宜春富硒温泉中 产蛋白酶嗜热菌株的筛选与鉴定
2018-07-11吕美云
吕美云
(宜春学院化学与生物工程学院,江西宜春 336000)
目前,蛋白酶已广泛应用于食品、药品、洗涤剂及皮革等诸多工业领域[1]。而酶作为一类生物活性大分子具不稳定性,尤其在高温、强酸、强碱和高渗等极端条件下更容易失活,限制了酶在工业上的应用[2]。高温蛋白酶中的嗜热蛋白酶具有对高温、酸碱、有机溶剂、蛋白变性剂较稳定且有良好抗性,因此,高温蛋白酶的研究和应用备受关注[3]。一般认为蛋白酶的热稳定性与产酶菌株的生长温度相关,产蛋白酶嗜热菌株可从温泉、火山口、深海热泉等高温环境获得[4]。自1962 年对嗜热菌蛋白酶报道以来[5],相继从各地温泉环境中筛选产蛋白酶嗜热菌,以获得热稳定性较高的高温蛋白酶。祝伟等[6]从腾冲热海温泉样品中分离的1株高温产蛋白酶菌株YMTC 1049,其生长温度范围为50~80 ℃。舒丹等[7]从西藏的尼木卡如林温泉附近的土样中分离出一株嗜热碱性蛋白酶产生菌SD-142,其最适反应温度为 60 ℃,最适pH为9.5,具有良好的pH稳定性和热稳定性。张云峰等[8]从温泉中分离出一株产嗜热蛋白酶芽孢杆菌DPE7,当温度为65 ℃,pH为9.0是最适宜的产酶条件。李杨霞等[9]从我国西南地区分别采取热泉和地下热水水样,分离得到47株中度嗜热菌,这些菌株在67或70 ℃时,可分泌多种耐热的水解酶,其中 38%的菌株分泌蛋白酶。廉立慧[10]从徂徕山温泉附近土样中获得产高温蛋白酶菌株L7,其最适温度为 55 ℃,最适pH10,并且具有良好的热稳定性和pH稳定性。
宜春温泉是国内罕见的富硒少硫的温泉,位于江西省宜春市温汤镇,长年水温达68 ℃,目前对其蛋白酶生产菌的资源研究与应用鲜有报导。本文以宜春温泉温汤古井水样为样品,用脱脂牛奶蛋白平板,55 ℃高温培养的方法,筛选出产蛋白酶嗜热菌株,并对其形态进行观察、生理生化鉴定和生长特性的研究,以及菌株16S rRNA基因序列的分析,探究其种属和系统进化关系,以期丰富产蛋白酶嗜热菌资源,为产蛋白酶嗜热菌的后继研究和高温蛋白酶的开发利用提供理论依据。
1 材料和方法
1.1 材料与仪器
实验样品 采自江西宜春温汤古井水样(温度68 ℃,pH6.5~6.7),4 ℃保存;培养基 种子液培养基:牛肉膏0.3%、蛋白胨1%、NaCl 0.5%,pH7.2,121 ℃高温灭菌30 min,斜面培养基加2%琼脂;富集培养基:TYE 培养基[11];筛选培养基:TYE 培养基+底物(脱脂奶粉 黑龙江省完达山乳业股份有限公司);Ezup柱式细菌基因组提取试剂盒、即用PCR试剂盒 上海生工生物工程有限公司。
DYY-6C型电泳仪 北京市六一仪器厂;50BS-Ⅱ型生化培养箱 上海新苗医疗器械制造有限公司;SW-CT-2D型双人单面超净工作台 苏州苏洁净化设备有限公司;T100 Bio-Rad伯乐 PCR仪 美国Bio-Rad公司;BIO-RAD Gel Doc XR+凝胶成像分析系统 美国Bio-Rad公司。
1.2 实验方法
1.2.1 产蛋白酶嗜热菌的筛选 用无菌过滤器滤过实验水样,将过滤的滤纸直接加入富集培养基,55 ℃摇床培养24 h。再将富集培养液稀释成10-1、10-2、10-3、10-4、10-55个梯度,选择10-3、10-4、10-5三个梯度,取100 μL涂布于脱脂牛奶培养基上,每个梯度3个平行,置于55 ℃培养48 h。观察菌落形状、大小以及是否产透明圈。以不同菌落特征和有透明圈的菌株作为目标菌株进一步划线分离。经过3次划线分离纯化单菌株,4 ℃保藏于斜面。测量不同菌株的透明圈直径(D)和菌落直径(d),根据D/d值作为菌株产蛋白酶能力强弱的初步判断。
1.2.2 产蛋白酶嗜热菌的生长特性的研究
1.2.2.1 菌体生长的最适温度 将23株菌种按3%接种量接种于液体培养基中,培养温度设置为25、35、45、55、65 ℃,150 r/min恒温摇床培养48 h,测其OD600值,得出最适温度。
1.2.2.2 菌体生长的最适pH 将液体培养基初始pH调至5、6、7、8、9,把23株菌种按3%接种量接种于液体培养基中,55 ℃,150 r/min恒温摇床培养48 h,而后测其OD600值,得出最适pH。
1.2.3 产蛋白酶嗜热菌株形态观察和部分菌株生理生化鉴定
1.2.3.1 形态观察 对纯化的菌株进行形态观察记录和革兰氏染色,镜检观察。
1.2.3.2 生理生化鉴定 根据《常见细菌系统鉴定手册》[12]对代表菌株进行生理生化鉴定,主要包括接触酶、海藻酸钠降解、硝酸盐还原、甲基红(M-R实验)、V-P、柠檬酸盐利用、酪素水解、淀粉水解、糖醇发酵(葡萄糖、乳糖、D-木糖、D-果糖、D-甘露糖、麦芽糖、L(+)-阿拉伯糖)、硫化氢、耐盐(氯化钠的梯度分别为2%、5%、7%、10%)、吲哚和Tween80降解实验。
1.2.4 产蛋白酶嗜热菌株的16S rRNA基因分子鉴定 用Ezup柱式细菌基因组提取试剂盒提取23株产蛋白酶嗜热菌株DNA(具体方法见试剂盒使用说明书)。以总DNA为模板,采用文献[13]中的细菌16S rDNA通用引物和50 μL反应体系进行PCR扩增。扩增条件:94 ℃预变性4 min;94 ℃变性1 min;56 ℃退火温度,复性45 s;72 ℃延伸1.5 min;共30个循环;最终72 ℃延伸10 min。扩增产物在电压95 V,1.0%琼脂糖凝胶下电泳。凝胶成像系统检测PCR 扩增结果,送至上海生工生物工程有限公司进行测序。将测序并拼接的菌株16SrRNA基因序列提交GenBank中进行BLAS对比,利用MEGA6.0 中邻接法(Neighbour-joining methods,NJ)构建进化树,进化树分支置信度采用Bootstrap法(支重复次数为1000)。
1.3 数据处理
三次测定数据采用Excel进行数据分析
2 结果与分析
2.1 产蛋白酶嗜热菌的筛选与特征
通过富集和筛选,以产蛋白酶菌株在脱脂牛奶培养基可产生透明圈为依据,从宜春温泉温汤古井水样中筛选到29株产蛋白酶菌株,编号为:N1~N29,根据D/d比值越大产蛋白酶能力越强,选取D/d值>1.0的23株产蛋白酶嗜热菌菌株进行菌落形态观察、革兰氏染色和生长特性的研究,结果见表1。从表1中可见,大多数菌株的菌落特征呈现为湿润、乳白、椭圆,均为革兰氏染色阳性菌,在培养20 h内多数没有产芽孢。这23株菌生长pH范围为pH5.0~9.0,生长的温度范围较广,为25~65 ℃,除N27外,其余菌株最适生长温度为45~55 ℃,18株菌最高生长温度在55~65 ℃,其中N2、N4、N10、N18、N19、N20在65 ℃能明显生长,视为兼性嗜热菌[14]。
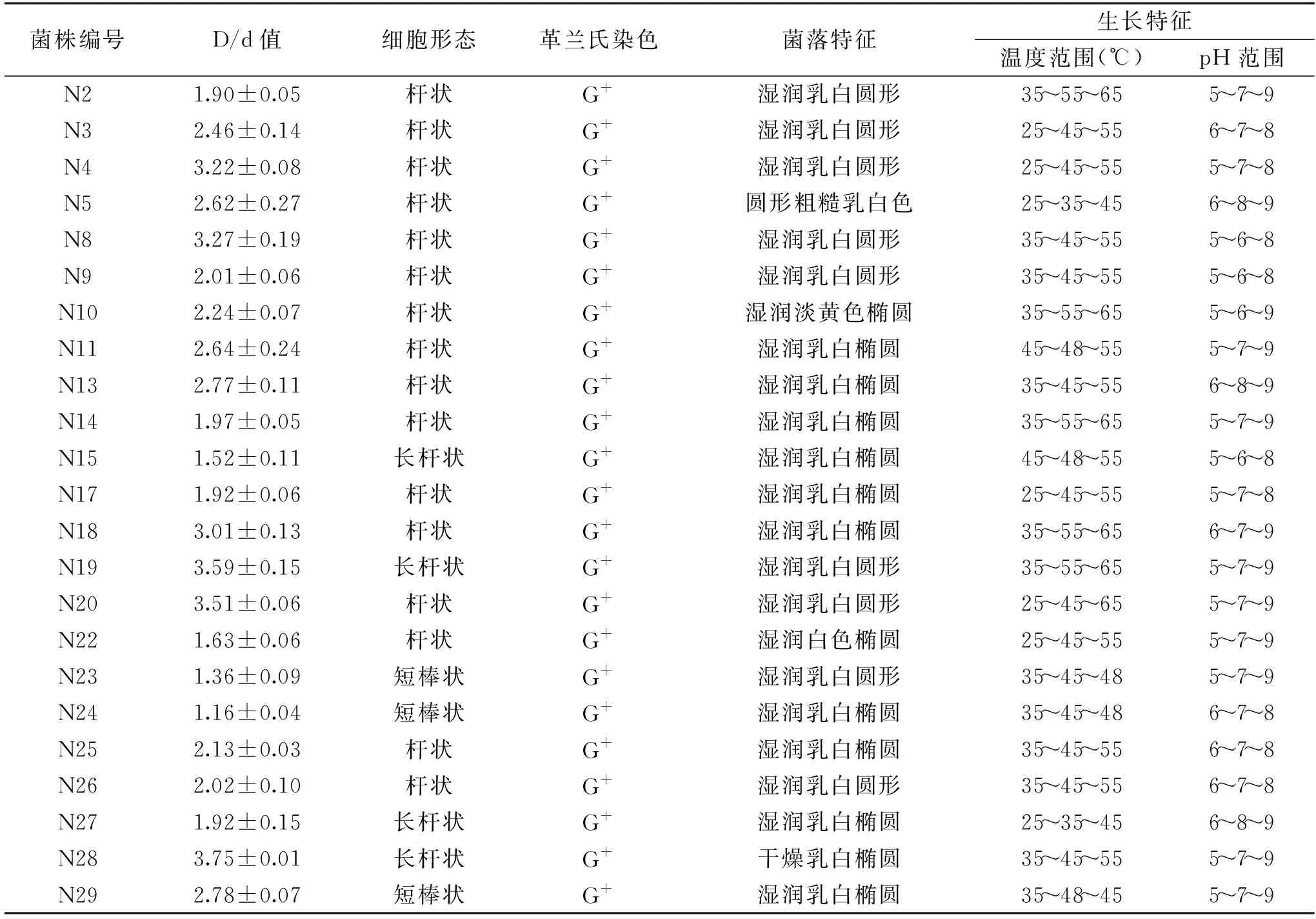
表1 产蛋白酶嗜热菌的筛选结果与特征Table 1 Screening results and characteristics of producing-protease thermophiles
2.2 生理生化特征
选取N2、N5、N9、N11、N17、N20为代表菌株进行生理生化鉴定,结果如表2所示,这6株代表菌株均不产气,对海藻酸钠实验、M-R实验、乳糖产酸实验、吲哚实验、Tween80 降解实验均为阴性,接触酶、葡萄糖产酸为阳性,可水解酪蛋白和淀粉(除N20不水解淀粉外),其余各项6株菌结果有差异,详见表2。
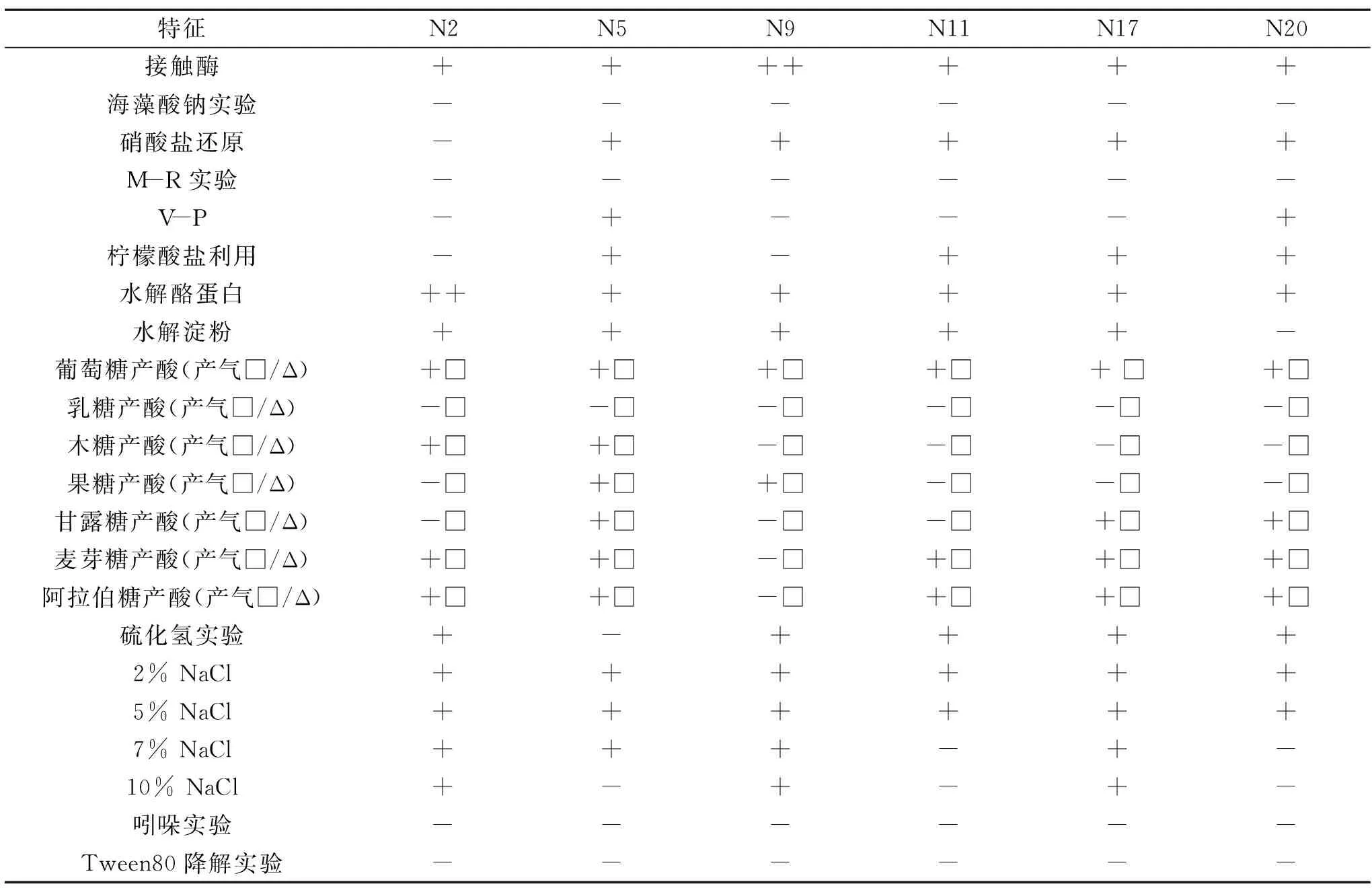
表2 部分产蛋白酶嗜热菌的生理生化结果Table 2 Physiological and biochemical results of the representative strains
2.3 16S rRNA的基因序列与系统发育分析
23株产蛋白酶嗜热菌菌株,经过总DNA提取、PCR扩增,结果见图1。由见图1可见均获得约1500 bp大小的目的片段,经双向测序并拼接后,将各序列提交到NCBI上的GeneBank数据库,获得登录号:KY433860-KY433881,经Blast比对,获得23株产蛋白酶嗜热菌菌株近缘种相似种属(见表3)。
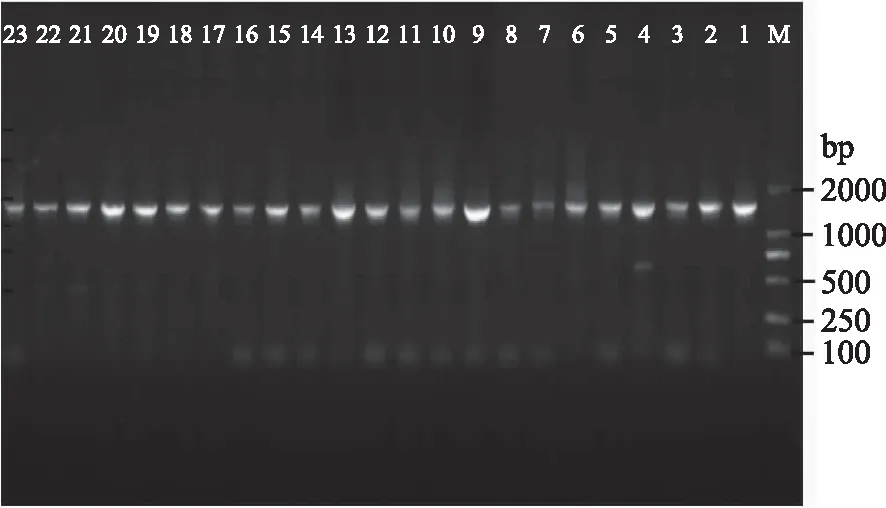
图1 23株菌株16S rRNA基因的PCR电泳结果Fig.1 PCR results of 16S rRNA genes PCR amplified of 23 strains注:M为DL2000标准品;1~23泳道依次为 N2、3、4、5、8、9、10、11、13、14、15、17、 18、19、20、22、23、24、25、26、27、28、29菌株。
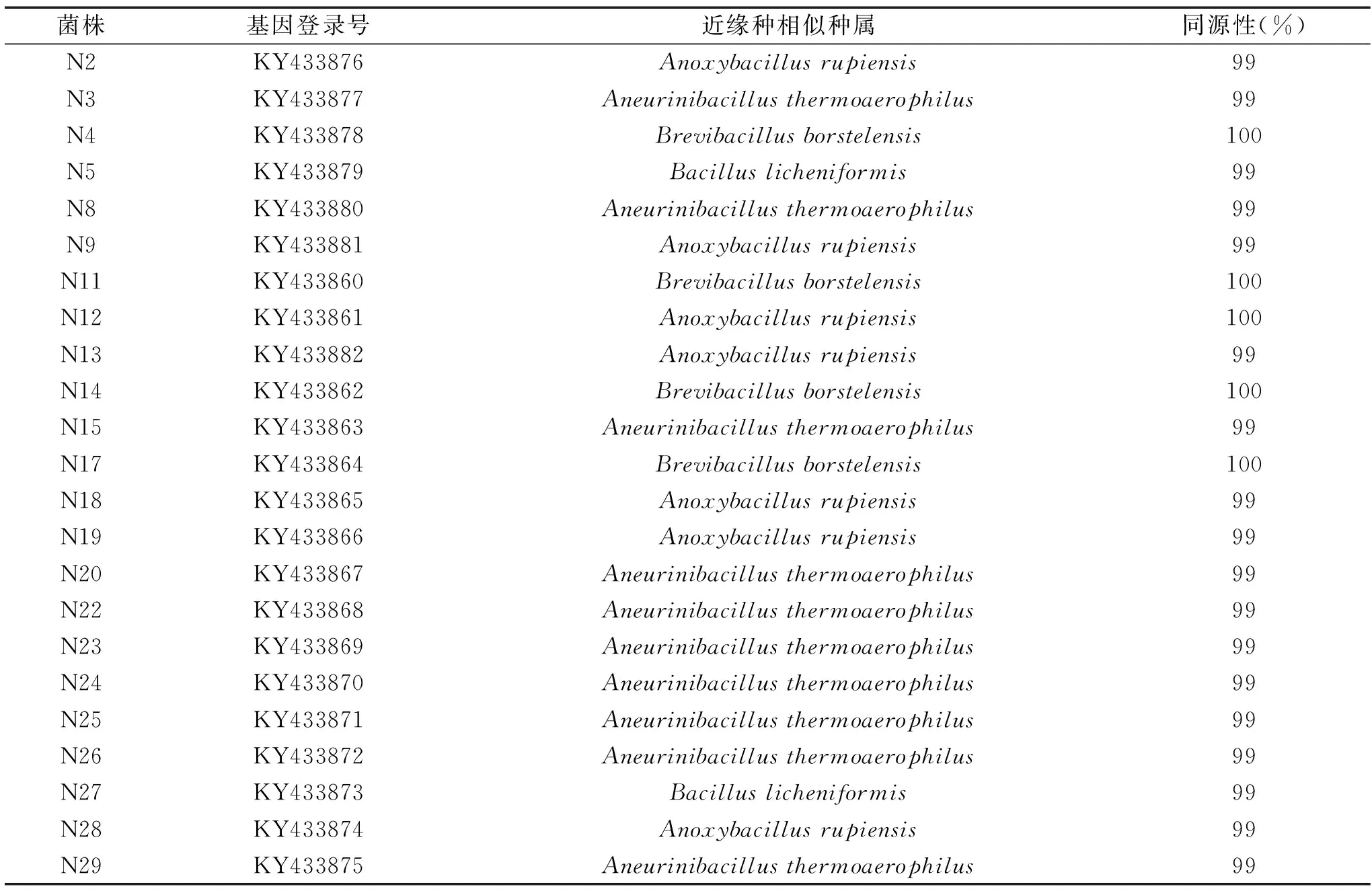
表3 23株菌株的同源性分析Table 3 Homology analysis of 23 strains
用MEGA6.0 中邻接法(Neighbour-joining methods,NJ)构建系统进化树如图3,由Blast比对和系统进化树可知23株产蛋白酶菌株可分别属于厌氧芽孢杆菌(Anoxybacillus)、硫胺素芽孢杆菌属(Aneurinibacillus)、短芽孢杆菌属(Brevibacillus)和芽孢杆菌属(Bacillus)四个属。N5、N27与Bacilluslicheniformis(CP014842.1)聚为一支,且与Bacilluslicheniformis同源性为99%,结合N5生理生化特征,可初步鉴定N5、N27为地衣芽孢杆菌(Bacilluslicheniformis)。4株短芽孢杆菌属(Brevibacillus)中,N11虽另聚为一支,但在生理生化上与N17基本一致,且N4、N11、N14、N17与Brevibacillusborstelensis同源性很高,均100%,初步认为这4株菌为Brevibacillusborstelensis。厌氧芽孢杆菌属(Anoxybacillus)7株菌株与Anoxybacillusrupiensis同源性为99%,N9、N13、N18、N19、N28和Anoxybacillusrupiensis聚为一支,可以认为同一种,而N2、N12则与Anoxybacillusamylolyticus聚为一支,初步鉴定为Anoxybacillussp,其余10株与Aneurinibacillusthermoaerophilus同源性为99%,(除N8外)均与Aneurinibacillusthermoaerophilus聚为一支,可初步鉴定为嗜热气硫胺素芽孢杆菌(Aneurinibacillusthermoaerophilus),占全部筛选菌株总数的39.1%。N8与N11聚集为一支,其分类单元有待于进一步研究。

图2 基于16SrRNA基因序列的产蛋白酶嗜热菌菌株系统发育树Fig.2 Phylogenetic tree of the Producing-protease thermophile based on 16S rRNA gene sequence
3 结论与讨论
本文从富硒温泉水样中筛选到23株产蛋白酶嗜热菌,这23株菌菌株生长pH范围为5.0~9.0,生长的温度范围较广,为25~65 ℃,除N27外,其余菌株最适生长温度为45~55 ℃,18株菌最高生长温度在55~65 ℃,其中N2、N4、N10、N18、N19、N20在65 ℃能明显生长,可视为兼性嗜热菌。通过生理生化和16S rRNA基因序列分析,结果表明,这些菌株分别属于厌氧芽孢杆菌属(Anoxybacillus)、硫胺素芽孢杆菌属(Aneurinibacillus)、芽孢杆菌属(Bacillus)和短芽孢杆菌属(Brevibacillus)4个属。其中2株(N5、N27)为地衣芽孢杆菌(Bacilluslicheniformis)。4株(N4、N11、N14、N17)为Brevibacillusborstelensis。7株厌氧芽孢杆菌属(Anoxybacillus)菌株中,N9、N13、N18、N19、N28初步鉴定为Anoxybacillusrupiensis,而N2、N12初步鉴定为Anoxybacillussp,剩余的10株(除N8外)均嗜热气硫胺素芽孢杆菌(Aneurinibacillusthermoaerophilus),占全部分离菌株总数的39.1%,为宜春富硒温泉水样中产蛋白酶嗜热菌的优势菌群。N8分类单元有待于进一步研究。
产蛋白酶嗜热菌株多见于芽孢杆菌属(Bacillus),已报导的有芽孢杆菌(Bacillussp. DPE_7)[8]、嗜热脂肪芽孢杆菌(Bacillusstearothermophilus)[15]、嗜热芽孢杆菌(ThermophilicBacillus)[16]、短小芽孢杆菌(Bacilluspumilus)[17],莫海威芽孢杆菌(Bacillusmojavensis)[18]等,本文2株地衣芽孢杆菌(Bacilluslicheniformis)为产蛋白酶嗜热菌株,这与刘唤明[19]报导的一致。
目前国内外报道的高温产蛋白酶的菌株中,短芽孢杆菌属(Brevibacillus)报道也不多,廉立慧[9]从徂徕山温泉附近土样中分离到一株产嗜热蛋白酶的菌株L7,首次报道该菌株为Brevibacillus属。龚志婷[20]报道腾冲温泉产蛋白酶的菌株主要分布在Geobacillus、Brevibacillus及Aneburinibacillus三属。本研究中筛选到4株Brevibacillus和9株Aneurinibacillus二属,未见Geobacillus属的产蛋白酶的嗜热菌株。而本文还获得7株Anoxybacillus属的菌株,Anoxybacillussp.在2000年作为新的种属首次报道[21],近年来,在我国云南腾冲温泉土壤中相继筛选到厌氧芽孢杆菌属(Anoxybacillus)产耐热普鲁兰酶菌株Anoxybacillussp.LM 18-11[22]和产ɑ-淀粉酶的菌株Anoxybacillussp.GXS-3[23],国内还未见有温泉环境中厌氧芽孢杆菌属(Anoxybacillus)产蛋白酶嗜热菌的报道,可见宜春富硒温泉有着丰富的产蛋白酶嗜热菌资源,为高温蛋白酶的开发和应用奠定基础。
