miR-936对人喉癌细胞株Hep-2增殖、迁移、侵袭和药物敏感性的影响
2018-07-02王海峰李培王志远林惜君郭程叶进
王海峰 李培 王志远 林惜君 郭程 叶进
喉癌是头颈部常见的恶性肿瘤,其中喉鳞状细胞癌是其主要病理类型,占90%以上。虽然过去几十年间喉癌的治疗方式获得了长足的进展,但患者五年生存率却没有获得提高,研究显示1975年~2010年间美国喉癌患者的5年生存率从66%降到63%[1-2]。喉癌治疗以外科手术为主,放化疗是不可或缺的组成部分,近年来新型分子治疗成为一种潜在选择[1]。大量研究表明,诸多因子参与到的喉癌的肿瘤学进程中,探明喉癌的分子机制对喉癌的早期诊断及研究新的分子靶标至为重要[3-6]。微小RNA(miR)在肿瘤的发生发展中扮演着重要的角色,它们既可以作为促癌基因也可以作为抑癌基因[7-8]。近年来,miR在喉癌中的研究越来越多, 各种研究表明在喉癌的早期诊断、治疗和预后等方面具有潜在作用[5]。目前miR-936偶见于其他肿瘤的报道中,但是尚未见miR-936在喉癌中的研究报道,本文主要讨论miR-936对喉癌细胞株Hep-2的影响。
材料与方法
一、细胞培养
Hep-2细胞、HEK293T细胞培养在含10%胎牛血清和青霉素(100 U/ml)、链霉素(100 ng/ml)的DMEM培养液中,置于37℃、5%CO2的培养箱中培养,以0.25%胰蛋白酶消化细胞传代,每 1~2 d传代1次,取处于对数生长期的细胞用于后续实验。
二、质粒构建及病毒包装
通过将合成好的pre-miR-936(F:CCGGTCAAGGCCACTGGGACAGTAGAGGGAGGAATCGCA-GAAATCACTCCAGGAGCAACTGAGAGACCTTCTAC-TTTACCAGGTCCTGCTGGCCCAGATTTTTG; R: AA-TTCAAAAATCTGGGCCAGCAGGACCTGGTAAAGTA-GAAGCAAGGTCTCTCAGTTGCTCCTGGAGTGATTT-CTGCGATTCCTCCCTCTACTGTCCCAGTGGCCTTGA)克隆到慢病毒载体pLKO.1-GFP质粒中,构建了miR-936慢病毒载体。将pLKO.1-GFP-miR-936与psPAX和pMD2G共转染HEK293T,分别在转染后24 h、48 h、72 h、96 h收集培养液上清。收集浓缩的培养液上清,用浓缩的慢病毒(Lenti-miR-936)溶液感染Hep-2细胞,之后用嘌呤霉素筛选建立Hep-2的miR-936稳定表达细胞株及对照组细胞株(pLKO.1-GFP空载包装病毒并感染Hep-2细胞),分别称为miR-936组及Vector组。
三、实时荧光定量PCR(RT-qPCR)
采用HiPure Total RNA Mini Kit(Magen)从Hep-2细胞中提取总RNA。采用HiFi-script cDNA kit(Cwbio)进行逆转录(miR-936逆转录引物:GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACT-GGATACGACCTGCGA)。以上所有操作遵照产品说明书。SYBR 荧光染料法RT-qPCR检测Hep-2-miR-936稳定株中miR-936的表达量,U6作为内参RT-qPCR引物:miR-936(F): CACGCAACAGTAGAGGGA,miR-936(R):CCAGTGCAGGGTCCGAGGTA;U6(F):GCGCGTCGTGAAGCGTTC,U6(R):GTGCAGGGTCCGAGGT。反应条件:95 ℃ 10 s; 95 ℃ 30 s, 60 ℃ 20 s(共40个循环),所有反应均设3个复孔,记录系统返回的CT值,采用获得相对定量结果。引物由Sangon Biotech订购。
四、MTT实验
将Hep-2细胞以0.5×104~1×104/孔的密度接种到96孔板中,设3个复孔,在增殖实验中分别在种板4 h、24 h、48 h、72 h、96 h后加入MTT(每孔终浓度0.5 mg/ml),培养4 h,吸去培养液,加入100 μl DMSO。在λ= 570 nm处测定吸光度。
五、划痕实验
将Hep-2细胞(5×105/孔)接种到六孔培养皿中,直到细胞80%~90%汇合,吸尽培养液,先在皿底用黑线标记,用10 μl移液枪吸头在细胞单层上垂直于标记线划痕,并用磷酸盐缓冲液(PBS)洗涤两次。使细胞在无血清培养基中迁移24 h和48 h,分别观察并拍摄划痕,选取3张图片进行统计分析,迁移的距离从显微照片中测量。
六、Transwell小室实验
将Hep-2细胞接种在改良的Boyden室(Corning)的上部隔室中,用无血清培养基培养。下室含有10%胎牛血清的培养基,在37℃、5%CO2中培养24 h。擦去膜上表面的细胞,并将侵入底部的细胞用10%结晶紫染色,然后随机选取3个不同的视野拍摄并计数。
七、鬼笔环肽(Phalloidin)染色实验
将细胞接种在玻璃盖板上24 h,然后用4%多聚甲醛固定20 min,并用0.1%Triton X-100在室温下透化15 min。将盖玻片在黑暗中用100 nM罗丹明-鬼笔环肽在室温下孵育30 min。 之后用100 nM DAPI核复染。 将盖玻片在PBS中冲洗并倒置晾干。然后将这些载玻片用中性香脂密封,并在共聚焦显微镜下观察。
八、药敏实验
将Hep-2细胞以0.5×104~1×104个细胞/孔的密度接种到96孔板中,设3个复孔,于37℃、5% CO2培养箱中孵育至其贴壁(一般4 h),加入所属浓度梯度的药,继续孵育72 h,在光学显微镜下观察其细胞状态。加入10 μl MTT,而后继续孵育4 h,吸尽培养基,加入50 μl DMSO,用摇床缓慢摇10 min,待结晶完全溶解,用酶标仪在570 nm处检测其吸光值(OD570)。
九、统计学处理
结 果
一、miR-936过表达Hep-2细胞株构建
由于构建的pLKO.1-GFP-miR-936质粒以及空载pLKO.1-GFP包含荧光基因GFP,成功感染的Hep-2细胞在蓝光激发下能够产生绿色荧光,未被感染的细胞则不能产生绿色荧光,因此比较同一光学显微镜视野下白光图片及荧光图片中细胞数量能够判断慢病毒感染效率,miR-936组感染效率为92%(细胞数量比值,荧光/白光=720/780),Vector组感染效率为0.91(细胞数量比值,荧光/白光=510/560)(图1A)。RT-qPCR结果显示,miR-936组细胞的miR-936 mRNA表达水平高于Vector组(2.36±0.47vs1.00±0.10,t=5.600,P﹤0.05),提示miR-936过表达的Hep-2稳转株构建成功(图1B)。
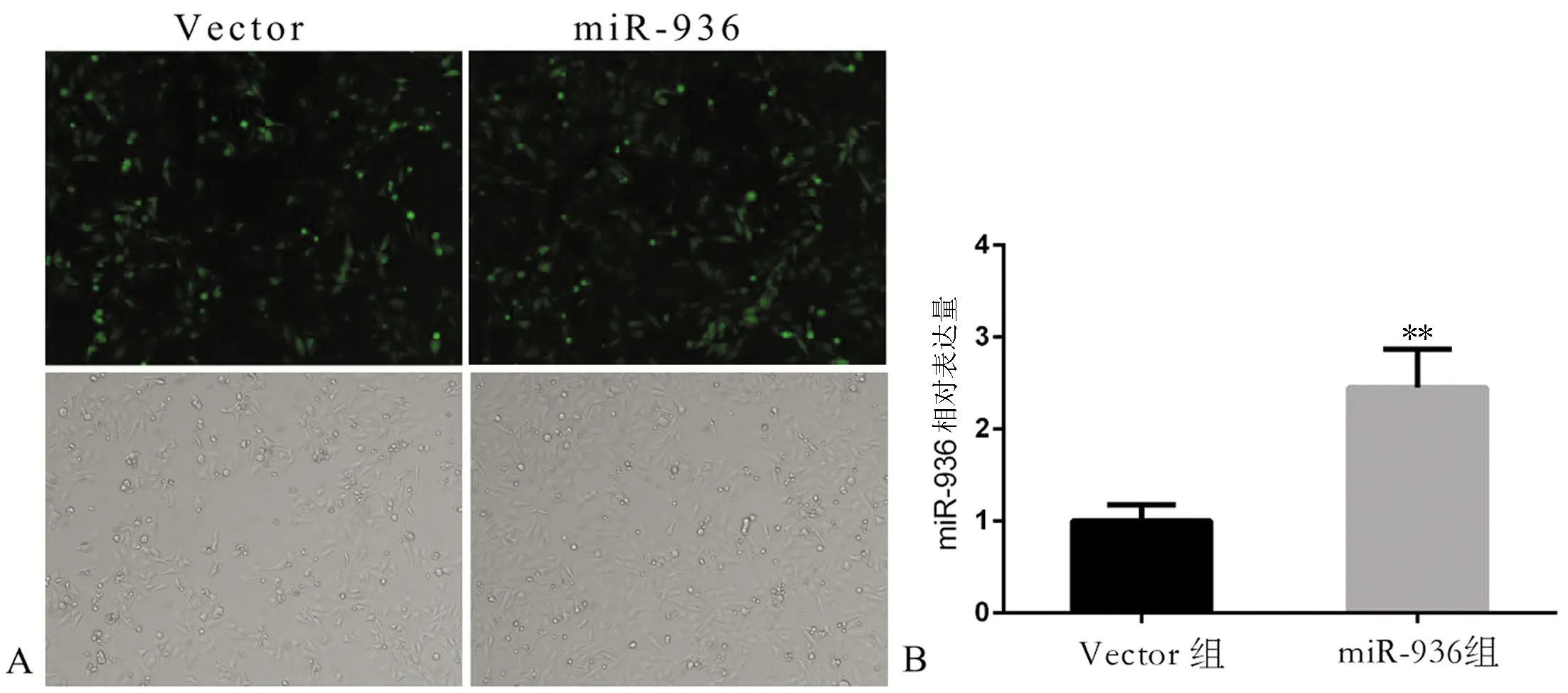
图1 miR-936过表达Hep-2细胞株和对照组构建(×100)
A:miR-936组与Vector组荧光表达情况;B: miR-936组与Vector组RT-qPCR检测miR-936的表达;与Vector组比较,**P<0.01
二、过表达miR-936抑制Hep-2细胞增殖能力
将miR-936组和Vector组细胞等量接种于96孔板,2组细胞在每个时间点分别设置3个复孔,0、4 h的MTT结果显示2组接种量几乎一致(t值分别为2.451和0.324,P均>0.05),而在48、72、96 h,2组出现差异,miR-936组细胞增殖速度低于Vector组(图2),差异有统计学意义(t值分别为4.939、6.154、3.739,P均<0.05)。
三、miR-936抑制Hep-2细胞迁移能力
划痕后细胞培养至24、48 h,miR-936组细胞的相对划痕宽度均宽于Vector组(t值分别为8.071、6.318,P均﹤0.01),提示miR-936过表达能够减弱Hep-2细胞的迁移能力,见图3。
四、miR-936抑制hep-2细胞侵袭能力
Transwell小室实验结果(图4)显示, miR-936组细胞的穿膜数量少于Vector组(t=8.342,P<0.01),提示miR-936过表达能够减弱Hep-2细胞的侵袭能力。
五、miR-936抑制Hep-2细胞表面微管微丝表达
鬼笔环肽染色显示miR-936组Hep-2细胞表面的微管微丝明显少于Vector组(图5),miR-936组细胞表面微管微丝的减少提示其侵袭迁移能力的减弱。

图2 miR-936对Hep-2细胞增殖的影响
*P<0.05,**P<0.01
六、miR-936增加Hep-2细胞对顺铂和阿霉素的敏感性
与Vector组相比,miR-936组Hep-2细胞对顺铂和阿霉素的敏感性增加(图6),DDP组0~100 μm的t值分别为0.000,-0.456,3.389,4.057,6.925,0.826,-2.782,2组在1、3、10 μm处比较差异有统计学意义(P均<0.05);DOX组0~10 μm的t值分别为0.000,5.181,5.598,3.112,1.610,0.437,0.831,2组在0.03、0.1、0.3 μm处比较差异有统计学意义(P均<0.05 )。
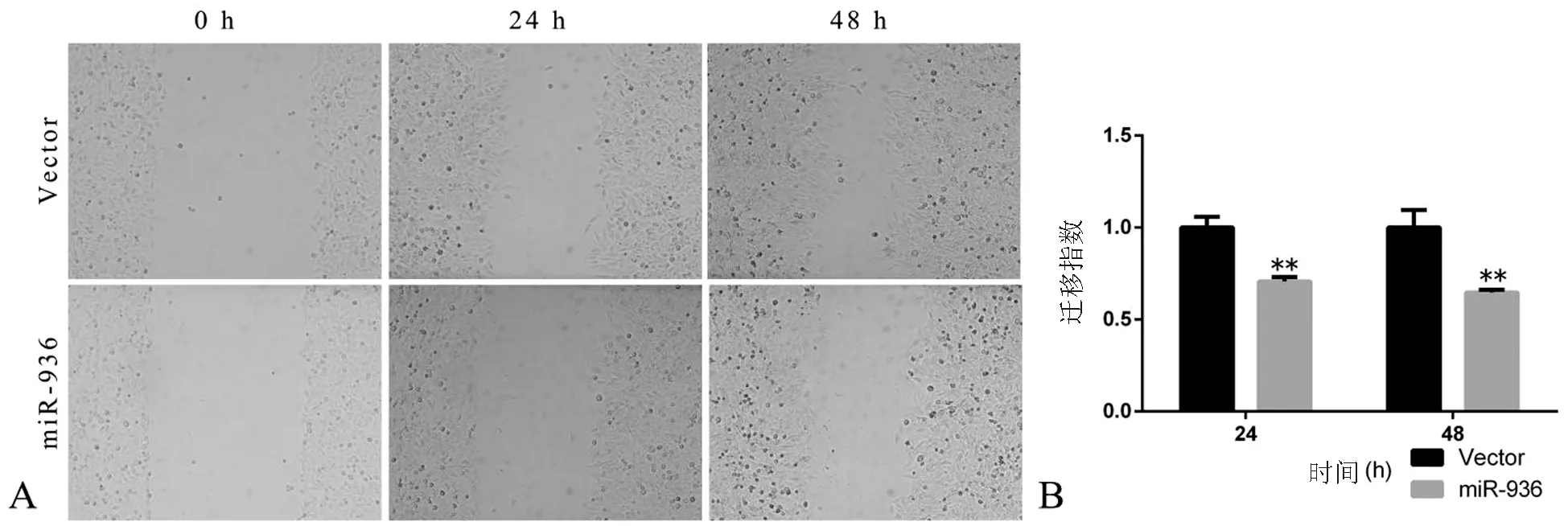
图3 miR-936对Hep-2细胞迁移的影响(×100)
A:2组细胞培养至0、24、48 h的相对划痕宽度;B:2组细胞相对划痕宽度比较,**P<0.01
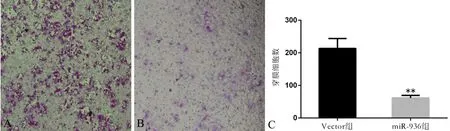
图4 miR-936对Hep-2细胞迁移的影响(×100)
A:Vector组;B: miR-936组;C:2组细胞的穿膜数量比较,**P<0.01
讨 论
miR是一类具有控制基因转录后表达水平的能力的调控分子。miR通过降解mRNA或阻止mRNA翻译来减少蛋白质的产生。因此,miR可以通过抑制其他促癌基因或者抑癌基因的基因转录后表达参与到肿瘤的分子机制网络中。有系统性研究指出,在喉癌中有潜在诊断价值的miR有miR-196a,miR-21,miR-27a等,对预后评价有潜在价值的miR有miR-101、miR-126、miR-152、miR-19a等,而一些miR如miR-1、miR-129-5p、miR-155、miR-206等有可能是潜在的治疗靶点[5]。
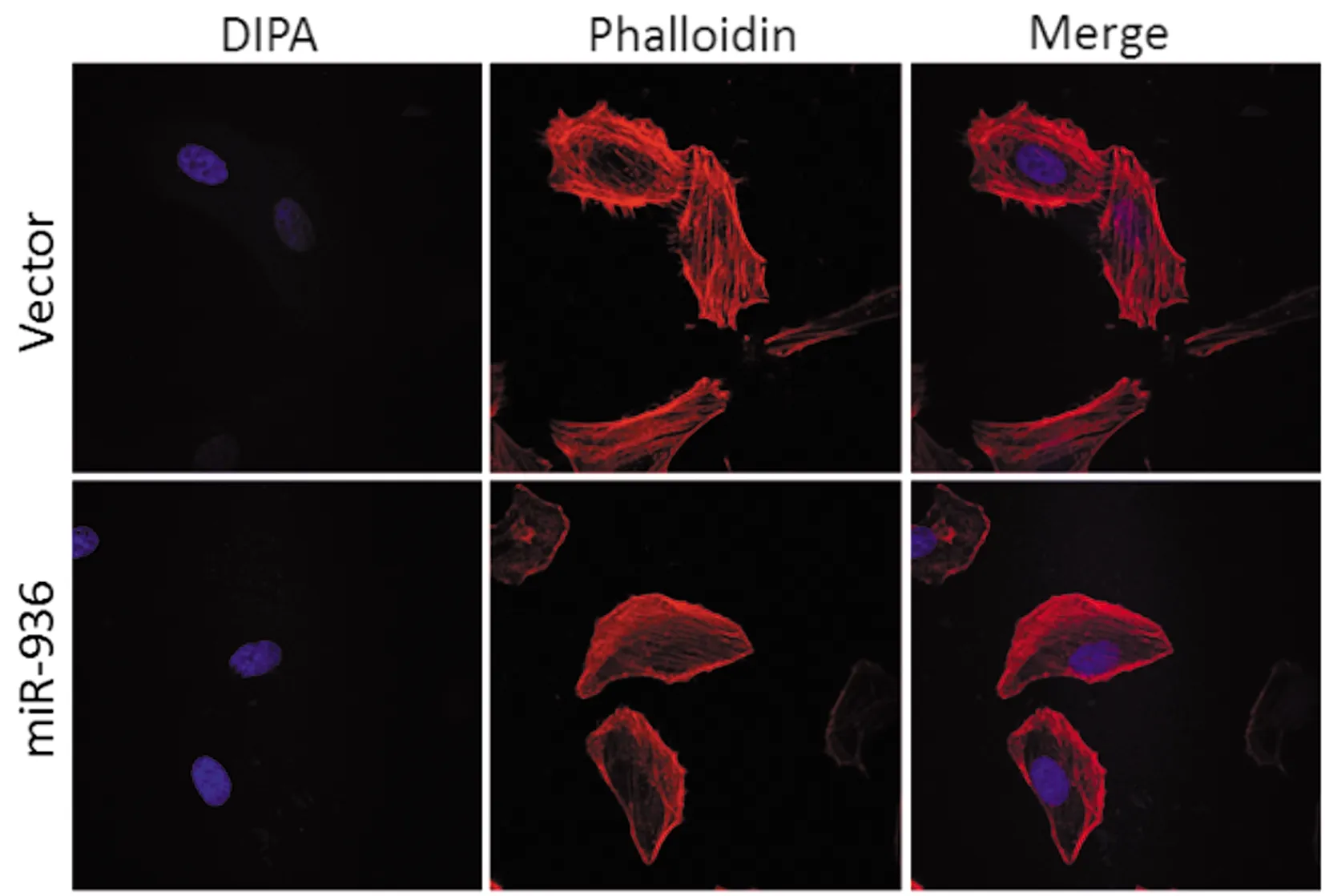
图5 miR-936对Hep-2细胞微管微丝的影响(×600)
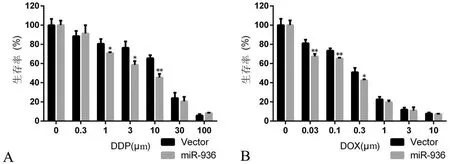
图6 miR-936增加Hep-2细胞对顺铂和阿霉素的敏感性
A:对顺铂的敏感性;B: 对阿霉素的敏感性;*P<0.05,**P<0.01
研究发现相比于正常组织,miR-936在 子宫内膜癌(EEC)和舌鳞状细胞癌中高表达[9-10]。在结直肠癌转移组织中也有报道miR-936呈现高表达[11-12]。另有研究发现与正常人相比,miR-936在肝细胞癌病人血液细胞微泡(Microvesicles)中高表达[13-14]。而另一项研究显示miR-936在人非小细胞肺癌细胞系(EPLC-32M1、A549和801D)和人胰腺导管腺癌细胞系(MiaPaCa-2、PANC-1和Hs766T)中呈现低表达[15-17]。由此可见,miR-936在不同肿瘤及肿瘤细胞株中可能扮演着不同角色,而其在喉癌中扮演的角色及在喉癌细胞生物学行为中的具体作用尚不可知。
本研究显示,过表达的miR-936组Hep-2细胞对顺铂和阿霉素的敏感性明显增加,具体机制尚不清楚。而Zhang等[18]研究发现抗肿瘤药物斑蝥素(Cantharidin)可以上调乳腺癌细胞MCF7中miR-936的表达。另有研究发现一种对肾细胞癌细胞有抑制作用的强心苷类药物(Amantadig)能够升高肾细胞癌细胞中miR-936的表达[19]。这些研究提示miR-936也可能作为抗肿瘤药物的下游基因发挥抗肿瘤作用。因此miR-936有可能既直接增加肿瘤细胞对于化疗药物的敏感性,同时也作为抗肿瘤药物的下游基因发挥抗肿瘤作用。
为了进一步研究miR-936对喉癌细胞的作用,我们分别进行了细胞增殖实验、划痕实验、Transwell侵袭实验以及鬼笔环肽染色实验。细胞增殖实验发现过表达的miR-936明显抑制了Hep-2细胞的增殖能力(与Vector组相比较)。划痕实验和Transwell侵袭实验发现过表达的miR-936能够明显抑制Hep-2细胞的迁移和侵袭能力,并且在鬼笔环肽染色实验中发现miR-936组细胞表面微管微丝明显减少。这些结果提示miR-936能够抑制喉癌细胞的侵袭和迁移能力。Zhang等[20]研究发现,相对于N0期(无淋巴结转移)胃癌组织,miR-936在N3期(超过7个淋巴结转移)的胃癌组织中明显低表达,提示miR-936低表达可能和胃癌转移有关。这与本研究发现的miR-936能够抑制喉癌细胞的侵袭和迁移能力的结果相一致。
综上所述,miR-936在不同肿瘤中可能有着不同的作用,本研究发现并初步验证了miR-936对于喉癌细胞系Hep-2的抑制作用,并从多个方面验证了miR-936作为抑癌基因抑制喉癌细胞系Hep-2的增殖、迁移和侵袭能力,并且增加其药物敏感性,提示miR-936在喉癌治疗中可能有潜在的应用价值。
[1] Steuer CE, El-Deiry M, Parks JR, Higgins KA, Saba NF. An update on larynx cancer. CA Cancer J Clin,2017,67(1):31-50.
[2] Siegel RL, Miller KD, Jemal A. Cancer statistics, 2015. CA Cancer J Clin,2015,65(1):5-29.
[3] de Miguel-Luken MJ, Chaves-Conde M, Carnero A. A genetic view of laryngeal cancer heterogeneity. Cell Cycle,2016,15(9):1202-1212.
[4] Rafferty MA, Fenton JE, Jones AS. An overview of the role and inter-relationship of epidermal growth factor receptor, cyclin D and retinoblastoma protein on the carcinogenesis of squamous cell carcinoma of the larynx. Clin Otolaryngol Allied Sci,2001,26(4):317-320.
[5] Li P, Liu H, Wang Z,He F, Wang H, Shi Z, Yang A, Ye J. MicroRNAs in laryngeal cancer: implications for diagnosis, prognosis and therapy. Am J Transl Res,2016,8(5):1935-1944.
[6] Nadal A, Cardesa A. Molecular biology of laryngeal squamous cell carcinoma. Virchows Arch,2003,442(1):1-7.
[7] Ventura A, Jacks T. MicroRNAs and cancer: short RNAs go a long way. Cell,2009,136(4):586-591.
[8] 葛曙雄,王涌. 微小RNA与肿瘤微环境. 新医学,2016,47(1):7-11.
[9] Snowdon J, Zhang X, Childs T, Tron VA, Feilotter H. The MicroRNA-200 family is upregulated in endometrial carcinoma. PLoS One,2011,6(8):e22828.
[10] Rentoft M, Fahlén J, Coates PJ, Laurell G, Sjöström B, Rydén P, Nylander K. miRNA analysis of formalin-fixed squamous cell carcinomas of the tongue is affected by age of the samples. Int J Oncol,2011,38(1):61-69.
[11] Chang J, Huang L, Cao Q, Liu F. Identification of colorectal cancer-restricted microRNAs and their target genes based on high-throughput sequencing data. Onco Targets Ther,2016,9:1787-1794.
[12] Kou Y, Qiao L, Wang Q. Identification of core miRNA based on small RNA-seq and RNA-seq for colorectal cancer by bioinformatics. Tumour Biol,2015,36(4):2249-2255.
[13] Lu L, Guo D, Chen X, Xiong W,Jie S,Li H. Abnormal miRNAs targeting chromosome open reading frame genes were enriched in microvesicles derived from the circulation of HCC. Biochem Genet,2016,54(2):120-133.
[14] Li H, Sun L, Chen X,Xiong W, Hu D, Jie S. Microvesicle microRNA profiles and functional roles between chronic hepatitis B and hepatocellular carcinoma. Clin Transl Oncol,2014,16(3):315-321.
[15] Liu M, Zhou K, Cao Y. MicroRNA-944 affects cell growth by targeting EPHA7 in non-small cell lung cancer. Int J Mol Sci,2016,17(10).pii: E1493.
[16] Lee E M, Shin S, Cha H J, Yoon Y, Bae S, Jung JH, Lee SM, Lee SJ, Park IC, Jin YW, An S. Suberoylanilide hydroxamic acid (SAHA) changes microRNA expression profiles in A549 human non-small cell lung cancer cells. Int J Mol Med,2009,24(1):45-50.
[17] Toste PA, Li L, Kadera BE, Nguyen AH, Tran LM, Wu N, Madnick DL, Patel SG, Dawson DW,Donahue TR. p85alpha is a microRNA target and affects chemosensitivity in pancreatic cancer. J Surg Res,2015,196(2):285-293.
[18] Zhang H, Yan X. Cantharidin modulates the E2F1/MCM7-miR-106b-93/p21-PTEN signaling axis in MCF-7 breast cancer cells. Oncol Lett,2015,10(5):2849-2855.
[19] Nolte E, Wach S, Silva IT, Lukat S, Ekici AB, Munkert J, Müller-Uri F,Kreis W, Oliveira Simões CM, Vera J, Wullich B, Taubert H, Lai X. A new semisynthetic cardenolide analog 3β-[2-(1-amantadine)-1-on-ethylamine]-digitoxigenin (AMA-NTADIG) affects G2/M cell cycle arrest and miRNA expression profiles and enhances proapoptotic survivin-2B expression in renal cell carcinoma cell lines. Oncotarget,2017,8(7):11676-11691.
[20] Zhang J, Zhu Z, Sheng J,Yu Z, Yao B, Huang K, Zhou L, Qiu Z, Huang C. miR-509-3-5P inhibits the invasion and lymphatic metastasis by targeting PODXL and serves as a novel prognostic indicator for gastric cancer. Oncotarget,2017,8(21):34867-34883.
