启动子的选择及优化在酿酒酵母代谢工程中的应用
2018-06-29王珂雯尹雪王宇李玉花
王珂雯 尹雪 王宇 李玉花
(东北林业大学生命科学学院,哈尔滨 150040)
天然产物作为生物体内自身代谢合成的一种微量次生代谢物,因其具有高效的药理活性,被人们广泛应用于医疗保健和食品等领域。近年来利用微生物进行基因工程操作,实现天然产物的大量合成已成为目前的研究热点,如利用大肠杆菌(Esherichia coli)或酵母合成可以预防心脑血管疾病的白藜芦醇、具有止痛功效的吗啡以及具备抗癌功效的人参皂苷和紫杉醇等药物[1-3];在链霉菌(Streptomyces)中实现了深海极端环境微生物中具有化学防御、信号传导的特殊代谢产物的异源合成[4]。酿酒酵母作为已完成全基因组测序的真核模式生物,与其他真核生物如烟草细胞相比,它在代谢工程研究中具有生物繁殖速度快、生长周期短、易于培养、不易污染等特点[5];而与原核生物如大肠杆菌相比,不仅可以高密度发酵,还具有其他优点:含有线粒体、内质网等细胞器,利于表达有生物活性的异源蛋白,特别是适合植物细胞色素P450类蛋白的表达[6];DNA重组效率高,能够一次性插入多个大片段的外源基因,并且能够稳定遗传。同时,最近酿酒酵母人工染色体从头合成的成功实现为未来发挥酵母巨大潜力提供可能[7]。目前研究人员利用酿酒酵母表达系统已经成功合成了生物碱、萜类、不饱和脂肪酸及其他一些重要的化合物(表1)。

表1 利用酿酒酵母生产的重要化合物
目前对代谢工程的改造主要集中在通过导入特定的催化酶、调节蛋白来构建新的代谢通路,或者优化酿酒酵母自身的代谢途径,而选择合适的基因启动子可以对能量和营养物质在终产物代谢途径和细胞自身代谢途径之间的供给交换进行精细和定量地调控,实现目标基因表达强度的最佳组合。因此,在酿酒酵母工程菌中选择合适的启动子显得尤为重要。启动子作为转录调控的重要元件,其活性的强弱可以显著影响异源基因转录的起始、持续时间和表达的程度,进而影响外源基因在宿主中的转录效率,同时也决定了细胞中mRNA的总体丰度和蛋白质含量,最终高效地合成目标产物。本文将基于如何选择和修饰酿酒酵母启动子用来高效合成目标产物,分别对酿酒酵母启动子结构、类型和优化策略几个部分进行了详细介绍。
1 酿酒酵母启动子结构
启动子位于结构基因5′端的上游区域,直接调控基因时空特异性表达。真核生物启动子由核心启动子(Core promoter)和上游启动子元件(Upstream promoter element)组成(图 1)。
1.1 核心启动子
核心启动子由TATA盒(TATA box)、TFⅡB识别元件(Transcription factor for RNA polymerase IIB recognition element,BRE)、 起 始 子(Initiator,Inr)和下游启动子元件(Downstream promoter element,DPE)4个元件构成,它们位于转录起始位点的上游40 bp到下游40 bp之间(图1)。但在酿酒酵母中,大部分元件都已退化[21],仅有20%的酿酒酵母基因启动子还存在着TATA盒,共同序列为TATA(A/T)A(A/T)(A/G)[22]。并且酿酒酵母启动子的 TATA盒位置具有可变性,一般位于转录起始位点上游的40-120 bp 处[23]。
1.2 上游启动子元件
酿酒酵母核心启动子TATA盒上游存在一段保守的调控区被称为上游启动子原件,可分为上游激活序列(Upstream activation sequences,UAS)和上游抑制序列(Upstream repression sequences,URS)[24]。转录激活因子和转录抑制因子可以分别与其特异性结合,从而促进或抑制RNA聚合酶的结合效率调节下游基因的转录。上游激活序列最先在酵母中被发现,如半乳糖代谢酶基因Gal的上游激活序列由4个相连的17 bp序列组成;精氨酸酶基因Car1的上游激活序列为一段13 bp的序列[25]。上游激活序列的作用类似于增强子,但与增强子不同的是,上游激活序列一般位于转录起始位点上游的100-500 bp之间,距离过远就会失去促进转录的活性[26]。
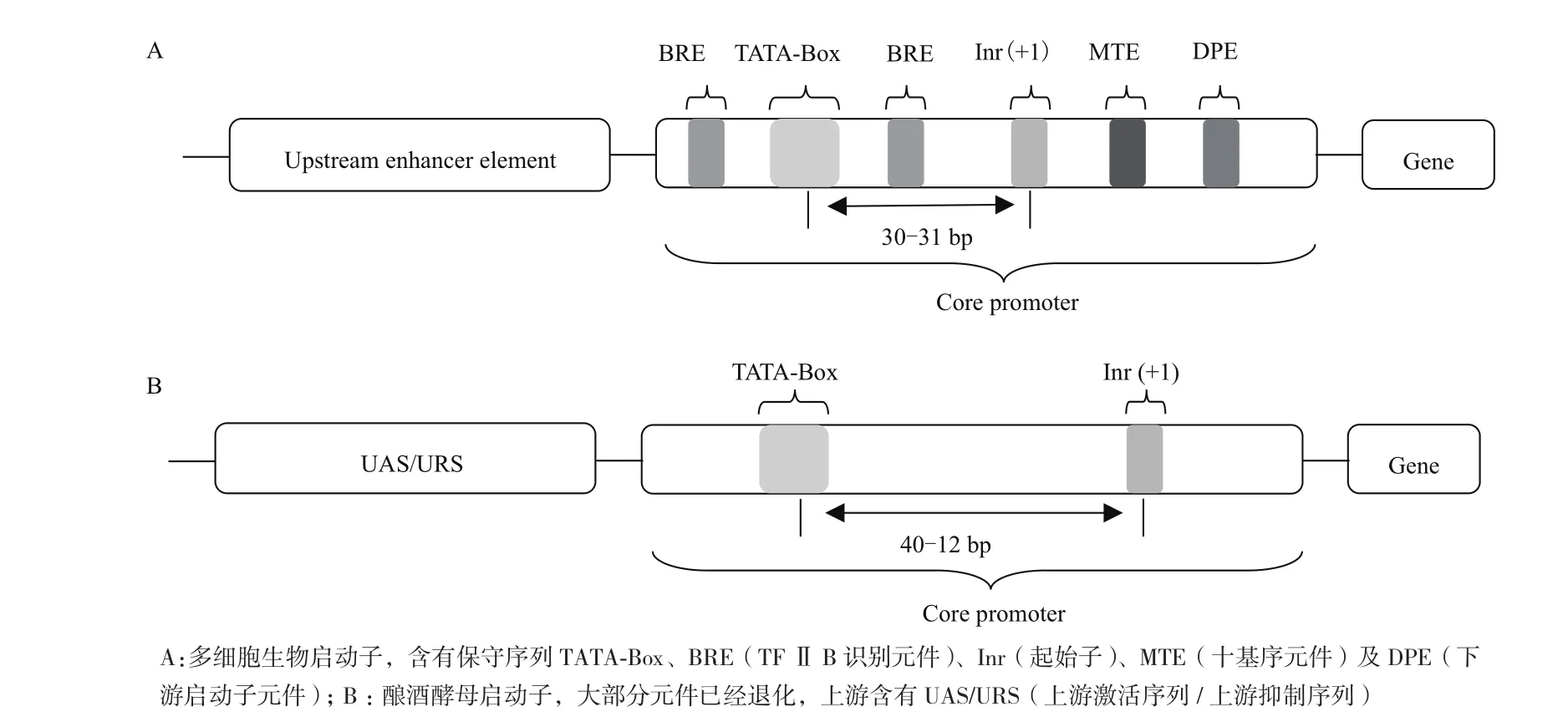
图1 启动子结构示意图
2 酿酒酵母启动子类型
在酿酒酵母工程菌的实际应用中,常用的天然启动子来源于酿酒酵母宿主本身和外源宿主。酿酒酵母宿主内源启动子根据其作用方式或功能可分为组成型启动子、诱导型启动子和双向启动子,而来源于外源宿主的启动子被称为异源启动子。选择合适类型的启动子不仅可以控制目标基因的表达,还可以对基因的表达量进行精细调节。
2.1 组成型启动子
组成型启动子又称非特异性启动子,在其调控下,同一基因在不同生长环境及生长阶段的表达无明显差异,且始终维持在一个相对稳定的水平。
基因表达量与组成型启动子的强弱密切相关,因此组成型启动子相对强度的大小在实际应用中尤为重要。当需要提高基因的表达量时,选择强启动子,反之则选弱启动子(表2),目前组成型启动子的相对强度可以采用多种报告系统来进行评估。Partow等[27-28]通过检测LacZ的活性,比较了7种组成型启动子的强度,获得的相对强度大小为:pTEF1≈pPGK1≈pTDH3> pTPI1≈pPYK1 > pADH1 >pHXT7。Sun等[29]从酿酒酵母中克隆了14种组成型启动子并通过GFP的发光强度和mRNA水平来检测启动子的相对强度,其中pTEF1强度相对最高,pPGI1的强度相对最低。Blount等[31]利用流式细胞仪检测了GFP的发光强度,从而确定pPFY1、pADH1、pCYC1、pBIO2、pCHO1中 pADH1的 强 度相对最高,pCHO1的强度相对最低。Monfort等[31-32]通过表达脂肪酶来比较酿酒酵母中的pACT1、pADH1、pPGK1及pTDH1的相对强度,最终发现pACT1的强度相对最高。
组成型启动子虽然能够持续高强度表达目的蛋白,但基因过量表达会造成代谢紊乱,同时会引起重组蛋白在生长过程中降解[33],并且组成型启动子不适用于毒蛋白的表达。
2.2 诱导型启动子
诱导型启动子在某些特定的物理及化学信号的诱导刺激下,可以大幅度地提高或降低基因的转录水平,因此可以通过人为添加诱导剂定时定量地对目标基因表达程度进行调控。诱导型启动子又可以按照功能分为激活型启动子和阻遏型启动子(表3)。
2.2.1 激活型启动子 激活型启动子即正调控启动子,一般是通过招募转录激活因子或释放转录抑制因子来实现转录调节的。其优点是只有当诱导物存在时,基因才可以正常表达,没有诱导物存在时基因表达水平很低甚至没有。
激活型启动子常用的诱导剂有碳源(半乳糖、葡萄糖)、铜离子、生物素及α1调节蛋白。目前在酿酒酵母中应用最多的激活型启动子是来源于降解半乳糖代谢途径的半乳糖诱导启动子(pGAL1、pGAL7、pGAL10)。当培养基中的碳源为半乳糖时,转录诱导因子Gal3p与半乳糖结合后,会与抑制因子Gal80p相互作用,转录激活因子Gal4p发挥活性,从而激活目标基因的表达[34-35]。目前利用该启动子系统已经成功在酿酒酵母中合成了青蒿酸[36]、人参皂苷[37]等物质。
2.2.2 阻遏型启动子 酵母启动子阻遏系统建立在阻遏蛋白与转录因子在空间构型相互作用的基础之上。当诱导物存在时,其会与激活蛋白结合,阻遏蛋白则与启动子上的元件结合,抑制基因的转录。
阻遏型启动子常用的抑制剂有碳源(葡萄糖)、磷酸盐、铁离子、蛋氨酸、渗透压及氧气。酿酒酵母adh2基因编码乙醇脱氢酶II,其转录起始于葡萄糖耗尽,乙醇作为碳源发酵的同时,转录激活因子乙醇脱氢酶调节蛋白Adr1可大量表达并起始ADH2的转录[38]。目前已经有研究团队利用该启动子表达外源蛋白,在酿酒酵母工程菌中合成了聚酮化合物[39]、三醋酸内酯[40]等物质。
应用诱导系统可以将细胞的生长阶段与目标蛋白的生产阶段分开,避免了外源蛋白早期表达对自身细胞的伤害。但该类型启动子存在着操作繁琐,诱导剂价格昂贵,添加的诱导剂可能会对细胞本身的生长产生负面影响等诸多在实际生产中难以解决的问题。

表2 酿酒酵母中常用的组成型启动子
2.3 双向启动子
双向启动子普遍存在于酿酒酵母的基因组中,它能以两个相反的方向启动转录。目前酿酒酵母中应用最多的双向启动子载体是pESC载体,通过pGAL1-pGAL10可以在同一时间转录两个不同的基因,这两个启动子的转录起始位点相距约600 bp。该系列的载体已用于人参皂苷CK的合成[37]。
2.4 异源启动子
在实际应用中,使用酵母内源启动子可能会干扰酵母的自身代谢反应,降低酵母的生长活性,因此可以采用外源启动子来构建代谢途径。其中常用的异源启动子可以来源于病毒、细菌和真菌等外源宿主。目前,在酿酒酵母中常用巨细胞病毒的启动子pCMV组成型表达外源基因,它的活性受亚砷酸盐的调控[52]。细菌中的tet操纵子,会被培养基中的四环素类似物多西霉素所抑制,而多西霉素并不会影响酵母的生长状态和内源基因的mRNA转录水平[53]。应用棉病囊霉(Ashbya gossypii)的 pTEF1进行特定位点的同源重组时,酵母内源的pTEF1并不会影响外源基因的重组效率,同时又避免了在同源重组过程中因序列相似度高而产生错误插入[54]。

表3 酿酒酵母中常用的诱导型启动子
3 酿酒酵母启动子优化
根据上文所述,在代谢工程中应用自身和异源启动子的方法已经获得了广泛的应用,但随着功能基因组学和代谢组学的快速发展,科学家们通过对现有的启动子进行突变、融合的优化方式来获得不同功能、不同强度的启动子,以期在进一步提高转录能力的同时还可以对宿主代谢进行更为精细的人工调控。以下将介绍酿酒酵母启动子工程的具体策略及相关的应用实例。
3.1 天然启动子突变优化
在明确天然启动子活性的情况下,可以通过突变转录因子结合位点来改变转录因子的结合能力,直接达到改变其启动强度的目的。根据作用方式的不同,可以将突变优化分为随机突变优化(易错PCR、饱和诱变)和定向突变优化(直接修饰)。
易错PCR(Error-prone PCR)适用于整个启动子区域,通过随机突变可产生具有不同的强度和功能的启动子。Redden和Nevoigt等[55-56]使用易错PCR将pTEF1随机突变并筛选出活性范围是天然pTEF1活性 8%-120%的11个突变体,最终发现两个活性中等的启动子可以降低副产物甘油的产量,该研究使酒精、甘油产量及细胞生长三者之间达到一个相对平衡的状态。此外,也有人将pDAN1随机突变并得到两个对氧气含量不敏感的突变体,使下游基因的表达量显著增加,大大提升了终产物的合成量[57]。2015年,张旭等[58]利用易错PCR 筛选得到的具有高表达性能的pTEFV1,使GFP和人血清白蛋白的表达能力较改造前有明显增加。
饱和诱变(Saturation mutagenesis)是指对基因的某一特定局部小区域内的碱基进行所有类型的突变。该技术最早应用于细菌启动子系统,有效地改变了单基因或多基因操纵子的表达水平[59]。Jeppsson团队参考天然启动子相关结构设计了一个名为YRP的启动子[60-61],降低了葡萄糖-6-磷酸脱氢酶的活性,进而增加乙醇的产量。对pPFY1的非必需区域进行饱和诱变可筛选得到36个有不同活性的非天然启动子。近年来也有人对pSUL1中的493 bp序列进行诱变及筛选,发现突变后的结构可以提高转录效率[62]。
事实上,对启动子强度的调控是基于核苷酸序列响应转录机制的累积效应。直接对目标启动子的上游元件进行增加或删除修饰就可以得到活性不同的启动子。将pGAL1上游激活元件UASGAl串联加倍得到的突变启动子UASGal-pGAL1在半乳糖诱导下其活性略高于pGAL1[64]。对pADH1的5′端调控区域进行不同程度的删减可以显著改变启动子对葡萄糖的应答,从而改变基因的表达强度[64-67]。此外,通过直接优化转录因子也可以达到增强启动子活性的目的。酿酒酵母pARO9在添加外源色氨酸的情况下可以组成型表达,而蛋白质工程优化后的转录因子Aro80p可增强与该启动子的结合效率,从而增强该启动子的活性[68-69]。基于pGAL的作用机制,敲除抑制因子Gal80后的启动子在葡萄糖中具有组成型表达的活性[70]。
3.2 人工启动子杂合优化
杂合启动子就是将两种或两种以上不同来源的启动子元件融合构成的一个新的启动子。杂合启动子的高强度和稳定性对于精细调控细胞中的基因表达具有极其重要的意义。
酿酒酵母中的杂合启动子大多是将自身不同的启动子核心区域与调节区域融合进而增强杂合启动子的活性。将不同比例或组合型的转录激活元件分别与pTDH3融合后,发现串联3倍的UASCLB启动子可使转录活性提高2.5倍。因此,利用该方法可以将弱启动子定向转变为强启动子。此外,与天然启动子相比杂合启动子的活性更加稳定,将3个串联的激活元件UASCLB与pTEF1的核心区域所组装而成的人工杂合paTEF1在不同宿主的不同生长阶段其活性均高于天然pTEF1[71]。
酿酒酵母杂合启动子的构建也可以采用将不同来源的启动子核心区域与调节区域融合的方式。其中,在酿酒酵母中应用最多的异源启动子为细菌的四环素操纵子。1997年,Gari等[72]构建了一系列由tetO-CYC1杂合启动子驱动的表达载体,在缺乏抑制剂多西霉素的环境下,基因的表达活性可以提高1 000倍,该启动子可应用于P-蛋白和羊毛甾醇脱甲基酶的表达[73-74]。而后Belli团队构建了一个酿酒酵母四环素操纵子诱导基因表达的双系统,将反式激活因子中的tetR进行突变优化并命名为tetR’,通过外源施加的四环素可快速激活下游基因表达[75-76]。
另外,也有利用杂合转录因子调节目标基因表达的可调控系统。例如,Shimizu-Sato等[77]通过表达两种嵌合蛋白光敏色素的Gal4-DNA结合域和PIF3-Gal4激活域,进而实现光信号调控下的基因表达调控目标,构建了一个可精确定量的诱导表达系统,有利于工业生产的大规模应用。也有应用人类激素来调控启动子的活性,构建含有β-雌二醇受体、Gal4结合域,以及疱疹病毒蛋白的转录激活结构域的杂合转录激活因子,通过添加β-雌二醇可以显著提高下游基因的表达水平[78]。2016 年,Hector等[79]利用芽孢杆菌中的DNA结合阻遏因子(XylR)与pTEF形成的新型杂合启动子,使pTEF的表达强度在木糖培养条件下增加25倍。
酿酒酵母启动子的优化在代谢工程精细调节方面拥有巨大的潜能,为了突出比较上述4种启动子优化策略的特点,现将其优缺点总结如下(表4)。

表4 酿酒酵母启动子优化类型的优缺点
4 总结和展望
酿酒酵母是一种食品级、基因组结构最为简单的真核微生物,具备安全、遗传操作便捷、培养简单和繁殖迅速等优势,是生产重组蛋白较理想的表达系统之一。启动子是代谢工程中最基本的功能元件,是最有效的调节代谢途径的工具。通过改造酿酒酵母启动子从而改变其自身的代谢途径来高效地获取目标天然产物已成为一种极具前景的策略。本文梳理了当前利用酿酒酵母合成天然产物过程中的启动子结构和在酿酒酵母工程菌中使用的4类天然启动子,总结概括了每一类别的特点。此外,为了提高酿酒酵母启动子活性达到能够更加高效的合成目标天然产物的目的,归纳了酿酒酵母启动子突变和杂合的两种优化策略,以及该技术在实际生产中的一些应用。
近年来,随着合成生物学的发展和技术手段的更新,越来越多的研究人员在探索原核启动子模型中做出了巨大贡献,如通过在原核启动子上采用“绝缘化”和“非门”(NOT gate)功能基因网络预测的方法,表明该方法可以显著提高它们的模块化属性和组装过程的可预测性[84]。此外,Rohlhill等[85]采用荧光激活细胞分类术(FACS)和高通量筛选对大肠杆菌甲醛诱导启动子进行精确地定量分析,确定了单核苷酸分辨率,从而更高效的协调基因表达,并在合成生物学和代谢工程中建立了可预测的基因电路。基于当前的研究,越来越多的研究者也会对真核启动子进行深入探索,有望提高启动子控制基因的表达效率。
未来,通过启动子工程技术,酿酒酵母启动子将朝着以下几个方向进行改进:(1)研发基因表达量可控的组成型启动子;(2)研发培养条件温和、操作简单、诱导剂成本低廉的诱导型启动子;(3)具有高效转录效率的异源启动子并在发酵培养过程中得到有效控制;(4)研发能够更加高效、稳定、可控、成本低廉、可工业化的酿酒酵母启动子将是未来启动子工程技术突破的关键。这些研究成果的推广与应用,将有力地推进人工构建生命系统的研究进程,从而满足人们在健康、医疗、环境、食品等领域的高层次需求。
[1]Galanie S, Thodey K, Trenchard IJ, et al. Complete biosynthesis of opioids in yeast[J]. Science, 2015, 349(6252):1095.
[2]Li M, Kildegaard KR, Chen Y, et al. De novo production of resveratrol from glucose or ethanol by engineered Saccharomyces cerevisiae[J]. Metab Eng, 2015, 32:1-11.
[3]Zhou K, Qiao K, Edgar S, et al. Distributing a metabolic pathway among a microbial consortium enhances production of natural products[J]. Nature Biotechnology, 2015, 33(4):377-383.
[4]许静, 徐俊. 海洋共附生微生物天然产物生物合成基因研究进展[J]. 微生物学报 , 2008, 48(7):975-979.
[5]王勇. 新本草计划——基于合成生物学的药用植物活性代谢物研究[J]. 生物工程学报, 2017, 33(3):478-485.
[6]Jiang H, Wood KV, Morgan JA. Metabolic engineering of the phenylpropanoid pathway in Saccharomyces cerevisiae[J]. Appl Environ Microbiol, 2005, 71(6):2962-2969.
[7]Kannan K, Gibson DG. Yeast genome, by design[J]. Science,2017, 355(6329):1024.
[8]Galanie S, Thodey K, Trenchard IJ, et al. Complete biosynthesis of opioids in yeast[J]. Science, 2015, 349(6252):1095-1100.
[9]Dai Z, Liu Y, Zhang X, et al. Metabolic engineering of Saccharomyces cerevisiae for production of ginsenosides[J]. Metabolic Engineering, 2013, 20(5):146-156.
[10]Xie W, Liu M, et al. Construction of a controllable β-carotene bio-synthetic pathway by decentralized assembly strategy in Saccharomyces cerevisiae[J]. Biotechnol Bioeng, 2014, 1:125-133.
[11]Westfall PJ, Pitera DJ, Lenihan JR, et al. Production of amorphadiene in yeast, and its conversion to dihydroartemisinic acid, precursor to the antimalarial agent artemisinin[J]. Proc Natl Acad Sci USA, 2012, 109(3):111-118.
[12]Leonard E, et al. Investigation of two distinct flavone synthases for plant-specific flavone biosynthesis in Saccharomyces cerevisiae[J]. Appl Environ Microbiol, 2005, 71(12):8241.
[13]Medina VG, Almering MJH, Maris AJAV, et al. Elimination of glycerol production in anaerobic cultures of a Saccharomyces cerevisiae strain engineered to use acetic acid as an electron acceptor[J]. Appl Environ Microbiol, 2010, 76(1):190-195.
[14]Chen X, Nielsen KF, Borodina I, et al. Increased isobutanol production in Saccharomyces cerevisiae by overexpression of genes in valine metabolism[J]. Biotechnol Biofuels, 2011, 4(1):21.
[15]Yu KO, Ju J, Kim SW, et al. Synthesis of faees from glycerol in engineered Saccharomyces cerevisiae using endogenously produced ethanol by heterologous expression of an unspecific bacterial acyltransferase[J]. Biotechnol Bioeng, 2012, 1:110-115.
[16]Peraltayahya PP, Ouellet M, Chan R, et al. Identification and microbial production of a terpene-based advanced biofuel[J].Nature Communications, 2011, 2(1):483.
[17]Sauer M, Branduardi P, Valli M, et al. Production of l-ascorbic acid by metabolically engineered Saccharomyces cerevisiae and Zygosaccharomyces bailii[J]. Appl Environ Microbiol, 2004, 70(10):6086-6091.
[18]Raab AM, Gebhardt G, Bolotina N, et al. Metabolic engineering of Saccharomyces cerevisiae for the biotechnological production of succinic acid[J]. Metab Eng, 2010, 12(6):518-525.
[19]Maris AJAV, Geertman JMA, Vermeulen A, et al. Directed evolution of pyruvate decarboxylase-negative Saccharomyces cerevisiae, yielding a c2-independent, glucose-tolerant, and pyruvate-hyperproducing yeast[J]. Appl Environ Microbiol,2004, 70(1):159-166.
[20]Zhao L, Wang J, Zhou J, et al[modification of carbon flux in Sacchromyces cerevisiae to improve l-lactic acid production][J].Acta Microbiologica Sinica, 2011, 51(1):50.
[21]Blazeck J, Alper HS. Promoter engineering:Recent advances in controlling transcription at the most fundamental level[J].Biotechnol J, 2013, 8(1):46-58.
[22]Basehoar AD, Zanton SJ, Pugh BF. Identification and distinct regulation of yeast tata box-containing genes[J]. Cell, 2004, 116(5):699-709.
[23]Rando OJ, Winston F. Chromatin and transcription in yeast[J].Genetics, 2012, 190(2):351.
[24]Redden H, Alper HS. The development and characterization of synthetic minimal yeast promoters[J]. Nature Communications,2015, 6:7810.
[25]Giniger E, et al. Specific DNA binding of gal4, a positive regulatory protein of yeast[J]. Cell, 1985, 40(4):767-774.
[26]Hahn S, Young ET. Transcriptional regulation in Saccharomyces cerevisiae:Transcription factor regulation and function, mechanisms of initiation, and roles of activators and coactivators[J].Genetics, 2011, 189(3):705-736.
[27]Partow S, Siewers V, Bjørn S, et al. Characterization of different promoters for designing a new expression vector in Saccharomyces cerevisiae[J]. Yeast, 2010, 27(11):955-964.
[28]Wang D, Wang L, Hou L, et al. Metabolic engineering of Saccharomyces cerevisiae for accumulating pyruvic acid[J].Annals of Microbiology, 2015, 65(4):2323-2331.
[29]Sun J, Shao Z, Zhao H, et al. Cloning and characterization of a panel of constitutive promoters for applications in pathway engineering in Saccharomyces cerevisiae[J]. Biotechnol Bioeng, 2012, 8:2082.
[30]Blount BA, Weenink T, et al. Rational diversification of a promoter providing fine-tuned expression and orthogonal regulation for synthetic biology[J]. PLoS One, 2012, 7(3):e33279.
[31]Monfort A, Finger S, Sanz P, et al. Evaluation of different promoters for the efficient production of heterologous proteins in baker’s yeast[J]. Biotechnology Letters, 1999, 21(3):225-229.
[32]Hubmann G, et al. Natural and modified promoters for tailored metabolic engineering of the yeast Saccharomyces cerevisiae[J].Methods Mol Biol, 2014, 1152:17-42.
[33]Emmerstorfer A, et al. Over-expression of ice2 stabilizes cytochrome p450 reductase in Saccharomyces cerevisiae and pichia pastoris[J]. Biotechnol J, 2015, 10(4):623-635.
[34]Zacharioudakis I, et al. Bimodal expression of yeast gal genes is controlled by a long non-coding rna and a bifunctional galactokinase[J]. Biochem Biophys Res Commun, 2017, 1 :63-69.
[35]Horák J. Regulations of sugar transporters:Insights from yeast[J]. Current Genetics, 2013, 59(1-2):1-31.
[36]Ro DK, Paradise EM, Ouellet M, et al. Production of the antimalarial drug precursor artemisinic acid in engineered yeast[J].Nature, 2006, 440(7086):940.
[37]Yan X, Fan Y, Wei W, et al. Production of bioactive ginsenoside compound k in metabolically engineered yeast[J]. Cell Research, 2014, 24(6):770-773.
[38]Bahieldin A, et al. Efficient production of lycopene in Saccharomyces cerevisiae by expression of synthetic crt genes from a plasmid harboring the adh2 promoter[J]. Plasmid, 2014, 72 :18.
[39]Lee KK, Da SN, Kealey JT. Determination of the extent of phosphopantetheinylation of polyketide synthases expressed in escherichia coli and Saccharomyces cerevisiae[J]. Analytical Biochemistry, 2009, 394(1):75-80.
[40]Bernal DAN. Metabolic Engineering of Saccharomyces cerevisiae for the production of aromatic componds[D]. Brisbane: School of Chemistry and Molecular Biosciences, University of Queensland,2012.
[41]Kim SI, Ha BS, et al. Evaluation of copper-inducible fungal laccase promoter in foreign gene expression in pichia pastoris[J].Biotechnol Bioprocess Engineering, 2016, 1:53-59.
[42]Gross A, Rödel G, Ostermann K. Application of the yeast pheromone system for controlled cell-cell communication and signal amplification[J]. Lett Appl Microbiol, 2011, 5:521-526.
[43]Ammerer G. Identification, purification, and cloning of a polypeptide(prtf/grm)that binds to mating-specific promoter elements in yeast[J]. Genes Dev, 1990, 4(2):299-312.
[44]Gancedo JM, Flores CL, Gancedo C. The repressor rgt1 and the camp-dependent protein kinases control the expression of the suc2 gene in Saccharomyces cerevisiae[J]. BBA-General Subjects,2015, 1850(7):1362-1367.
[45]Silva NAD, Srikrishnan S. Introduction and expression of genes for metabolic engineering applications in Saccharomyces cerevisiae[J]. Fems Yeast Research, 2012, 12(2):197-214.
[46]Poor CB, Wegner SV, Li H, et al. Molecular mechanism and structure of the Saccharomyces cerevisiae iron regulator aft2[J].Proc Natl Acad Sci USA, 2014, 111(11):4043-4048.
[47]Zhu Y, Sun J, Zhu Y, et al. Endogenic oxidative stress response contributes to glutathione over-accumulation in mutant Saccharomyces cerevisiae y518[J]. Appl Microbiol Biotechnol,2015, 99(17):7069-7078.
[48]Noble J, Sanchez I, Blondin B. Identification of new Saccharomyces cerevisiae variants of the met2 and skp2 genes controlling the sulfur assimilation pathway and the production of undesirable sulfur compounds during alcoholic fermentation[J]. Microbial Cell Factories, 2015, 14(1):68.
[49]Anton C, Zanolari B, Arcones I, et al. Involvement of the exomer complex in the polarized transport of ena1 required for Saccharomyces cerevisiae survival against toxic cations[J].Molecular Biology of the Cell, 2017, mbc. E17-09-0549.
[50]Zhang C, Li Z, Zhang X, et al. Transcriptomic profiling of chemical exposure reveals roles of yap1 in protecting yeast cells from oxidative and other types of stresses[J]. Yeast, 2016, 33(1):5-19.
[51]Martínez JL, Liu L, Petranovic D, et al. Engineering the oxygen sensing regulation results in an enhanced recombinant human hemoglobin production by Saccharomyces cerevisiae[J].Biotechnol Bioeng, 2015, 112(1):181.
[52]Hubmann G, Thevelein JM, Nevoigt E. Natural and modified promoters for tailored metabolic engineering of the yeast Saccharomyces cerevisiae[M]. Springer New York, 2014.
[53]Williams TC, Averesch NJH, Winter G, et al. Quorum-sensing linked rna interference for dynamic metabolic pathway control in Saccharomyces cerevisiae[J]. Metab Eng, 2015, 29:124.
[54]Gueldener U, Heinisch J, Koehler GJ, et al. A second set of loxp marker cassettes for cre-mediated multiple gene knockouts in budding yeast[J]. Nucleic Acids Res, 2002, 30(6):e23.
[55]Redden H, Morse N, et al. The synthetic biology toolbox for tuning gene expression in yeast[J]. FEMS Yeast Res, 2014, 1 :1-10.
[56]Nevoigt E, Kohnke J, et al. Engineering of promoter replacement cassettes for fine-tuning of gene expression in Saccharomyces cerevisiae[J]. Appl Environ Microbiol, 2006, 8:5266-5273.
[57]Nevoigt E, Fischer C, Mucha O, et al. Engineering promoter regulation[J]. Biotechnol Bioeng, 2007, 96(3):550-558.
[58]张旭, 王晶晶, 刘建平. 基于启动子和宿主改造的酿酒酵母表达系统优化研究[J]. 中国生物工程杂志, 2015(1):61-66.
[59]Jensen PR, Hammer K. Artificial promoters for metabolic optimization[J]. Biotechnol Bioeng, 2015, 58(2-3):191-195.
[60]Johnson AN, Weil PA. Identification of a transcriptional activation domain in yeast repressor activator protein 1(rap1)using an altered DNA-binding specificity variant[J]. Journal of Biological Chemistry, 2017, 292(14):5705-5723.
[61]Jeppsson M, Johansson B, Jensen PR, et al. The level of glucose-6-phosphate dehydrogenase activity strongly influences xylose fermentation and inhibitor sensitivity in recombinant Saccharomyces cerevisiae strains[J]. Yeast, 2003, 20(15):1263.
[62]Rich MS, Payen C, et al. Comprehensive analysis of the sul1 promoter of Saccharomyces cerevisiae[J]. Genetics, 2016, 1 :191.
[63]Blazeck J, Garg R, Reed B, et al. Controlling promoter strength and regulation in Saccharomyces cerevisiae using synthetic hybrid promoters[J]. Biotechnol Bioeng, 2012, 109(11):2884-2895.
[64]Ruohonen L, Penttilä M, Keränen S. Optimization of bacillus alphaamylase production by Saccharomyces cerevisiae[J]. Yeast,1991, 7(4):337-346.
[65]Ruohonen L, Aalto MK, Keränen S. Modifications to the adh1 promoter of Saccharomyces cerevisiae for efficient production of heterologous proteins[J]. J Biotechnol, 1995, 3 :193-203.
[66]Denis CL, Ferguson J, Young ET. Mrna levels for the fermentative alcohol dehydrogenase of Saccharomyces cerevisiae decrease upon growth on a nonfermentable carbon source[J]. Journal of Biological Chemistry, 1983, 258(2):1165-1171.
[67]Wang S, Cheng G, Joshua C, et al. Furfural tolerance and detoxification mechanism in candida tropicalis[J]. Biotechnol Biofuels, 2016, 9(1):250.
[68]Kim S, Lee K, Bae SJ, et al. Promoters inducible by aromatic amino acids and γ-aminobutyrate(gaba)for metabolic engineering applications in Saccharomyces cerevisiae[J]. Appl Microbiol Biotechnol, 2015, 99(6):2705-2714.
[69]Leavitt JM, Tong A, Tong J, et al. Coordinated transcription factor and promoter engineering to establish strong expression elements in Saccharomyces cerevisiae[J]. Biotechnol J, 2016, 11(7):866.
[70]Malakar P, Venkatesh KV. Gal regulon of Saccharomyces cerevisiae performs optimally to maximize growth on galactose[J]. Fems Yeast Research, 2014, 14(2):346-356.
[71]唐瑞琪, 熊亮, 白凤武, 等. 酿酒酵母人工杂合启动子与天然启动子活性比较[J]. 生物技术通报, 2017(1):120-128.
[72]Garí E, Piedrafita L, Aldea M, et al. A set of vectors with a tetracycline-regulatable promoter system for modulated gene expression in Saccharomyces cerevisiae[J]. Yeast, 2010, 13(9):837-848.
[73]Wawiórka L, Molestak E, Szajwaj M, et al. The multiplication of ribosomal p-stalk proteins contributes to the fidelity of translation[J]. Mol Cell Biol, 2017, 37(17). Plie00060-17.
[74]Revankar SG, Fu J, et al. Cloning and characterization of the lanosterol 14α-demethylase(erg11)gene in cryptococcus neoformans[J]. Biochem Biophys Res Commun, 2004, 324(2):719-728.
[75]Blount BA, Weenink T, Ellis T. Construction of synthetic regulatory networks in yeast[J]. Febs Letters, 2012, 586(15):2112.
[76]Bellí G, Garí E, Piedrafita L, et al. An activator/repressor dual system allows tight tetracycline-regulated gene expression in budding yeast[J]. Nucleic Acids Res, 1998, 26(4):942-947.
[77]Shimizu-Sato S, Huq E, Tepperman JM, Quail PH. A lightswitchable gene promoter system[J]. Nature Biotechnology,2002, 20(10):1041-1044.
[78]Mcisaac RS, Gibney PA, Chandran SS, et al. Synthetic biology tools for programming gene expression without nutritional perturbations in Saccharomyces cerevisiae[J]. Nucleic Acids Res, 2014, 6 :e48.
[79]Hector RE, Mertens JA. A synthetic hybrid promoter for xyloseregulated control of gene expression in Saccharomyces yeasts[J].Molecular Biotechnology, 2017, 59(1):24-33.
[80]高义平, 赵和, 吕孟雨, 等. 易错PCR研究进展及应用[J].核农学报, 2013, 27(5):607-612.
[81]Jeppsson M, Johansson B, Jensen PR, et al. The level of glucose-6-phosphate dehydrogenase activity strongly influences xylose fermentation and inhibitor sensitivity in recombinant Saccharomyces cerevisiae strains[J]. Yeast, 2003, 20(15):1263-1272.
[82]Wang S, Cheng G, Joshua C, et al. Furfural tolerance and detoxification mechanism incandida tropicalis[J]. Biotechnol Biofuels, 2016, 9(1):250.
[83]余君涵, 马雯雯, 王智文, 等. 人工合成启动子文库研究进展[J]. 微生物学通报, 2016, 43(1):198-204.
[84]Zong Y, Zhang HM, Cheng L, et al. Insulated transcriptional elements enable precise design of genetic circuits[J]. Nature Communications, 2017, 8(1):52.
[85]Rohlhill J, Sandoval NR, Papoutsakis ET. Sort-seq approach to engineering a formaldehyde-inducible promoter for dynamically regulated escherichia coli growth on methanol[J]. Acs Synthetic Biology, 2017, 6(8):1584-1595.
