万海霞邓洪平蒋庆庆
万海霞,邓洪平,何 平,蒋庆庆,刘 钦
西南大学生命科学学院,三峡库区生态环境教育部重点实验室,重庆 400715
丰都车前(Plantagofengdouensis)为车前科(Plantaginaceae)车前属(PlantagoL.)二年生草本植物,是三峡库区特有濒危植物[1- 2]。其野外分布十分狭窄,生境破碎,个体数量稀少。据文献记载,该种仅分布于重庆市忠县、丰都县和巴南区等3个彼此隔离的长江江心岛上[3]。但这3个江心小岛因三峡工程建设、蓄水,陆续被完全淹没,致使丰都车前曾被认为是目前所发现的因三峡工程建设而导致原始生境毁灭、自然种群灭绝的唯一草本植物[3]。
2015年,在重庆市江津支坪镇涂家村附近,长江与綦江交汇处的冲积小岛——大中坝上,发现残留有丰都车前一小种群。经多次踏查、寻访,发现该小岛属于季节性水淹小岛,主要受夏季洪讯的影响,出现一段时间的水位上涨导致水淹;附近工业、农业的发展严重威胁到该野生小种群的生存。目前,对丰都车前保护主要集中在分类、迁地保护、生物学特性观测等方面,而关于其繁殖器官特征、传粉习性以及与季节性水淹的适应等研究未见报道。本研究以丰都车前现存野生种群和移栽种群为材料,综合研究分析其开花习性、繁育系统及传粉适应性,为进一步探究其濒危机制、有效保护这一三峡库区珍稀濒危植物、实现更好的繁殖应用等提供一定理论依据。
1 材料与方法
1.1 实验材料
丰都车前(Plantagofengdouensis)野外植株叶片具3脉,披针形至线状披针形,叶缘羽状锐裂,长为宽的2.5倍以上,无毛,或在裂片弯缺处具短毛;穗状花序;花冠淡黄、红粉色,裂片狭三角形;花药红黄色;蒴果纺锤状椭圆形,种子腹面具1纵槽。以上主要特征显著区别于近缘种湿生车前和疏花车前。
研究材料一部分来自野生种群,该种群位于重庆市江津支坪镇涂家村附近长江与綦江交汇处的江心小岛——大中坝(29°16′56″ N,106°23′12″ E,平均海拔202 m);另一部分为2016年5月、2017年2、3月依次从上述野生种群移栽于西南大学生态园内的移栽种群。
1.2 研究方法
1.2.1花部形态特征与开花动态观测
在丰都车前进入生殖生长时,对其野生种群进行开花动态观察。种群中第1朵花开放起,每天9:00点观察1次,直至种群花朵全部凋谢为止,时间间隔即为种群花期;挂牌标记人为干扰较小的花序20个,记录花序花期,即每个花序的第1朵花开放至最后1朵花凋谢;进行花部形态和单花开花进程观测时,每天7:00—18:00每隔2 h定点观测1次并拍照,测量并记录花部构成、雌雄蕊位置形态变化、花瓣颜色、开花直径、色味有无、花药开裂等直至花朵枯萎。以花瓣开口处花丝可见的雄蕊初露期为开花标志,花瓣、花药变黄或枯萎为开花结束标志[4]。同时,记录每天的天气变化情况。
1.2.2花粉活力与柱头可受性
对野生种群不同发育阶段的花,采用MTT法[5-6]检测花粉活力与柱头可受性。具有活力的花粉或具有可受性的柱头被染成蓝色,反之则不被染色。
花粉活力=被染色花粉数/测量花粉总数×100%
1.2.3花粉胚珠数与杂交指数
(1)单花花粉量、胚珠数和P/O比值
随机选取野生种群中即将开放的花20朵,在解剖镜下轻轻剥去花冠和花瓣,用镊子取下所有花药于1.5 mL离心管中,滴加5 mol/L的氢氧化钠溶液1—2滴,10 min后用解剖针将花药碾碎,滴加蒸馏水至1.5 mL,置离心管于超声波清洗仪中处理30 min[4],使花粉从花药中分离并均匀分散于液体中。处理完成后,取10 μL悬浮液于载玻片上,在显微镜10倍放大视野中统计花粉数量,每个重复3次,计算出单花花粉量。同时将对应花的子房置于解剖镜下进行解剖,统计每朵花对应的胚珠数。计算出花粉胚珠比(P/O),即平均单花花粉量除以平均单花胚珠数,再依照Cruden[7]的评判标准得出丰都车前的繁育系统类型。P/O值降低,则近交程度升高;P/O值升高,则远交程度上升[7]。
(2)杂交指数OCI的估算
对野生种群中丰都车前开花过程和开花直径进行观察测量,根据Dafni[8]的标准判断其杂交指数。具体方法是:(1)花朵直径<1 mm记为0,1—2 mm记为1,2—6 mm记为2,>6 mm记为3。(2)花药开裂时间与柱头可受期同时或雌蕊先熟记为0,雄蕊先熟记为1。(3)柱头与花药的空间位置同一高度记为0,空间分离记为1。三者之和为杂交指数OCI值。
1.2.4单花花粉散出量变化
在丰都车前开花前随机标记野生种群中同一发育阶段的花30朵,测量丰都车前开花后单花花粉散出量随时间的变化。具体方法是分别采集开花后4 h、1 d、2 d等不同时间的花朵,参照单花花粉数的测量方法统计开花后不同时段单花花粉散出后的剩余量,前后相邻时间段剩余量的差值即为对应时间段的花粉散出量。
1.2.5人工授粉实验
选择丰都车前移栽种群中长势良好的植株花序,进行以下6组授粉处理研究:(1)不做任何处理,检测自然条件下的结实率,3个重复,共80朵花;(2)去雄套袋,在花药露出并还未开裂时去雄、套袋,验证是否存在无融合生殖,3个重复,共70朵花;(3)套袋自交,直接在花蕾期套袋,检测是否存在主动自花自交,3个重复,共59朵花;(4)同株异花授粉,在花蕾期套袋,待柱头具可受性时,取同株异花花粉授粉、套袋,在花药露出并还未开裂时去雄、再套袋,3个重复,共40朵花;(5)异株异花授粉,在花蕾期套袋,待柱头具可受性时,取异株异花花粉授粉、套袋,在花药露出并还未开裂时去雄、再套袋,3个重复,共48朵花;(6)自然异花授粉,去雄、不套袋,检测在室内通风处是否存在风媒传粉,2个重复,共48朵花。
结实率=结实数/处理花数×100%
1.2.6访花昆虫观察
2016、2017年的3—5月份于野生种群盛花期连续数日观察,并捕捉访花昆虫带回实验室制成标本,用作鉴定。选择天气晴朗的时候对访花昆虫定点拍照、摄像,记录访花行为、访花时间、访花频率等。
1.2.7分布地水位变化
分布地水文变化规律依据沈倩[9]、袁慎鸿[10]等对三峡库区相关研究,再结合实地考察得出的数据。
2 结果与分析
2.1 丰都车前的开花习性及花部性状
丰都车前的穗状花序具9—32朵单花,自下而上次第开放。单花花期3—4 d,花序花期5—8 d,种群花期从每年2月下旬持续至5月中旬,长达3个月,种群开花过程处于三峡库区现行水位调节变化状态的消落期,此阶段水位低于分布地海拔,开花授粉等不受水淹影响。单花开花动态见(图1A—M)。参考刘方炎[11]、徐凯[12]分别对滇榄仁和长萼兰花蕉单花开放进程的划分标准,以及丰都车前开花过程雌雄蕊变化情况,其单花开花过程可分为6个时期:①花蕾期,随着花序不断发育,花梗延长,花蕾增大(图1A、B);②雌蕊伸长期,柱头先露出花冠外部,不断伸长生长,红色花冠也渐渐露出,被青绿色的萼片包围(图1C);③雄蕊初露期,花瓣包着花药渐渐露出超过最外层的萼片,随着雄蕊不断生长,4个花药基部连着弯曲花丝相互向内靠拢 (图1D、E),此时萼片边缘的颜色由青白色变为淡红紫色,雌蕊从顶端开始干枯,由乳白色变为黄棕色;④花瓣展开期,花瓣展开,弯曲花丝慢慢伸展,带着花药陆续弹出花冠口,花药逐个纵向开裂(图1F—I),0.5 h内几乎全部开裂,同时花丝伸展,花朵完全开放,此时雄蕊高于柱头伸展在花柱四周,四个花瓣成十字型展开;⑤花瓣反折期,花药由鲜亮的红紫色逐渐变为干枯的浅棕褐色,花瓣逐渐向后反卷,有的甚至反向包住花冠口,且变得干枯 (图1J、K);⑥凋谢枯萎期,花丝逐渐枯萎,花药下垂(图1L),花药低于柱头,直至花朵凋谢(图1M)。

图1 丰都车前开花过程中花部形态变化及花部特征Fig.1 Plantago fengdouensis flower features and changes of flower Shape in flowering processA、B花蕾期;C雌蕊伸长期;D、E、F雄蕊展露期;G、H、I花瓣展开期;J、K花瓣反折期;L、M枯萎凋谢期; O丰都车前生境照
丰都车前花部形态特征见表1和图1(N1、N2),开花直径约3.25 mm,每花具苞片1枚,三角状卵形,萼片2对;花冠联合、淡黄、红粉色,无毛,冠筒与萼片约等长,花冠裂片狭三角形,在开花后反折;4枚雄蕊着生于花冠筒内面近基部,花药狭卵形、红黄色,干后变为淡棕色,花药基部与花丝相连,为基着花药(图1:N1、N2)。大部分花朵在开花时,雌蕊已干燥枯萎;少部分则在花朵开放后才逐渐枯萎。

表1 丰都车前花形态特征
2.2 丰都车前花粉活力与柱头可受性

图2 丰都车前花粉活力与柱头可受性变化Fig.2 The variation of pollen viability and stigma receptivity of Plantago fengdouensisA:花蕾期;B:雌蕊伸长期;C:雄蕊初露期;0—6 h:花瓣展开期:6—36 h:花瓣反折期;36 h之后:枯萎凋谢期
采用MTT法[5-6]检测丰都车前不同阶段花粉活力和柱头可受性,结果如图2、图3所示:丰都车前在花药未开裂前花粉已有较高活力(约80%),花药开裂后的0—3 h内花粉活力增至最强,有的达到90%以上,开花后24 h内保持在30%以上。第2 d开始花粉活力急剧下降,1.5 d后不到20%,至第3 d几乎没有活力。

图3 丰都车前不同阶段柱头可受性变化情况Fig.3 Stigma receptivity of Plantago fengdouensis in different floral stages左上角标识为柱头可受性,+示柱头具可受性,++示柱头具较强可受性,+++示柱头具最强可受性,+/-示部分柱头具可受性,-示不具可受性
丰都车前在花瓣展开前柱头可受性已达最高,之后逐渐减弱。其中,大部分柱头在花瓣展开时已枯萎失活丧失可受性,少部分在花药开裂后仍具较弱可受性。花粉活力最高期滞后于柱头最佳可受期,一定程度上避免了因自花传粉而导致种群衰退。但有少部分重叠,也为自花授粉提供可能,在外界环境突变时保障了种群的生殖成功。
2.3 丰都车前花粉散出变化
丰都车前花粉散布变化检测结果如图4所示:丰都车前花朵开放后花药立刻相继开裂,花粉不断散出。4 h内花粉散出量为总量的1/6, 1 d后单花花粉剩余量约为开花前单花花粉量的1/2,说明丰都车前在开花后第1 d内花粉散出量大,这与丰都车前开花后的花粉活力变化保持一致,保障了丰都车前传粉过程的及时性和有效性。
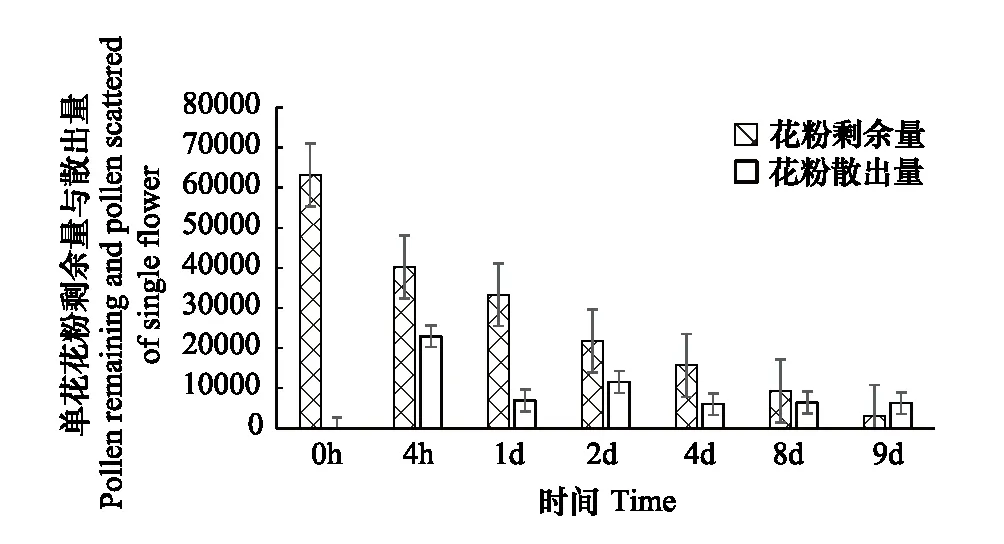
图4 丰都车前花粉散布变化Fig.4 Pollen dispersal of Plantago fengdouensis
2.4 繁育系统
2.4.1花粉/胚珠比(P/O)
花粉胚珠比是单花花粉量与胚珠数的比值。统计计算得出结果见表2:丰都车前的P/O约为15373.29,介于2108.0—195525.0,依照Cruden[7]的评判标准,属于专性异交类型。
2.4.2杂交指数(OCI)
杂交指数(OCI)统计结果见表2:丰都车前的花冠直径为3.25 mm,介于2—6 mm之间,记为2。花药4个,在花药未开裂前,柱头已成熟具有可受性,记为0。柱头与花药的空间位置在不断变化,在4个花药分离前,弯曲花丝未伸展时,柱头高于花药;完全盛开时,则花药高于柱头;随着花继续开放,花药开裂,花丝慢慢枯萎弯曲,花药向四周下垂,低于柱头,空间分离记为1。所以丰都车前的杂交指数OCI为3,根据Dafni[8]的标准,其繁育系统为部分自交亲和,有时需要传粉者。
花粉胚珠比及杂交指数结果表明:丰都车前的繁育系统类型是以异交为主,自交亲和,有时需要传粉者。
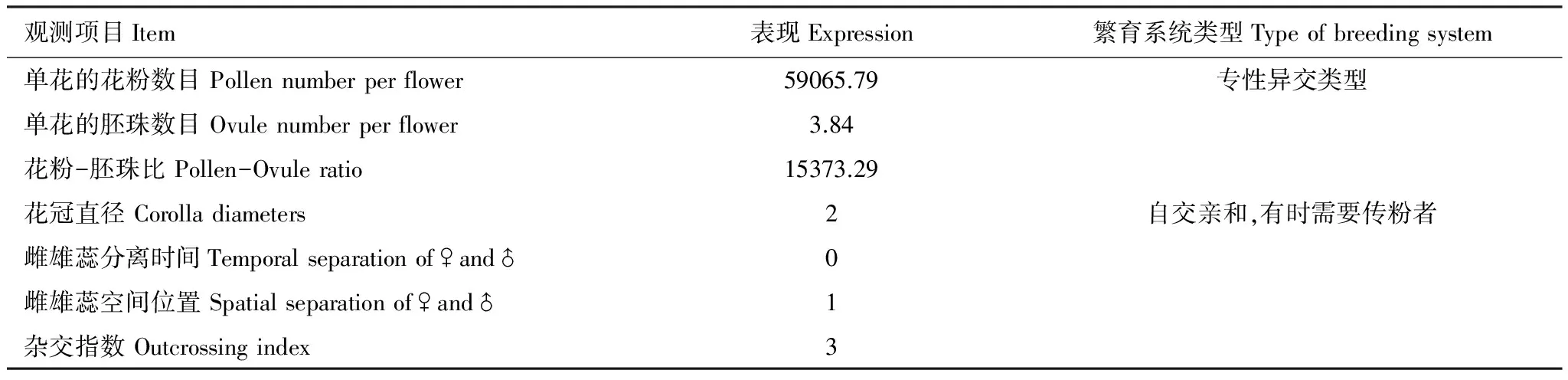
表2 丰都车前的花粉胚珠比和杂交指数
2.5 套袋实验
套袋实验结果(表3),丰都车前在自然条件下能够结实,结实率为100%;去雄套袋结实率为0,即不存在无融合生殖;未去雄、直接套袋处理条件下结实率为31.54%,存在自主自花授粉现象;异株异花、同株异花、去雄不套袋自然异花授粉处理下,结实率为依次为91.85%、 57.78%、70.84%,结实受传粉者限制。
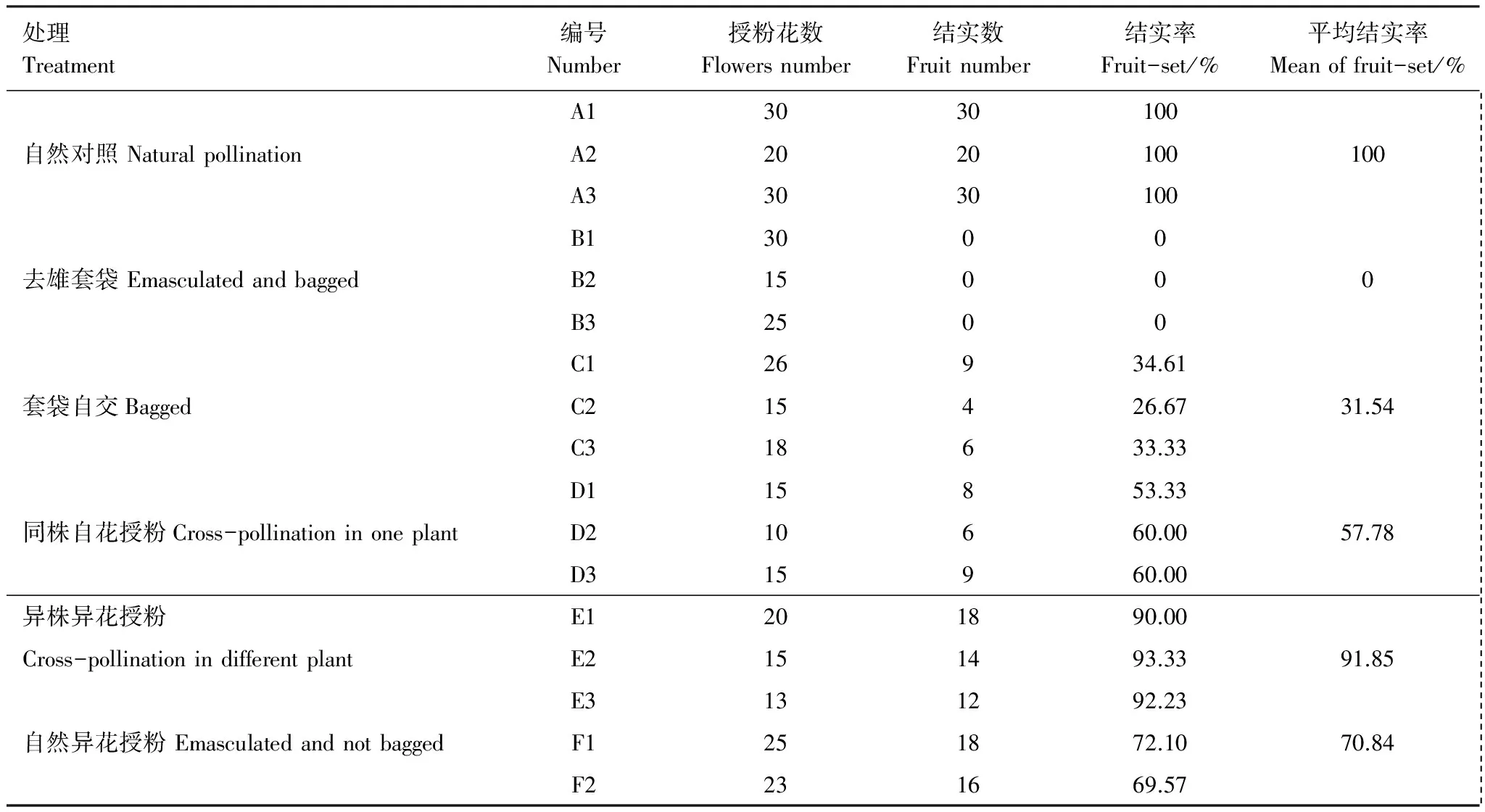
表3 不同处理条件下丰都车前结实率的差异
2.6 访花昆虫及传粉行为
丰都车前花朵艳丽,能有效吸引昆虫访花(图5)。当日开花时间多集中在每日8:00—12:00、 9:00—11:00呈现开花高峰。花开放0.5 h内花药全部开裂,大部分访花昆虫的访花高峰时间段为晴朗天气的9:30—12:30。对访花昆虫鉴定,包括中华蜜蜂(图5)和2种食蚜蝇,其一为斜斑鼓额食蚜蝇(图5);从访花昆虫的访花行为、访花频率等综合考察,中华蜜蜂为主要的访花昆虫,其次是斜斑鼓额食蚜蝇,少见另一种食蚜蝇。中华蜜蜂偏好访问刚开放的花朵,单花停留时间(5.0±1.8)s,平均访花频率为11.5朵/min,通过停落在花瓣和柱头上采集花粉带于后足花粉筐,访花后,在头部、胸部腹面、足等身体多处布满花粉,随后以连续访问的方式访问附近花朵,是丰都车前最有效的传粉者。2种食蚜蝇则更偏向访问开花时间较久的花朵,访花后虽然身体同样沾有花粉,但此时柱头几乎不具可受性,花粉活力较弱,不能起到有效传粉。综合比较,2种食蚜蝇的传粉贡献不及中华蜜蜂。

图5 丰都车前的访花昆虫Fig.5 Insect visitors of Plantago fengdouensis从左到右为斜斑鼓额食蚜蝇Scaeva pyrastri (Linnaeus),斜斑鼓额食蚜蝇Scaeva pyrastri (Linnaeus),中华蜜蜂Apis cerana
2.7 水位变化
丰都车前江津支坪镇大中坝分布地长江水位变化如图6所示,结合其分布地平均海拔约202 m可以得出,每年的6月至10月为分布地受夏季洪讯干扰的淹没期;10月末至次年的5月都是处于低水位,即落水期,此阶段丰都车前不仅完成了植株叶的生长,在每年3—5月份还顺利开花座果。因此,分布地的夏季洪讯并没有影响丰都车前开花传粉;相反,其花期与分布地水位落水期相协调进一步保障了丰都车前的续存。
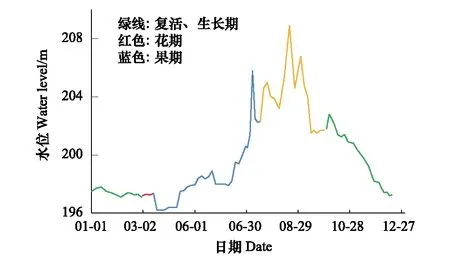
图6 丰都车前江津分布地水位变化Fig.6 Change of water level in Jiangjin population of Plantago fengdouensis落水期:10月—次年5月;洪汛、涨水期:6月—10月
3 结论与讨论
3.1 丰都车前花部特征与繁育系统关系
植物花部特征分为花部构成和开花式样两个层次,二者综合作用影响对传粉者吸引和花粉的散布,进而主导开花植物的交配机遇[13]。从丰都车前的花部特征来看,其花瓣、花药颜色艳丽,雌雄蕊露出花冠外,花冠、萼片较长时间保持新鲜状态且宿存,从而增加整个花序的花展示面积等均能有效吸引昆虫前来传粉;雌雄蕊露出花冠外的花部特征也增大传粉者与柱头的接触机率,增大了授粉机遇,表现出对虫媒传粉的适应。另一方面,丰都车前花小、花多密集成穗状;花粉粒小、花粉量极大,花粉干燥且易于被风吹走;雄蕊花丝细长、花药外展于空气中,与花丝连接不紧密,雌蕊柱头较长且成羽毛状等均适合风媒传粉的特点,也表现出对风媒传粉的适应性。这在套袋实验得到进一步证实。套袋实验表明,丰都车前不存在无融合生殖,自花可育;在异株异花授粉、同株异花授粉、去雄不套袋处理下,结实率依次降低,表明丰都车前以异花授粉为主,自花授粉为辅,传粉者在有性生殖过程中具有一定贡献。统计位于室内通风处的自然异花授粉组结实情况,得到有一定结实率,表明了风力也是丰都车前的传粉媒介。
在传粉过程中,较强活力的花粉能否及时遇到较高可受性的柱头是传粉成功的关键,该过程受环境因素的影响[14],雌雄异熟、异位被认为是植物避免花内自交、提高异交率的机制[15-16]。丰都车前不同发育阶段花粉活力和柱头可受性检测结果可以看出,丰都车前具有明显的雌雄异熟特征,表现为雌蕊先熟。柱头在花瓣展开前先伸出花冠口、达到最高可受性;雄蕊在花瓣展开后才露出花冠外、之后花药开裂,而此时大部分柱头已枯萎失活丧失可受性,只有少部分具可受性,一定程度避免了自花授粉的发生,从而提高了异交机会,但少部分具较弱可受性,也为自花授粉提供可能。同株异花授粉的交配方式,可能使自交亲和的物种出现自交衰退[17-18],丰都车前无性繁殖方式——根颈基部萌生新芽、“集中开花模式”为同株异花授粉提供了机会。从测定的P/O比值看,花粉/胚珠比约为15373.29,依照Cruden[7]的评判标准,丰都车前应属于专性异交类型。但杂交指数OCI值为3,根据Dafni[8]的标准,其繁育系统为自交亲和,有时需要传粉者。综合得出:丰都车前的繁育系统属于异交为主,自交亲和,有时需要传粉者,为风媒和虫媒混合传粉类型,这与长叶车前[19-20]、北美车前[21-22]等车前属植物结论一致,此种混合传粉类型在流苏树中也有类似的报道[23]。
3.2 丰都车前开花特性与传粉
有花植物传粉过程中,传粉者起着关键作用。不同访花昆虫有着不同的访花行为,不同的访花行为产生不同的访花效率,进而对开花植物产生不同的传粉效应[24]。丰都车前的主要访花者中华蜜蜂喜好访问刚开放的花朵,同时在开花第1 d内丰都车前花粉散出量最大、花粉活力也最强,两者协调一致,保障了传粉的及时性和有效性;腹触式传粉是中华蜜蜂对木荷最有效的传粉的方式[25],中华蜜蜂也是通过飞行采集丰都车前的花粉,过程中其胸腹部能充分接触花药和柱头,能够携带大量花粉,并将花粉传递给接下来访问的花朵,实现了植株的异花授粉。两种食蚜蝇则偏向访问开花时间较久的花,此时柱头大部分已不具可受性,花粉活力较低;推测它们不是主要的传粉者。但以上两种传粉者不同时期访花,避免对花粉的竞争,使有效传粉者能充分发挥作用。
丰都车前单花序开花同步性高、数量多,表现出“集中开花模式”,多花的开花式样能吸引较多的访花昆虫和访花者连续访花[26-28]。延长花期是植物应对生长环境易变或传粉昆虫稀少所形成的适应机制[29-30]。丰都车前从2月下旬至5月中旬,整个期间都有花序不断发育开放,花序多且穗状花序单花量大,种群花期表现出“持续开花模式”,能够持续吸引昆虫访花,这种开花模式的意义在新疆两种柽柳[31]的研究中也有报道。
3.3 丰都车前的开花物候与分布地枯水期的预适应
水分胁迫是三峡库区消落带内植被生存最主要的限制因素之一。丰都车前原有的分布地——重庆市忠县、丰都和巴南区3个长江江心岛,因所在海拔低于三峡水库最低蓄水位而被完全淹没,三峡库区江津段属于受冬季蓄水和夏季洪汛双重影响消落区[32]。本次研究的丰都车前大中坝种群位于库区尾端,水位波动不大,丰都车前大部分植株不被水淹,因而迄今仍残存。其开花时期正处于三峡库区现行水位调节变化状态的消落期——水位较低,洪汛到来前大部分植株便完成了开花过程及座果,从而避开了夏季汛期水淹对开花过程的迫害,这与袁慎鸿[10]的研究结论相符;同时,该野生种群分布地平均海拔高于三峡库区最高蓄水位,水淹浅,出露早,为丰都车前完成生活史提供可能,进一步保障了该种群的续存。与黄成名等[33]在三峡植物园管理处观察结果相比,各生殖期有所提前,这可能是因为江津野生种群与三峡植物园管理处丰都车前的生境不同,或者是应对库区新的水位节律变化做出的一种预适应。对于其种子能否以及如何度过淹没期,种子的活力、萌发情况如何等都还有待进一步探讨。
致谢:中国科学院动物研究所袁峰教授鉴定传粉昆虫,西南大学生命科学学院陶建平教授鉴定丰都车前,西南大学生命科学学院李文平、倪东萍、李丘霖老师对野外观察记录和实验给予帮助,特此致谢。
