川西林线交错带岷江冷杉幼苗异龄叶形态对长期模拟增温的响应
2018-07-18曾瑞琪赵家国刘银占陈国鹏鲜骏仁
曾瑞琪,赵家国,刘银占,陈国鹏,鲜骏仁,*
1 四川农业大学环境学院, 成都 611130 2 旺苍县林业和园林局, 广元 628200 3 河南大学生命科学学院,全球变化生态学国际联合实验室, 开封 475004 4 甘肃农业大学林学院, 兰州 730070
全球气候变暖已是不争的事实。IPCC在2013的评估表明:1880—2012年间全球平均气温增加了0.85℃,预计到21世纪末还将增加0.3—4.8℃,全球温室效应仍在继续和扩大,对高寒地段的影响将更大[1]。温度控制着生态系统中几乎所有生化反应速率,直接或间接作用于植物生理过程,进而调控植物功能性状的变化[2- 5],是生态系统过程中重要的驱动力之一。叶片是植物进行光合作用的主要器官,其生长发育和形态特征直接影响植物的行为和功能[6]。叶形态是反映植物适应环境变化所形成生存对策的重要指标之一,如叶面积与植物光合作用、呼吸作用、蒸腾作用等均有直接或间接地联系,是评价植物群落净初级生产力、衡量群落结构是否合理的重要指标[7- 8];叶片厚度则会影响植物对资源的获取和利用[9- 10]。植物生长过程中,各叶形态之间有很强的关联性[11]。不同年龄叶片(下文简称“异龄叶”)由于其形成年份、发育阶段等的不同,其叶片物质分配、环境敏感性均有差异[12- 14]。持续上百年的全球变暖已显著影响到植物的物候和生理过程[15- 16],在变化环境条件下,由于异龄叶叶形态响应增温的敏感度不同,其可塑性也势必存在差异[17- 19]。因此,开展长期模拟增温对异龄叶片叶形态的影响研究对准确预测植物适应全球变暖的模式是十分重要的。
川西亚高山林线交错带是长江上游典型的生态敏感和脆弱区,是针叶林向高山草甸过渡的关键区域,相对简单的植物群落结构、较矮的植物高度及林线普遍存在的边缘效应等,为了解高寒生态系统对气候变化的响应提供了理想的天然研究平台[20]。在全球变暖的背景下,亚高山林线交错带极有可能被其下缘针叶林建群种入侵。岷江冷杉(Abiesfaxoniana)是川西北亚高山针叶林的建群种之一,位于林线交错带之下。在全球变暖的背景下,岷江冷杉的叶片是否会在形态上表现出对长期增温的适应,这种适应是否存在年际差异?以往研究显示,增温条件下,岷江冷杉幼苗芽开放提前,叶片呈现变长变薄的趋势,且叶面积、比叶面积显著增加,比叶重显著下降,物候和侧枝生长也对增温表现为正响应[20- 21],但都并未系统涉及岷江冷杉幼苗叶片形态。因此,本研究假设岷江冷杉幼苗异龄叶会对长期模拟增温产生差异性响应。利用前期设置于此的开顶式生长室(Open-top chamber,OTC)模拟全球变暖,研究长期模拟增温对岷江冷杉幼苗异龄叶片叶长、叶宽、叶面积和叶厚等形态的影响,并分析其可塑性变化,探讨岷江冷杉叶功能性状对长期模拟增温的响应,旨在为全球变暖背景下的亚高山森林群落功能变化研究提供基础数据。
1 材料和方法
1.1 研究区概况
本研究位于四川省平武县王朗国家级自然保护区(32°59′N, 104°01′E,海拔2300—4980 m)。地处青藏高原东缘,属山地季风性气候[22]。年平均气温1.5—2.9℃,7月平均气温12.7℃,1月平均气温-6.1℃。年降水量在801—825 mm之间,雨季多集中在5月到8月[16]。岷江冷杉林(2600—3500 m)是保护区主要森林群系,土壤为暗棕壤[23],0—15 cm和15—40 cm深度,土壤有机碳分别为(180.2±5.5)、(180.2±3.7) g/kg,土壤总氮为(4.5±0.8)、(0.8±4.0) g/kg,土壤总磷为(5.5±0.7)、(4.1±0.5) g/kg[24]。
1.2 样地设置
2005年9月,在大窝凼外侧坡树线交错带的平缓地段设置 3个1.5 m×1.5 m×2.6 m、顶端开口面积1.2 m×1.2 m的OTC,OTC由聚碳酸酯制作,每个OTC间距在5 m以上[20]。同时在OTC附近设置3个对照样方(CK),其底面积同样为1.5 m×1.5 m。10月,从样地附近移植长势良好且相当的目标物种到OTC及CK内,岷江冷杉年龄大约12—15 a,株高约50—60 cm。在OTC及CK内均种植5株,即中心1株,4个拐角各1株,拐角幼苗据OTC边缘均30 cm。2006年,定期对OTC及CK内物种浇水,大约间隔15 d一次,以确保物种存活。每个OTC及CK内均放置温度传感器(6507A,±0.1℃,Finland)。监测表明:在生长季,OTC中空气温度(1.2 m)平均较CK高2.9℃,土壤温度(5 cm)增加0.4℃[20]。增温全年进行,试验地周围用网围栏封育,避免动物干扰,OTC和CK均不再人工施肥和浇水,OTC外膜保持洁净,期间不定期对OTC装置进行维护。
1.3 研究方法
1.3.1植物取样
本实验以岷江冷杉幼苗的1 a叶、2 a叶、3 a叶为研究对象。2016年8月下旬,用枝剪从OTC及对照样地内逐株个体的不同方向采集生长良好的枝条,根据枝条上芽鳞痕形成的环节判断枝段年龄,并在其上采集叶片[25],被采集叶片均保证其完整性且没有发黄衰老症状。叶片随机采集,每个OTC、CK内各龄级保证10个有效样本,则不同处理下各龄级样本分别为30个,分别装入自封袋,记录编号带回实验室。用数显卡尺(±0.01 mm)测量叶长、叶宽、叶厚,使用叶面积仪(CI- 203,CID,USA)测定单叶面积。
1.3.2统计分析
数据整理和统计分析在Microsoft Excel 2010(Microsoft Corporation, Redmond, WA)和SPSS 20.0(SPSS Inc., Chicago, IL, USA)上进行,作图在Origin 8.0(Origin Lab, MA)上进行。有温度和叶龄二因素,温度类型为主区处理因素,叶龄为副区处理因素,对两种条件下叶片各形态用裂区试验方差分析方法进行显著性分析,用邓肯多重比较(Duncan′s multiple range test)检验在方差分析中有差异的变量间的差异显著性。利用箱图来展现增温实验下异龄叶叶形态其数据整体位置和分散情况,箱图中5个线段由上到下分别代表该形态最大值、上四分位数、中位数、下四分位数、最小值。采用表型可塑性指数(phenotypic plasticity index,PPI)解释表型可塑性的大小:
PPI = (MAX-MIN)/MAX
式中,MAX和MIN分别表示该环境下的最大和最小形态值,PPI取值为0—1[11,26],其值的大小与表型可塑性的大小呈正相关。通过PPI计算叶表型可塑性大小,并与常用的表型可塑性指标变异系数(coefficient of variation,CV;CV=标准差/平均值)比较。
2 结果分析
2.1 增温和叶龄对叶形态的影响
2.1.1叶长、叶宽及长宽比的响应
OTC增温极显著降低岷江冷杉幼苗叶片的长度和宽度(P<0.01),对长宽比的影响很小(表1);区组间对叶长的影响不显著,温度、叶龄及交互作用均对叶长有显著影响(表2);仅叶龄对叶宽产生显著性作用(P<0.05),其余因素影响均不显著(表2)。
岷江冷杉幼苗叶片各叶形态值的范围和离散程度对增温、叶龄的响应有差异。OTC中1 a叶的长度较对照增加2.01%(表1),叶长值变化范围(12.24—25.98 mm)较对照(10.63—28.31 mm)减小、分布更汇聚(图1A);OTC中2 a、3 a叶的叶长比对照极显著降低21.87%(P<0.01)、15.11%(P<0.01)(表1);增温条件下2 a、3 a叶长值分布更汇聚,特别是2 a叶(OTC:1.331—2.527 mm;CK:1.801—3.112 mm)(图1B、C);增温处理下异龄叶间的叶长差异不显著,而对照中差异显著(P<0.05)(表1)。增温使1 a、2 a叶叶宽比对照极显著减小13.48%、13.99%,3 a叶叶宽较对照减小8.43%(表1),叶宽分布更分散(图1D、E、F);增温与对照条件下,异龄叶间叶宽值变化均不显著(表1)。增温未对叶长宽比产生显著影响(表1),但叶龄间响应并不一致:增温使1 a叶整体长宽比值比对照显著增大18.82%,值分布较对照小,表现出明显的汇聚(图1G,表1);增温使2 a叶、3 a叶整体长宽比值分别比对照减小7.93%、5.70%,值分布范围更分散(2 a:6.02—20;3 a:5.35—12.44)(图1H、I,表1);增温处理下的2 a叶长宽比分别与1 a、3 a叶有显著差异(P<0.05),而对照处理下的异龄叶间叶长宽比变化均不显著(表1)。

表1 模拟增温对岷江冷杉幼苗叶形态的影响
同排不同小写字母表示增温处理下不同年份间存在显著差异 (P<0.05);同排不同大写字母表示对照处理下不同年份间存在显著差异 (P<0.05); *表示在0.05水平上有显著差异;**表示在0.01水平上有显著差异;OTC:开顶式增温室 open-top chamber;CK:控制对照 control check
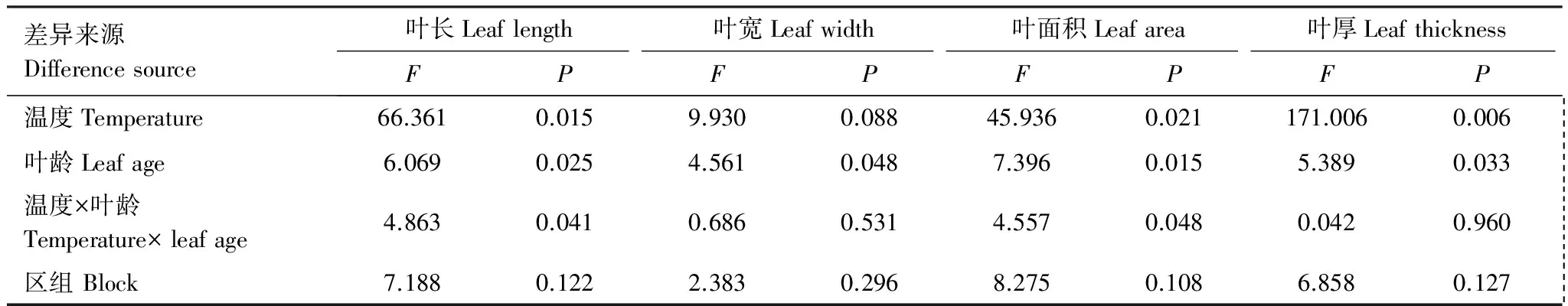
表2 裂区分析的方差分析结果

图1 增温对岷江冷杉幼苗叶长、叶宽以及叶长宽比的影响Fig.1 Effects of simulated warming on the leaf length, leaf width, and leaf aspect ratio of A. faxoniana seedlings图中黑点表示各形态值的大小分布情况
2.1.2叶面积、叶厚及叶体积的响应
OTC极显著降低岷江冷杉幼苗的叶面积(比对照减小17.76%,P<0.01)、极显著增加叶厚(比对照增加7.27%,P<0.01)、显著降低叶体积(比对照减小11.49%,P<0.05)(表1);区组间对叶面积的影响不显著,温度、叶龄及交互作用均对叶面积有显著影响(表2);温度对叶厚有极显著影响(P<0.01),叶龄对叶厚有显著影响(P<0.05),温度和叶龄的交互作用、区组间对叶厚的影响均不显著(表2)。
岷江冷杉幼苗叶面积、叶厚、叶体积对增温、叶龄的响应有差异。增温抑制异龄叶片叶面积扩展,特别是2 a叶,减少达28.41%(表1),OTC内叶面积介于14—64 mm2,较CK(28—64 mm2)分布范围增大(图2B);1 a、3 a叶叶面积较对照分别减少12.97%、8.81%,离散程度较对照增大(图2A、C,表1);增温处理的异龄叶间叶面积变化均不显著,对照处理的2 a叶叶面积分别与1 a、3 a叶有显著差异(P<0.05)(表1)。增温促进异龄叶片变厚,1 a、2 a、3 a叶厚分别比对照增大7.84%、3.51%、6.78%,分布范围也被扩大,值更发散(对照处理中叶厚值多集中在中位数附近),其中1 a、3 a叶增温后值分布范围为0.31—0.74 mm、0.43—0.86 mm,而对照内为0.41—0.63 mm、0.45—0.69 mm(图2D、E、F,表1);增温处理下3 a较1 a叶叶厚变化显著,对照处理的1 a叶叶厚分别与2 a、3 a叶差异显著(P<0.05)(表1)。增温抑制异龄叶叶体积膨大,使其较对照显著减小11.49%,值更发散:OTC内1—3 a叶叶体积范围为4.4—32.4、 7.92—37.76、4.3—35.2 mm3,而CK内分别为7.74—28.42、11.76—40.6、9.52—29.48 mm3(表2,图2G、H、I)。增温处理下异龄叶间体积变化均不显著,对照处理的2 a叶分别与1 a、3 a叶叶体积差异显著(P<0.05)(表1)。

图2 增温对异龄岷江冷杉幼苗叶面积、厚度以及体积的影响Fig.2 Effects of simulated warming on the leaf area, thickness and volume of A. faxoniana seedlings
2.2 增温对异龄叶叶形态可塑性的影响
2.2.1叶长、叶宽可塑性的响应
表型可塑性指数分析结果表明(图3):增温提高了岷江冷杉幼苗异龄叶叶宽的可塑性和2 a、3 a叶叶长的可塑性,增幅最大达68.22%(3 a叶叶宽)(图3);降低当年生叶叶长的可塑性、降幅为15.32%(图3)。除2 a叶叶宽的CV对照大于增温外(图3),其余各龄级指标CV的比较结果与PPI一致。说明2种表型可塑性分析方法的结果相似,均可用于分析长期增温对岷江冷杉异龄叶叶片长、宽影响的研究。
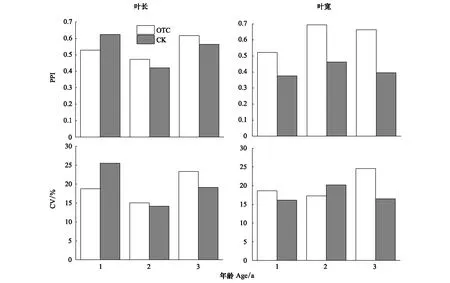
图3 岷江冷杉幼苗叶长、叶宽表型可塑性指数(PPI)和变异系数(CV)Fig.3 The phenotypic plasticity index (PPI) and coefficient of variation (CV) of leaf length, leaf width of A. faxoniana seedlings
2.2.2叶面积、叶厚可塑性的响应
两种可塑性结果分析均表明(图4):增温条件下叶面积和叶厚度较对照均有较大的可塑性,但不同方法中,可塑性增长比例不同,如增温条件下2 a叶叶厚的CV较对照增大69.02%,而PPI仅增大12.78%(图4);3 a叶叶面积PPI增大16.66%,而CV仅增大8.03%(图4)。增温处理下3 a叶叶厚的CV值是各龄级较对照增幅最大的叶厚可塑性,即比对照增大143.57%(图4)。

图4 岷江冷杉幼苗叶面积、叶厚表型可塑性指数(PPI)和变异系数(CV)Fig.4 The phenotypic plasticity index (PPI) and coefficient of variation (CV) of leaf area, leaf thickness of A. faxoniana seedlings
3 讨论
3.1 叶形态对增温的响应
叶片的扩展性生长(如叶片长、宽、面积、厚度)极易受到周围环境因子,特别是温度的影响。高寒地区的环境温度普遍低于该区域植物生长所需的最适温度,故增温通常能促进极地和高山地区植物的营养生长[27- 29]。本研究发现,增温显著降低异龄岷江冷杉幼苗叶长、叶宽、叶面积形态值,这与前人研究结果不一致:如徐振峰等的研究表明,短期(2 a)模拟增温促进岷江冷杉幼苗叶片增长变薄[20];杨兵等的研究也表明,长期(65个月)模拟增温亦促进岷江冷杉幼苗叶面积的增大[21]。导致这种差异的原因可能有二:一是模拟增温的时间长短不同;二是研究期间土壤的水分条件不一样:徐振峰等研究的2007年降雨较当地多年平均降雨量大[20],杨兵等其研究过程中则不定期浇水以保证土壤水分适于植物生长[21]。在利用OTC自然增温过程中,个别年份会因降雨充足致使土壤水分含量下降不明显,但长期增温过程中土壤水分降低的趋势不会改变(一是因为OTC开口较小影响雨水进入,二是增温加剧土壤水分丢失)[30- 31],这一负面效应使叶片通过减小叶片平均叶长、叶面积等,使叶形态倾向旱生,来达到防止过多水分散失、保持最适叶表温度的目的[32]。同时,减小叶面积也起到降低高蒸腾伤害的作用,是叶片对温度更高、更干旱环境的适应方式[16,33],这与Yates等提出的植物因有较小叶面积来自我保护的结论一致[34]。结果显示,OTC长期增温效应使冷杉叶片叶厚显著大于对照(P< 0.05)。叶片增厚,保水能力增强[35],也进一步验证了之前的分析,即该增温条件下岷江冷杉幼苗叶片形态向更适应相对干热生境的方向调整。同时,较小较厚的叶片也可减缓叶片在增温条件下的生长速度,利于储备更多的碳用于防御结构[36]。增温显著增大叶片体积,对叶片长宽比无显著影响,说明岷江冷杉幼苗叶片细胞分裂和体积扩展方向对增温的响应存在差异。
3.2 叶形态的年际差异
虽然增温在总体上改变岷江冷杉幼苗的叶片形态,但不同年龄之间叶形态亦有明显差异,表明年际间的温度和降水等环境因素的波动也会对植物的增温响应产生明显影响。这与Oguchi等对藜(Chenopodiumalbum)[19]和Frak等对杂交核桃(Juglansnigraxregia)[18]的研究结果一致。尽管本研究未提供长期的增温监测数据(2009年项目结束而未进行温度的连续监测),但从本实验的前期研究可推测:OTC增温的效应存在明显的年际差异,特别是降雨量差异较大的年份,这必然将影响植物的物候和叶片的生长进程,使叶片的形态特征表现出明显的年际差异。前期利用OTC短期增温的研究也证明了这一推测:2007年较多的阴雨天气,使王朗国家级自然保护区大窝凼外侧坡树线OTC在生长季节的温度增加2.2℃,由于大量降雨,增温并未导致土壤水分减少过多,因而增温促进了绵穗柳(Salixeriostachya)叶片长、宽的生长,进而增加单叶面积[37- 38];而2009年降雨不如2007年,生长季节OTC内的温度平均升高2.9℃、土壤水分平均减少2.4%,由此导致绵穗柳长度减小、宽度显著减小、叶面积显著降低,表现出明显的旱生趋向,尽管增温导致绵穗柳单个叶片的面积减少,但增温使单个植株上的叶片总数大量增加,由此弥补单叶面积的减小,因而总体仍表现为增温促进生长[16];OTC增温效应对绵穗柳的物候、叶寿命等也均存在明显年际差异现象[16,39]。持续OTC增温对棚内干旱情况的累积作用,也加剧了OTC内年度微气候的差异程度[31]。此外,较老叶片的次生代谢也可能导致叶片形态的改变,如Kayama等对3种云杉(Piceaglehnii,P.jezoensis和P.abies)的研究表明,钙离子和某些次级代谢产物在老叶中的积淀,导致叶片增厚[40]。因此,植物年际微生境中的适应性调整会导致叶形态表现出明显的年际差异。
3.3 叶表型可塑性及叶形特征参数对增温的响应
本研究表明,岷江冷杉幼苗叶形态(长、宽、面积和厚度等)适应模拟增温的方向总体均倾向于旱生,即减小水分散失、保持叶片最适温度,年度气候差异对叶形态的响应亦产生重要影响。但在增温和异龄两个条件交互影响下,对叶宽和叶厚两个形态没有显著性影响,可以理解为两个处理条件在叶宽、叶厚形态下的趋势抵消。表型可塑性存在于大多数植物中[41],其可通过使物种具有更宽的生态幅和更好的耐受性,来增强物种对变化环境的适应能力,使物种更好地占据地理和生境优势[14,42]。本研究结果显示增温总体上增大岷江冷杉幼苗叶形态的可塑性,说明在研究期内叶形态的调整是植物对异质增温环境的一个适应方式,但这种调整同样受到研究期间年际微气候差异的影响[43],即不同年龄的叶形态可塑性的变化幅度不同。其中1 a叶增温处理下的叶长表型可塑性较对照小,而2 a和3 a叶均是较对照大,进一步验证了这一观点,即“叶片适应存在明显年际差异”,同时增温条件下1 a叶长值比对照大但可塑性却比对照小这一结果,也说明在OTC增温条件下,小叶较小的蒸腾作用更能适应干热环境、保护自我[34]。
综上所述,OTC中的岷江冷杉幼苗叶片对长期增温作出相应的适应性调整;叶片形态特征表现出显著的旱生倾向,但因年际气候差异而存在明显的叶龄变化;完全支持实验初期提出的假说——“岷江冷杉幼苗异龄叶会对长期模拟增温产生差异性响应”。本实验仅从叶片形态的视角探讨岷江冷杉幼苗对长期增温的响应,将来的适应性研究还可尽量整合微生境气候等资料,辅以生理、树木构型、地下生态过程等方面,以全面揭示岷江冷杉对全球变暖的响应机制。
致谢:四川农业大学环境学院李泞钰、范潇月、陶媛和冯时等同学给予帮助,四川王朗国家级自然保护区提供采样便利条件,特此致谢。
