PTEN调控STAT3信号通路影响心肌成纤维细胞增殖的研究
2018-07-06万敏娜金钟奎
万敏娜 金钟奎 汤 澄
(宜春市人民医院心血管内科,宜春 336000)
糖尿病心肌病是一种慢性疾病,是独立于糖尿病以外的并发症,主要以心肌纤维化、胶原纤维沉积为主要特征,心肌纤维化的主要原因是成纤维细胞过度增殖,细胞外基质大量合成,远远超出降解的速度[1,2]。第10号染色体同源缺失性磷酸酶-张力蛋白(Phosphatase and tensin homolog deleted on chromosome 10,PTEN)是一种与细胞生长和凋亡等有关的基因,在恶性肿瘤、肺组织纤维化等疾病中具有调控作用,基因敲除PTEN后小鼠心脏和体重之间比例增加,心肌纤维化程度加重,心肌收缩能力降低[3,4]。心肌成纤维细胞表达PTEN,在醛固酮等致病因子的作用下其表达水平下调,PTEN在糖尿病心肌病大鼠模型的心肌组织中表达水平低于正常大鼠[5,6]。信号转导与转录因子3(Signal transducers and activators of transcription 3,STAT3)广泛存在于哺乳动物组织器官中,在细胞生长、疾病发生中具有关键作用,STAT3磷酸化水平升高后,可以促进细胞生长,抑制STAT3信号通路后的心肌成纤维细胞增殖能力[7,8]。本研究旨在探讨PTEN对高糖诱导心肌成纤维细胞增殖的影响,以期为研究糖尿病心肌病的发病机制提供参考。
1 材料与方法
1.1材料 24 h内出生的乳鼠由江西中医药大学提供;PTEN过表达载体(pSicoR-PTEN)由江西中医药大学实验室构建保存;PTEN、甘油醛-3-磷酸脱氢酶(Glyceraldehyde-3-phosphate dehydrogenase,GAPDH)引物由上海生工合成;STAT3通路阻断剂AG490为美国MCE产品;RNA提取试剂盒、二喹啉甲酸(Bicinchoninic acid,BCA)蛋白定量试剂盒为德国QIAGEN产品;Ⅱ型胶原酶、胰蛋白酶为美国Sigma产品;STAT3一抗、磷酸化的STAT3(p-STAT3)一抗、GAPDH一抗为美国Santa Cruz产品;DAB显色试剂盒为北京Solarbio产品;cDNA合成试剂盒为大连TaKaRa产品;qRT-PCR试剂盒为美国Roche产品;Lipofectamine2000为美国Thermo产品。
1.2方法
1.2.1心肌成纤维细胞分离培养 心肌成纤维细胞分离培养参照文献[9],将24 h内出生的乳鼠用酒精消毒后,取心脏,放在4℃的磷酸缓冲盐溶液(Phosphate buffered saline,PBS)中洗涤后,加入75%的乙醇溶液,把血细胞去除后,剪碎,加入1倍体积的0.1%的Ⅱ型胶原酶和0.125%胰蛋白酶,放在37℃的水浴中消化,期间每隔10 min摇晃一次,收集消化液,加入含有10%胎牛血清的DMEM混合后,种植到细胞培养瓶中,放在37℃,5%CO2培养箱中培养。心肌细胞和心肌成纤维细胞的贴壁时间差为60 min。将上清液吸除后,加入新鲜细胞培养液,继续培养。用0.25%的胰蛋白酶消化传代。取第3代细胞用于实验。
1.2.2qRT-PCR检测高糖作用后细胞中PTEN表达水平 取第3代心肌成纤维细胞,分别用含有5.5 mmol/L的D葡萄糖和30 mmol/L的D葡萄糖的细胞培养液培养,记为Control和HG。24 h后,加入Trizol,每个6孔板孔中加入1 ml的Trizol,充分裂解后。加入1/5体积(200 μl)三氯甲烷,充分混合后,4℃,12 000 r/min离心15 min。吸取上清溶液,加入等体积的异丙醇,在室温条件下放置10 min,离心,把上清弃掉,用适量的75%乙醇洗涤后,晾干。用DEPC水溶解以后,紫外分光光度计检测浓度、纯度。反转录合成cDNA,qRT-PCR检测试剂盒检测PTEN的水平,内参基因为GAPDH,ΔΔCt法计算PTEN水平。PTEN上游引物:5′-TGACAGCCATCATGAAAGA-3′,下游引物:5′-CCGCTCTCATCAAAAGGTTCATTC-3′。GAPDH上游引物:5′-AAGGTGAAGGTCGGAGTCAAC-3′,下游引物:5′-GGGGTCATTGATAACAACAATA-3′。实验重复3次,取均值。
1.2.3Western blot检测高糖作用后细胞中PTEN表达水平 按照1.2.2中方法培养收集Control和HG细胞,加入细胞裂解液,在冰上裂解50 min,4℃,16 000 r/min离心15 min,蛋白存在于上清中,用BCA蛋白浓度检测试剂盒对蛋白浓度进行定量检测。蛋白样品与5×上样缓冲液按照4∶1的比例混匀后,100℃孵育5 min。每孔中加入40 μg蛋白,SDS-PAGE电泳初始电压为80 V,电泳0.5 h以后,用120 V 继续电泳。90 V,4℃转膜90 min。5%牛血清白蛋白在室温封闭60 min。加入1∶900稀释的一抗4℃过夜后,加入1∶3 000稀释的辣根过氧化物标记的二抗室温反应1 h,DAB显色,在暗室中曝光,Quantity one分析条带灰度值,以目的条带灰度值/GAPDH灰度值表示蛋白表达水平。实验重复3次,取均值。
1.2.4细胞转染及分组 心肌成纤维细胞培养至对数期后,把细胞浓度调整到每毫升含有2×105个细胞,种植到6孔细胞培养板中,观察细胞融合度约为60%进行细胞转染。将pSicoR-PTEN和对照pSicoR用Lipofectamine2000转染至细胞中,用含有5.5 mmol/L的D葡萄糖的细胞培养液培养,记为PTEN和NC。转染pSicoR-PTEN后的细胞经含有30 mmol/L的D葡萄糖细胞培养液培养后记为PTEN+HG。Control和HG细胞处理方法同1.2.2。
1.2.5MTT检测细胞增殖 Control、HG、PTEN+HG细胞以每孔加入5 000个细胞种植到96孔细胞培养板。培养24 h以后,每孔中加入20 μl的浓度为5 mg/ml的噻唑蓝(Methylthiazolyldiphenyl-tetrazolium bromide,MTT),在37℃孵育4 h。每组设置5个复孔。将上清吸除以后,每孔中加入二甲基亚砜溶液150 μl,避光,室温反应10 min。用酶标仪检测每孔490 nm的A值。实验重复3次,取均值。
1.2.6Western blot检测STAT3、p-STAT3水平 Control、HG、PTEN+HG细胞培养24 h后,Western blot检测细胞中STAT3、p-STAT3水平,STAT3、p-STAT3一抗以1∶800和1∶500稀释。步骤同1.2.3。实验重复3次,取均值。
1.2.7STAT3通路阻断剂AG490对细胞增殖影响 转染pSicoR-PTEN后的心肌成纤维细胞用含有30 μmol/L的STAT3通路阻断剂AG490和含有30 mmol/L的D葡萄糖的细胞培养液培养记为PTEN+HG+AG490。 PTEN+HG、PTEN+HG+AG490细胞培养24 h后,MTT检测细胞增殖,Western blot检测细胞中STAT3、p-STAT3水平。步骤同1.2.3和1.2.5。实验重复3次,取均值。
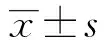
2 结果
2.1高糖作用后心肌成纤维细胞中PTEN水平 如图1和表1所示,Control、HG细胞中PTEN mRNA水平依次为:1.00±0.10、0.26±0.03,PTEN蛋白水平为:1.08±0.11、0.34±0.04。HG细胞中PTEN mRNA水平和蛋白水平均明显低于Control,差异具有统计学意义(t1=42.724,t2=10.951,P<0.05)。高糖作用后的心肌成纤维细胞中PTEN表达水平下降。
2.2细胞转染后PTEN表达水平 结果如图2和表2所示,Control、NC、PTEN细胞中PTEN mRNA水平依次为:1.00±0.09、1.03±0.07、1.87±0.16,PTEN蛋白水平为:0.98±0.08、0.96±0.12、1.68±0.14。NC细胞中PTEN mRNA水平和蛋白水平与Control相比差异无统计学意义(t1=0.364,t2=0.211,P>0.05)。PTEN细胞中PTEN mRNA水平和蛋白水平明显高于Control,差异有统计学意义(t1=10.568,t2=7.388,P<0.05)。心肌成纤维细胞转染pSicoR-PTEN后能够提高细胞中PTEN基因和蛋白水平,转染对照载体pSicoR对细胞中PTEN基因和蛋白水平没有影响。

图1 Western blot检测高糖作用后细胞中PTEN水平Fig.1 Level of PTEN in cells after high glucose detected by Western blot
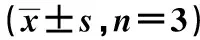

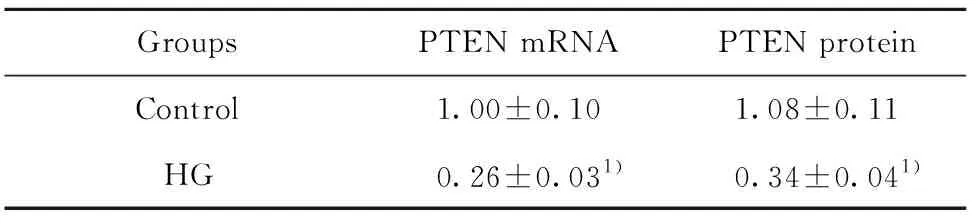
GroupsPTEN mRNAPTEN proteinControl1.00±0.101.08±0.11HG0.26±0.031)0.34±0.041)
Note:Vs control,1)P<0.05.
2.3PTEN对高糖诱导的细胞增殖影响 如表3所示,Control、HG、PTEN+HG细胞A值依次为:0.32±0.04、0.54±0.06、0.41±0.04。HG细胞A值与Control相比明显升高,差异具有统计学意义(t=5.660,P<0.05)。PTEN+HG细胞A值明显低于HG,差异具有统计学意义(t=3.344,P<0.05)。高糖诱导心肌成纤维细胞增殖,PTEN可以减弱高糖诱导的细胞增殖作用。
2.4PTEN对高糖环境细胞中STAT3、p-STAT3水平影响 如图3和表4所示,Control、HG、PTEN+HG细胞STAT3水平依次为:0.62±0.05、0.60±0.08、0.61±0.06,p-STAT3水平依次为:0.14±0.02、0.52±0.06、0.28±0.03。HG细胞p-STAT3水平与Control相比明显升高,差异具有统计学意义(t=11.516,P<0.05)。 PTEN+HG细胞p-STAT3水平明显低于
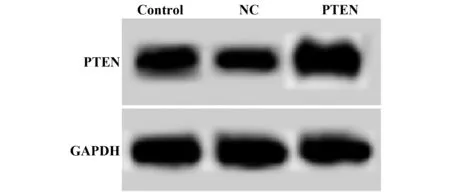
图2 Western blot检测转染后细胞中PTEN蛋白水平Fig.2 Level of PTEN protein in transfected cells detected by Western blot



GroupsPTEN mRNAPTEN proteinControl1.00±0.090.98±0.08NC1.03±0.070.96±0.12PTEN1.87±0.161)1.68±0.141)F71.97137.455P0.0000.000
Note:Vs control,1)P<0.05.


GroupsA valueControl0.32±0.04HG0.54±0.061)PTEN+HG0.41±0.042)F16.191P0.004
Note:Vs Control,1)P<0.05;vs HG,2)P<0.05.

图3 Western blot检测 PTEN对细胞STAT3、p-STAT3水平影响Fig.3 Effect of PTEN on cell STAT3,p-STAT3 level detected by Western blot

GroupsSTAT3p-STAT3Control0.62±0.050.14±0.02HG0.60±0.080.52±0.061)PTEN+HG0.61±0.060.28±0.032)F0.07267.837P0.9310.000
Note:Vs Control,1)P<0.05;vs HG,2)P<0.05.
HG,差异具有统计学意义(t=7.273,P<0.05)。高糖诱导心肌成纤维细胞中STAT3磷酸化,PTEN降低高糖环境细胞中STAT3磷酸化水平。
2.5抑制STAT3信号通路对细胞增殖影响 结果见图4和表5中所示,PTEN+HG、PTEN+HG+AG490细胞A值依次为:0.46±0.04、0.35±0.03,STAT3水平依次为:0.84±0.08、0.86±0.09,p-STAT3水平依次为:(0.23±0.02)、(0.11±0.01)。PTEN+HG+AG490细胞p-STAT3水平和A值明显降低,与PTEN+HG相比,差异具有统计学意义(t1=3.811,t2=9.295,P<0.05)。抑制STAT3信号通路可以进一步降低心肌成纤维细胞增殖能力,其与PTEN具有协同作用,可以抑制高糖诱导的心肌成纤维细胞增殖。

图4 Western blot检测各组细胞中STAT3、p-STAT3水平Fig.4 STAT3,p-STAT3 levels of cells in each group detected by Western blot

GroupsA valuep-STAT3STAT3PTEN+HG0.46±0.040.23±0.020.84±0.08PTEN+HG+AG4900.35±0.031)0.11±0.011)0.86±0.09
Note:Vs PTEN+HG,1)P<0.05.
3 讨论
人类PTEN基因定位于染色体10q23,其由8个内含子、9个外显子组成,其编码的蛋白质由403个氨基酸组成,在细胞浆、细胞核和细胞膜均有表达,其含有的PDZ结构域能够特异性的识别PDZ基序蛋白,从而调控三磷酸磷脂酰的活性,影响细胞内信号转导[10-12]。PTEN与心血管系统疾病发生有关,与心力衰竭、心肌收缩、心肌缺血再灌注、糖尿病心肌病等关,其编码的蛋白质具有蛋白和脂质磷酸酶活性,能够抑制细胞的生长[13,14]。研究显示,PTEN在心肌肥厚大鼠模型中表达上调,是一种抗心肌肥厚因子,基因敲除PTEN的小鼠心脏的外观变大,出现明显的心肌纤维化[15,16]。付兴根等[17]在研究螺内酯对糖尿病心肌病大鼠的影响时发现,糖尿病心肌病大鼠心肌组织中的PTEN mRNA和蛋白水平明显低于正常大鼠,PTEN参与糖尿病心肌病发生过程。本研究结果显示,高糖作用后的心肌成纤维细胞中PTEN水平下降,心肌成纤维细胞增殖能力增加,这与上述报道相符合,说明PTEN参与糖尿病心肌病的发生过程。
STAT3能够促进细胞增殖,在心力衰竭、缺血再灌注等中具有保护作用,在糖尿病心肌病中过度激活[18-20]。STAT3是STATs家族中的一员,而STATs家族与细胞增殖、分化、生长、凋亡等有关,是目前研究最多的信号通路之一,STAT3蛋白大小为89~92 kD,其含有多个结构域,如:DNA结合区域、螺旋区域、连接区域、C端转录活化区域和SH2结构域等,正是由于这些结构域,其在细胞正常生物学特性维持中具有重要作用[21-25]。STAT3在受到细胞外和细胞内的相关因子作用后,STAT3磷酸化,而2个磷酸化的STAT3可以形成二聚体后转移至细胞核内,发挥信号传递的作用[26-28]。STAT3与糖尿病心肌病有关,无论是在Ⅰ型糖尿病还是Ⅱ型糖尿病中均发现STAT3信号通路异常,并且与间质纤维化等有关,在高糖作用后的心肌成纤维细胞中发现p-STAT3水平升高[27,29]。PTEN和STAT3作为重要的调控细胞生长因子,二者之间也存在一定的联系[30]。
本研究结果显示,心肌成纤维细胞经高糖作用后,细胞中的STAT3磷酸化水平升高,而PTEN表达水平升高后,经高糖诱导后的细胞中STAT3磷酸化有所降低。进一步用STAT3信号通路抑制剂处理后发现,抑制STAT3信号通路和过表达PTEN具有协同作用,都能够抑制高糖诱导的心肌成纤维细胞增殖,说明PTEN通过抑制STAT3信号通路降低高糖诱导的心肌成纤维细胞增殖能力。
综上,高糖诱导心肌成纤维细胞增殖,降低细胞中PTEN表达水平,过表PTEN后可以降低高糖诱导的心肌成纤维细胞增殖水平,作用机制与抑制STAT3信号通路有关。本研究明确了PTEN在高糖诱导的心肌成纤维细胞增殖中的作用,为探讨糖尿病心肌病及心肌纤维化的发病机制奠定了基础。本研究存在一定的局限性,只在心肌成纤维细胞中进行了探讨,后续试验中会继续探讨PTEN在心肌细胞、平滑肌细胞等多种细胞中的作用。
参考文献:
[1] Huynh K,Bernardo BC,McMullen JR,etal.Diabetic cardiomyopathy:mechanisms and new treatment strategies targeting antioxidant signaling pathways[J].Pharmacol Therapeu,2014,142(3):375-415.
[2] Seferovic PM,Paulus WJ.Clinical diabetic cardiomyopathy:a two-faced disease with restrictive and dilated phenotypes[J].Europ Heart J,2015,36(27):1718-1727.
[3] Yu L,Li F,Zhao G,etal.Protective effect of berberine against myocardial ischemia reperfusion injury:role of Notch1/Hes1-PTEN/Akt signaling[J].Apoptosis,2015,20(6):796-810.
[4] Yang X,Qin Y,Shao S,etal.MicroRNA-214 inhibits left ventricular remodeling in an acute myocardial infarction rat model by suppressing cellular apoptosis via the phosphatase and tensin homolog(PTEN)[J].Int Heart J,2016,57(2):247-250.
[5] ingh B,Singh V,Krishnan A,etal.Regeneration of diabetic axons is enhanced by selective knockdown of the PTEN gene[J].Brain A J Neurol ,2014,137(4):1051-1067.
[6] 梁德贤,李庆军,陈康荣,等.糖尿病心肌PTEN蛋白表达变化及其在糖尿病心肌IPo中的影响[J].中国民族民间医药,2016,25(17):32-36.
Liang DX,Li QJ,Chen KR,etal.To investigate the changes of PTEN protein expression in diabetic myocardium and its effect on IPo in diabetic myocardium[J].Chin J Ethnomed Ethnopharm,2016,25 (17): 32-36.
[7] Li H,Yao W,Liu Z,etal.Hyperglycemia abrogates ischemic postconditioning cardioprotection by impairing AdipoR1/Caveolin-3/STAT3 signaling in diabetic rats[J].Diabetes,2016,65(4):942-955.
[8] Saarimäki-Vire J,Balboa D,Russell MA,etal.An activating STAT3 mutation causes neonatal diabetes through premature induction of pancreatic differentiation[J].Cell Rep,2017,19(2):281-294.
[9] Zhao X,Wang K,Liao Y,etal.MicroRNA-101a inhibits cardiac fibrosis induced by hypoxia via targeting TGFβRI on cardiac fibroblasts[J].Cell Physiol Biochem,2015,35(1):213-226.
[10] Wang S,Tie J,Wang R,etal.SOX2,a predictor of survival in gastric cancer,inhibits cell proliferation and metastasis by regulating PTEN[J].Cancer Letters,2015,358(2):210-219.
[11] Ren P,Gong F,Zhang Y,etal.MicroRNA-92a promotes growth,metastasis,and chemoresistance in non-small cell lung cancer cells by targeting PTEN[J].Tumor Biol,2016,37(3):3215-3225.
[12] Li WF,Ou Q,Dai H,etal.Lentiviral-mediated short hairpin RNA knockdown of MTDH inhibits cell growth and induces apoptosis by regulating the PTEN/AKT pathway in hepatocellular carcinoma[J].Int J Mol Sci,2015,16(8):19419-19432.
[13] 薛 锐,夏中元,孟庆涛.PTEN在神经系统损伤、心血管疾病发生发展中的作用研究进展[J].山东医药,2016,56(3):93-94.
Xue R,Xia ZY,Meng QT.Research Progress on the role of PTEN in the development of nervous system injury and cardiovascular disease[J].Shandong Med,2016,56 (3): 93-94.
[14] Pan J,Ling Y,Shang W,etal.Uric acid promotes cardiomyocyte apoptosis through regulating microRNA-21/PTEN/AKT/eNOS axis in myocardial ischemia/reperfusion injury[J].Int J Clin Exp Med ,2017,10(4):6303-6312.
[15] Roe ND,Xu X,Kandadi MR,etal.Targeted deletion of PTEN in cardiomyocytes renders cardiac contractile dysfunction through interruption of Pink1-AMPK signaling and autophagy[J].Biochim Biophys Acta,2015,1852(2):290-298.
[16] Gao Y,Chu M,Hong J,etal.Hypoxia induces cardiac fibroblast proliferation and phenotypic switch:a role for caveolae and caveolin-1/PTEN mediated pathway[J].J Thoracic Dis,2014,6(10):1458-1468.
[17] 付兴根,邢佑敏,韩 刚,等.螺内酯对糖尿病大鼠心肌PTEN表达的影响[J].中国现代医学杂志,2014,24(11):28-33.
Fu XG,Xing YM,Han G,etal.Effect of spironolactone on the PTEN expression in the myocardium of diabetic rats[J].Chin J Mod Med,2014,24(11): 28-33.
[18] Cen M,Hu P,Cai Z,etal.TIEG1 deficiency confers enhanced myocardial protection in the infarcted heart by mediating the Pten/Akt signalling pathway[J].Int J Mol Med,2017,39(3):569.
[19] Li CM,Shen SW,Wang T,etal.Myocardial ischemic post-conditioning attenuates ischemia reperfusion injury via PTEN/Akt signal pathway[J].Int J Clin Exp Med,2015,8(9):15801-15807.
[20] 薛 锐,夏中元,雷少青,等.PTEN在糖尿病因素削弱缺氧后处理对心肌细胞保护效应中的作用:与GSK-3β介导的线粒体凋亡途径的关系[J].中华麻醉学杂志,2017,37(1):108-111.
Xue R,Xia ZY,Lei SQ,etal.The role of PTEN in the protective effect of hypoxic treatment on cardiac myocytes in diabetic factors: the relationship with GSK-3β mediated mitochondrial apoptosis pathway[J].Chin J Anesthesiol,2017,37 (1): 108-111.
[21] Wang L,Li J,Li D.Losartan reduces myocardial interstitial fibrosis in diabetic cardiomyopathy rats by inhibiting JAK/STAT signaling pathway[J].Int J Clin Exp Pathol,2015,8(1):466-473.
[22] Das A,Salloum FN,Filippone SM,etal.Inhibition of mammalian target of rapamycin protects against reperfusion injury in diabetic heart through STAT3 signaling[J].Basic Res Cardiol,2015,110(3):1-11.
[23] 袁 超,管金川,曹 红,等.脊髓背角MCP-1-JAK2/STAT3信号转导参与大鼠2型糖尿病神经病理性痛的机制研究[J].中国病理生理杂志,2015,31(8):1467-1471.
Yuan C,Guan JC,Cao H,etal.Mechanism of MCP-1-JAK2/STAT3 signal transduction in spinal cord dorsal contributing to rats with type 2 diabetic neuropathic pain.[J].Chin J Pathophys,2015,31 (8): 1467-1471.
[24] 陈 云,杨 爽,王力宁,等.STAT3在糖尿病肾病肾小球系膜细胞自噬调控中的作用[J].中华肾脏病杂志,2016,32(10):772-778.
Chen Y,Yang S,Wang LN,etal.The role of STAT3 in the regulation of autophagy in glomerular mesangial cells of diabetic nephropathy[J].Chin J Nephrol,2016,32 (10): 772-778.
[25] Wang TS,Gao F,Qi QR,etal.Dysregulated LIF-STAT3 pathway is responsible for impaired embryo implantation in a Streptozotocin-induced diabetic mouse model[J].Biol Open,2015,4(7):893-902.
[26] Lo SH,Hsu CT,Niu HS,etal.Ginsenoside Rh2 improves cardiac fibrosis via PPARδ-STAT3 signaling in type 1-like diabetic rats[J].Int J Mol Sci,2017,18(7):1364.
[27] Wang X,Sun W,Shen W,etal.Long non-coding RNA DILC regulates liver cancer stem cells via IL-6/STAT3 axis[J].J Hepatol,2016,64(6):1283-1294.
[28] Sobotta MC,Liou W,Stöcker S,etal.Peroxiredoxin-2 and STAT3 form a redox relay for H2O2signaling[J].Nat Chem Biol,2015,11(1):64-70.
[29] 张 旭,陆 燕,王 琼,等.2型糖尿病大鼠STAT3酪氨酸磷酸化与IRS1、2及其丝氨酸磷酸化的关系[J].营养学报,2017,39(1):71-75.
Zhang X,Lu Y,Wang Q,etal.The relationship among tyrosine phosphorylation of STAT3,IRS1,IRS2 and their serine phosphorylation in rats with type 2 diabetic[J].ACTA Nutrimenta Sinica,2017,39 (1): 71-75.
[30] Moon SH,Kim DK,Cha Y,etal.PI3K/Akt and STAT3 signaling regulated by PTEN control of the cancer,stem cell population,proliferation and senescence in a glioblastoma cell line[J].Int J Oncol,2013,42(3):921-928.