青枯雷尔氏菌III型分泌蛋白致病侵染的生物信息学初步分析
2018-07-06满娇,张武
满 娇,张 武
(大理大学 农学与生物科学学院,云南 大理 671003)
青枯病是由青枯雷尔氏菌引起的一种植物细菌性疾病,可侵染许多重要经济作物(如马铃薯、花生、番茄、烟草、甘薯、茄子等)以及一些贵重药材和花卉,特别是对茄科植物的危害更大[1]。青枯病是世界上危害最大、分布最广和造成损失最为严重的植物病害之一,该病的发生、传播和防治一直是国内外研究的重点和热点。
青枯雷尔氏菌作为革兰氏阴性菌的一种,可通过分泌效应物[2](effectors,专指病原菌分泌到宿主植物体内的分泌蛋白)引起致病侵染,研究其分泌机制对于病原菌的致病机理具有重要意义[3]。分泌蛋白是由细胞合成并分泌到质膜外的蛋白质,分为两大类,一类是经典分泌蛋白(classical secreted protein),该类分泌蛋白的N-端含有16~26个疏水性氨基酸残基组成的信号肽;另一类为非经典分泌蛋白,这类分泌蛋白缺少信号肽,III型分泌系统(type III secretion systems,T3SS)产生的分泌蛋白即为信号肽非依赖性。III型分泌蛋白作为重要的效应物蛋白,在许多植物-病原微生物互作过程中发挥极其关键的作用[4]。
分泌蛋白的研究可以用实验检测和理论预测两种方法。常规的蛋白质实验检测可以用western blot和Elisa(酶联免疫吸附测定),而Matthew等使用MALDI-TOF质谱仪,通过对细菌蛋白质组特异性修饰,可以直接从菌落中检测细菌的分泌蛋白,甚至识别病原菌的毒力和侵染蛋白质[5]。近年来,随着生物信息学和第三代单分子测序的迅猛发展,利用已有的蛋白质数据库去预测和分析分泌蛋白,进而通过实验验证已成为蛋白质组研究的常用手段,可以大大减轻繁琐的实验工作量。本文对青枯雷尔氏菌III型分泌蛋白的预测及分析,将有助于深入研究病原菌和宿主植物互作的分子机制。
1 材料与方法
1.1 材料
本实验材料是青枯雷尔氏菌全蛋白质组的5 116个蛋白质,氨基酸序列由NCBI数据库获得。青枯雷尔氏菌基因组大小约5.8 Mb,是由3.7 Mb的环形染色体和2.1 Mb的大质粒所组成,具有高GC含量。
1.2 方法
EffectiveDB(http://www.effectors.org/)是一个预测III型分泌系统的工具,主要功能是预测蛋白质是否具有III型分泌特征。EffectiveCCBD预测是否有伴侣蛋白结合在分泌蛋白25~70位氨基酸结合位点上,伴侣蛋白的目的是提高III型分泌蛋白在分泌前及分泌时的稳定性,这是III型分泌系统所产生的分泌蛋白的特别属性。
本文首先用EffectiveDB和EffectiveCCBD筛选出含有III型分泌系统的分泌蛋白,其次分别使用信号肽预测软件SignalP4.1(http://www.cbs.dtu.dk/services/SignalP/)和非经典分泌蛋白预测软件SecretomeP2.0(http://www.cbs.dtu.dk/services/SecretomeP/)剔除掉可能含有信号肽的分泌蛋白,再结合结构域预测软件SMART、ExPASy在线分析III型分泌蛋白的结构、功能并进行常规特征描述。
2 结果与分析
2.1 III型分泌蛋白的预测
利用EffictiveDB、EffectiveCCBD、SignalP4.1和SecretomeP2.0对青枯雷尔氏菌的5 116个蛋白质氨基酸序列进行预测和分析,共得到156个III型分泌蛋白,约占蛋白质总数的3.05%。
2.2 III型分泌蛋白的功能预测及分析
经过预测后,对其功能进行初步分析,发现在156个III型分泌蛋白中,75个为有功能描述的蛋白质,81个为假定蛋白(hypothetical protein)。图1为156个蛋白质的功能分类。
细菌T3SS分泌的效应蛋白大致可分为4类[4]:第一类为细胞质膜上的分泌系统自身组装蛋白;第二类为转位子蛋白,主要能在细胞膜上形成跨膜通道,使效应蛋白通过;第三类为分泌的效应蛋白,这是T3SS产生的关键效应物,这类蛋白与病原微生物的致病侵染密切相关,被转移进入寄主细胞引起相应的病理变化;第四类为T3SS 伴侣蛋白,与特异的效应蛋白结合,保护效应物在未分泌前不被降解,并能有效地分泌、转移效应蛋白。前两大类蛋白质,一共有31个(图1B),约占整个III型分泌蛋白的20%,由此可见,细菌T3SS在组装自身分泌装置方面消耗了大量的能量与精力。在有功能描述的分泌蛋白中,编码III型分泌系统的许多结构基因与编码细菌的鞭毛装置基因具有一定的同源性[6],鞭毛蛋白在细菌表面组装鞭毛装置,可能同分泌蛋白运输到宿主体内有关。丝氨酸蛋白酶是一类以丝氨酸为活性中心而命名的蛋白水解酶,在真菌中丝氨酸蛋白酶与病原菌的侵染密切相关,通常用来降解宿主植物体内的蛋白质[7],这说明丝氨酸蛋白酶可能与青枯雷尔氏菌的侵染相关。
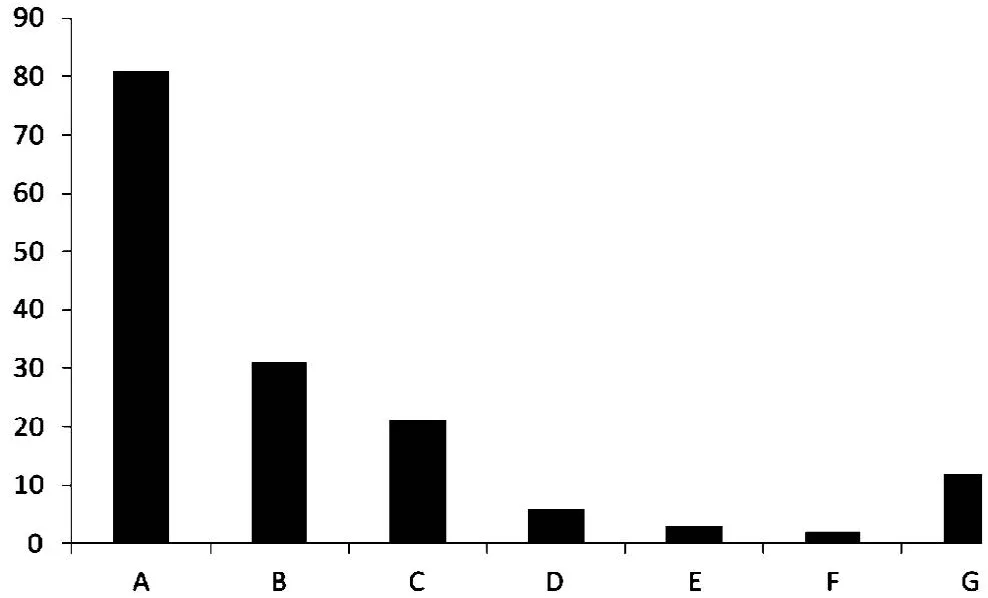
图1 青枯雷尔氏菌III型分泌蛋白功能分类
在其它III型分泌蛋白中,还发现了青霉素结合蛋白和热休克蛋白,这两种蛋白质在其它微生物中广泛存在[8],是病原微生物在逆境条件下抗胁迫的分子基础,可以扩大细菌在不良环境中的耐受性和生存范围。在本次预测中还发现了AvrA蛋白,蔺志杰[9]发现在肠炎沙门菌中AvrA效应蛋白为抑制炎症分子,在肠炎沙门菌感染宿主过程中可抑制宿主细胞的过度炎性反应,推测此种III型分泌蛋白在青枯雷尔氏菌中可能也有抑制宿主植物过敏反应的功能。HrpB和HrcU (hypersensitive response,HR,过敏反应)主要功能是抑制宿主植物的基础抗性或者防卫反应,调控其他T3SS相关基因的表达。另外,通过位置分析,发现HrcU和HrpB等4个III型分泌蛋白的基因集中位于青枯雷尔氏菌的大质粒上,有可能形成一个“毒力岛”,这与很多病原微生物的毒力蛋白基因分布是高度一致的[10-11]。表1是部分青枯雷尔氏菌III型分泌蛋白的常规特征描述,由 ExPASy(http://web.expasy.org/protparam/)在线分析完成。
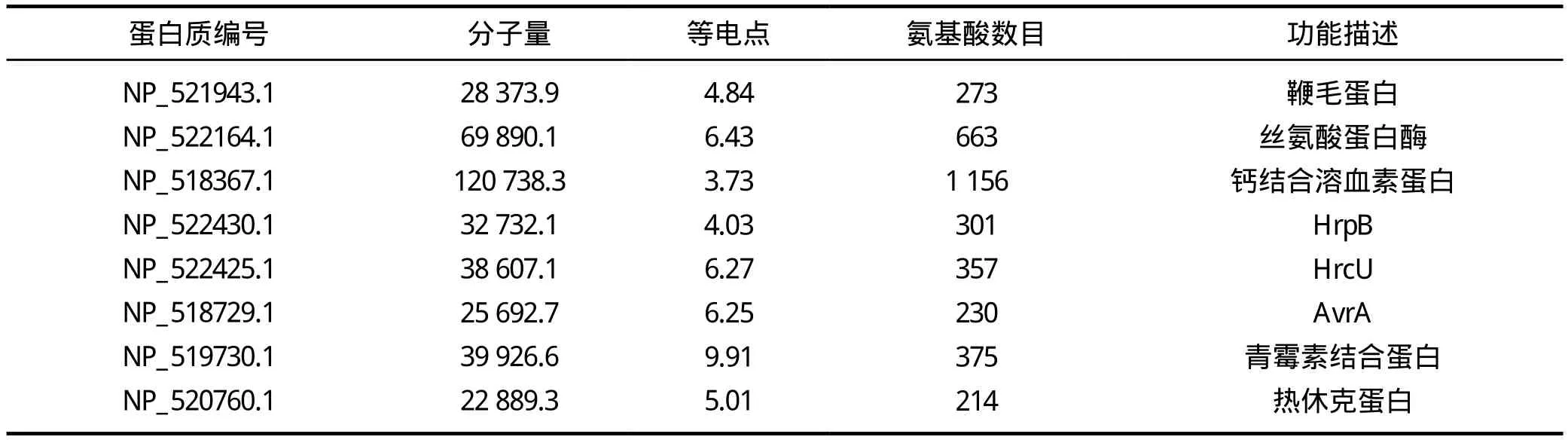
表1 部分典型III型分泌蛋白特征描述
为了更好的分析III型分泌假定蛋白在青枯雷尔氏菌中的功能,我们用SMART软件进行结构域分析。Hemolysin Cabind是在假定蛋白中含量最多的一种结构域,包括HlyD结构域,这两种结构域功能与表1中钙结合溶血素蛋白功能类似,是一种钙结合细菌溶血素蛋白,含有此类结构的分泌蛋白可能在钙离子的介导下进入到宿主植物体内,与细菌产生毒力和致病性有关(Hemolysin Cabind等结构域功能在欧洲分子生物学实验室EMBL-EBI网站的PFAM中查询);假定蛋白中的Flagellin_N结构域与鞭毛组装有关,可能参与青枯雷尔氏菌T3SS系统的组装。
5 结论
本文主要利用生物信息学工具来预测和分析青枯雷尔氏菌的III型分泌蛋白及其功能,结果发现,III型分泌蛋白作为重要的致病因子,除部分蛋白质功能未知外,还包括有外膜/孔道蛋白、鞭毛蛋白、抗应激蛋白、丝氨酸蛋白酶、钙结合溶血素蛋白等类型,主要参与了鞭毛合成、毒力侵染等过程。
目前,由青枯雷尔氏菌引起的病害是灾难性的,所以,对其基因组和蛋白质组进行生物信息学分析、掌握分泌蛋白的致病侵染的机制非常必要。本文改进的方法是结合实验检测方法,可以提取青枯雷尔氏菌的分泌蛋白进行分离纯化和分析鉴定,且在代谢组水平上全面分析基因的功能和彼此形成的代谢调控网络。了解青枯雷尔氏菌在寄主植物体内的致病机理,对于青枯病的控制具有重要的现实意义。
[1]车建美,林海云,刘波,等.插入序列ISRso21在青枯雷尔氏菌中的分布以及插入位点多态性分析[J].农业生物技术学报,2014(11):1424-1433.
[2]王蔚淼,潘玲.细菌Ⅲ型分泌系统的研究进展[J].畜牧与兽医,2006(6):58-60.
[3]张勇,李牧原,罗锋.青枯菌三型分泌系统研究进展[J].微生物学报,2015(6):675-682.
[4]朱秀秀,高必达,赵廷昌,等.植物病原细菌Ⅲ型分泌系统及Pseudomonas syringae pv.tomato的信号分子分泌研究进展[J].湖南农业科学,2009(2):19.
[5]Matthew MC,Emily AW,George MK,et al.Direct detection of bacterial protein secretion using whole colony proteomics[J].Molecular & cellular proteomics,2012(9):596-604.
[6]He SY.Type Ⅲ protein secretion system in plant and animal pathogenic bacteria[J].Annu Rev Phytopathol,1998,36:363-392.
[7]Monod M,Capoccia S,Léchenne B,et al.Secreted proteases from pathogenic fungi[J].International Journal of Medical Microbiology,2002,292:405-419.
[8]张武,马金田,潘伟,等.菜豆根瘤菌Rhizobium etli CFN42全基因组含信号肽分泌蛋白的鉴定与分析[J].基因组学与应用生物学,2014(5):961-969.
[9]蔺志杰.肠炎沙门菌效应蛋白AvrA参与宿主炎性反应机制[D].杨州:扬州大学,2017.
[10]沈禹颐.病原性细菌毒力岛的研究进展[J].中国兽医科技,2003(3):41-42.
[11]尹磊,祁克宗,宋祥军,等.大肠杆菌Ⅲ型分泌系统2毒力岛研究进展[J].微生物学通报, 2017(12):3031-3037.
