水稻OsMPK15的cDNA克隆和转录水平分析
2018-06-29石佳杨丹丹葛慧雯杜京尧梁卫红
石佳 杨丹丹 葛慧雯 杜京尧 梁卫红
(河南师范大学生命科学学院,新乡 453007)
促分裂原活化蛋白激酶(Mitogen-activated protein kinase,MAPK)是一类丝氨酸/苏氨酸(Thr/Ser)蛋白激酶,普遍存在于哺乳动物、高等植物和真菌等高等真核生物体内[1]。MAPK的结构和功能在进化中都是高度保守的。由MAPK介导的细胞信号转导参与了包括细胞分裂、细胞分化以及多种胁迫应答过程,并且MAPK还涉及激素介导的信号通路[2]。
研究显示,植物的MAPK家族参与了调控植物生长发育的过程,包括配子形成、胚胎发育、形态发生、衰老、脱落、受精和种子的形成[3]。如MAPK家族可以作为受体样激酶(Receptor-like kinase,RLKs)的下游信号分子参与调节植物的生长发育过程,其中对拟南芥AtMPK3和AtMPK6研究较为深入,其能作为RLKs的下游信号分子参与调节植物生长发育、细胞通讯过程[4-6]。有研究显示,烟草MAPK级联信号通路NPK1-NQK1-NRK1在调控细胞分裂中发挥重要的作用[7-9]。植物的MAPK还参与逆境胁迫以及激素信号应答。如拟南芥MAPK级联反应中,AtMKK1或AtMKK2通过激活下游的AtMPK4,不仅调控水杨酸(Salicylic acid,SA)的积累、活性氧(Reactive oxygen species,ROS)的平衡,还对低温、高盐和机械损伤等非生物胁迫也能作出响应[10-12]。
水稻MAPK同样参与了水稻的生长发育、生物胁迫应答和非生物胁迫应答过程,有些MAPK与调控植物激素信号通路有关,如OsMPK6的活性能被白叶枯病菌的侵染所激活,从而引起抗逆植物激素的积累,活化茉莉酸(Jasmonic acid,JA)和水杨酸信号通路,进而上调抗病基因的表达量,增强植株局部对白叶枯病菌的抗性[13-14]。OsMPK5参与水稻对高盐、干旱、紫外、臭氧、重金属、高温及低温胁迫等几乎所有非生物胁迫应答过程[15-16];OsMEK1-OsMAP1级联通路则广泛参与了水稻的低温胁迫应答过程[17]。尽管许多MAPK参与水稻的生物与非生物胁迫应答及生长发育过程[2],但其生物学功能和作用机制仍有待深入研究。
本实验室前期对水稻MAPK家族成员OsMPK14的研究显示,该基因参与多种水稻非生物胁迫应答过程[18],水稻功能未知基因OsMPK15与其同源性最高。为此,本研究拟克隆OsMPK15的cDNA编码区,并通过qRT-PCR技术检测OsMPK15的表达特性,旨为后续的功能研究奠定基础。
1 材料与方法
1.1 材料
水稻材料为粳稻品种日本晴(Oryza sativa L.
japonica.cv. Nipponbare)。克隆载体pEASY-T载体和高纯度质粒小提试剂盒购自北京全式金有限公司,大肠杆菌(Escherichia coli)DH5α由河南师范大学生命科学学院植物发育分子细胞生物学实验室保存。TRIzol总RNA提取试剂、反转录试剂盒、荧光定量试剂盒 SYBR PrimeScript RT-PCR Kit、SYBR®Premix Ex TaqTM、限制性内切酶均购自大连宝生物公司;植物激素脱落酸(Abscisic acid,ABA)、水杨酸、茉莉酸、聚乙二醇(Polyethylene glycol,PEG6000)、氯化钠(Sodium chloride)、氨苄霉素(Ampicillin)购于北京鼎国生物公司。
1.2 方法
1.2.1 水稻幼苗的种植及处理 取日本晴水稻种子若干置于小烧杯中,用75%的酒精消毒30 s,再用无菌水冲洗5-6次,每次冲洗1 min。用无菌水完全淹没种子,浸泡1 h。将种子转移至培养皿中,加入适量的水(不能完全淹没种子),28℃黑暗培养2-3 d。待种子长出幼芽后,采用人工气候箱进行培养,培养条件为28℃、光照强度16 000 lx、光照时间为16 h/d。将水培生长15 d的水稻幼苗转移到培养皿中。待水稻幼苗适应2 d后,分别用含有150 mmol/L NaCl(盐胁迫)、30% PEG6000(模拟干旱胁迫)、100 μmol/L JA、100 μmol/L SA 和 100 μmol/L ABA的Yoshida培养液[19]处理水稻幼苗,处理方法参照张静等[20]方法进行。分别在处理0、3、6、9、12和24 h后取水稻幼苗的根组织适量,经液氮速冻后,保存于-80℃超低温冰箱中备用。
1.2.2 水稻的大田种植及取材 试验田平整、浸泡作为苗床育苗,2014年5月1日将种子分散播种至苗床,加水淹没苗床,2014年5月25日进行分苗,在水稻生长至幼穗期取其根、茎、叶、叶鞘、幼穗(生长至1-2 cm、3-5 cm、5-8 cm)组织,液氮速冻后保存于-80℃冰箱。
1.2.3 总RNA的提取和cDNA的合成 采用TRizon法提取上述组织的总RNA,经快速电泳,检测确认后,按照反转录试剂盒说明书,配制反转录反应体系,制备cDNA。
1.2.4 水稻OsMPK15的cDNA编码区的克隆 以上述制备的水稻根组织的cDNA为模板,用根据水稻OsMPK15编码框设计引物P1/P2,扩增OsMPK15的cDNA编码区。PCR反应体系为cDNA 1 μL、上下游引物各 0.5 μL、dNTP 2.5 μL、5×Taq buffer 5 μL、Taq酶 0.5 μL,加 ddH2O 至终体积 50 μL;反应程序为 95℃ 2 min ;95℃ 20 s,53℃ 20 s,72℃ 1 min,40个循环;72℃ 5 min;4℃保存。以1%的琼脂糖凝胶电泳检测PCR产物,采用快速琼脂糖凝胶DNA回收试剂盒回收目的条带。取3 μL回收产物与pEASY-T载体(含T4连接酶)连接后转入大肠杆菌DH5α感受态细胞,将转化产物涂布于含有100 μg/mL Ampicillin的LB固体培养基上,于37℃恒温培养箱倒置过夜培养。挑取LB固体培养基的单克隆,接种至5 mL含有100 μg/mL Ampicillin的LB液体培养基中,37℃,200 r/min,震荡培养过夜,双酶切鉴定后测序。将测序获得的OsMPK15的cDNA编码区提交至 NCBI(http://www.ncbi.nlm.nih.gov/)进行序列分析和同源检索。采用DNAMAN软件进行序列比对,绘制系统发育树。引物(表1)均委托苏州金唯智生物公司合成。
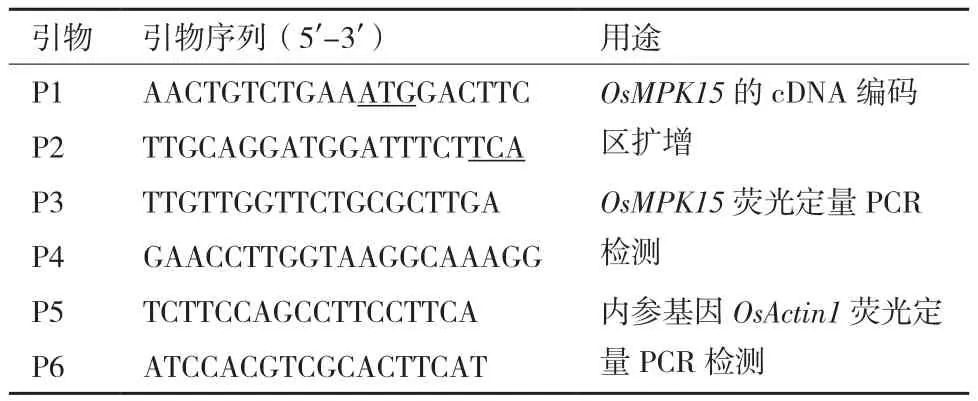
表1 引物序列信息
1.2.5 水稻OsMPK15不同组织中的表达分析 以组成型表达的OsActin1为内参基因进行实时定量荧光PCR检测,以反转录制备的各个组织样品的cDNA为模板,采用P3/P4和P5/P6引物组合分别扩增OsMPK15和OsActin1。荧光定量反应体系为SYBR®Premix Ex TaqTM(2×)10 μL,上下游引物(10 μmol/L) 各 0.4 μL,ROX Reference Dye Ⅱ(50×)0.4 μL,cDNA 2 μL,加 ddH2O 至终体系 20 μL。使用ABI 7500 实时PCR仪(Applied Biosystems)进行PCR扩增。反应采用两步法PCR扩增标准程序95℃30 s;95℃ 5 s;60℃ 30 s,40个循环。每个组织进行3次重复,采用2-△△Ct法分析结果,数据统计采用Excel软件,显著性分析采用Spass v19.0软件。
1.2.6 OsMPK15对非生物胁迫和激素的响应分析取经过干旱、高盐、激素处理的水稻幼苗根组织,以组成型表达的OsActin1为内参基因,进行OsMPK15的qRT-PCR分析,操作同1.2.5。
2 结果
2.1 水稻OsMPK15的cDNA编码区的克隆
以提取的总RNA反转录得到的cDNA为模板,P1/P2为引物,扩增得到1条约1 500 bp的特异性片段,与预期的OsMPK15序列的cDNA编码区序列大小符合(图1-A)。将上述PCR产物进行胶回收,与pEASY-T载体连接。将连接产物转化大肠杆菌,筛选转化子。挑取若干单菌落分别扩大培养,提取质粒进行双酶切鉴定(图1-B)。将筛选到的阳性克隆进行测序,测序结果显示,通过RT-PCR技术克隆的OsMPK15序列与文献推测的序列[21]一致,OsMPK15的cDNA编码区长1 497 bp,预期编码498个氨基酸和1个终止密码子。

图1 OsMPK15的cDNA编码区的扩增和克隆
2.2 水稻OsMPK15的序列分析
将OsMPK15的cDNA编码区序列提交至NCBI,分析其编码蛋白的保守结构域,发现该蛋白在第13-304处的氨基酸对应MAPK特有的丝氨酸/苏氨酸蛋白激酶结构域。利用DNAMAN软件,对水稻MAPK家族E组的3个成员OsMPK13、OsMPK14和OsMPK15进行比对(图2),发现尽管构成3种蛋白的氨基酸数目不同,但是序列同源性很高,尤其是N端序列相似性很高,其序列差异主要体现在C端;且丝氨酸/苏氨酸蛋白激酶结构域具有高度保守性,都具有TDY基序。其中OsMPK14和OsMPK15蛋白同源性达到82.87%。
将OsMPK15蛋白质序列提交至NCBI进行BLAST检索,选择其中11种同源性较高的,来自花生、番茄、葡萄、卷柏、小麦、拟南芥、烟草、棉花及苜蓿的MAPK,采用DNAMAN软件进行多序列比对分析,结果(图3)显示,与水稻OsMPK15相似性最高的是小麦TaMPK2,同源性高达79.78%,提示二者可能是同源基因。
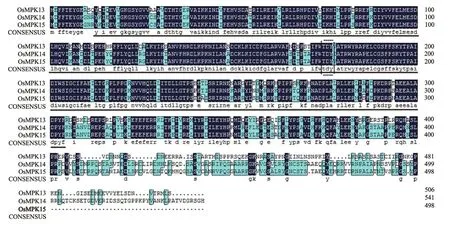
图2 水稻MAPK家族E组3个成员的氨基酸序列比对

图3 基于水稻OsMPK15与其他植物MAPK序列构建的系统发育树
2.3 OsMPK15在幼穗期水稻中的表达特性
以组成型表达的OsActin1为内参基因,通过荧光定量PCR检测OsMPK15在幼穗期水稻根、茎、叶、叶鞘以及不同发育阶段幼穗组织中的相对表达水平。结果(图4)显示,OsMPK15在水稻幼穗期所检测的各组织中普遍表达,但表达水平存在差异。其中在叶和叶鞘中的表达量显著高于在其他组织中的表达量,是其他组织中表达量的4-6倍。
2.4 激素对OsMPK15在幼苗根中表达的影响
以JA、SA和ABA分别处理15 d的水稻幼苗,以水稻OsActin1作为内参基因,采用qRT-PCR技术检测OsMPK15在3种激素处理后的根中24 h之内的表达变化。结果(图5)显示,OsMPK15对3种激素均有应答,在检测的24 h内,均出现表达水平上调,而后恢复的趋势,但是表达峰值出现的时间不同,其中JA处理12 h后其表达上调了4倍,SA处理3 h后表达上调了4倍,ABA处理9 h后其表达上调了7倍,提示该基因对这些激素的敏感度和应答模式存在差异。
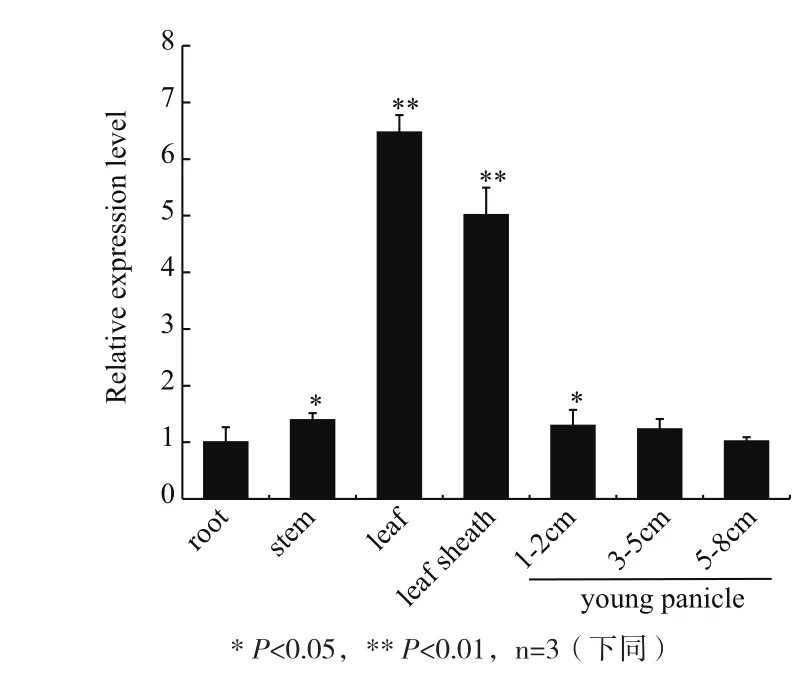
图4 OsMPK15在水稻幼穗期各组织中的表达
2.5 非生物胁迫对OsMPK15基因表达的影响
对水稻幼苗进行模拟干旱和高盐处理,采用实时荧光PCR检测该基因在根中24 h之内的表达变化,发现在模拟干旱的胁迫下,OsMPK15的表达量出现明显上调,在处理的12 h内其表达量维持在0 h的3倍左右,随后呈现下降趋势;在高盐处理条件下,其表达量也有上调,在9 h内表达量逐渐上升,达到2倍后呈现下降趋势(图6)。结果表明,干旱和高盐对OsMPK15的表达都有影响,但是相比之下,干旱对该基因表达的影响更为显著。

图5 JA、SA和ABA激素处理对OsMPK15基因表达的影响

图6 干旱和盐胁迫对OsMPK15表达的影响
3 讨论
无论酵母还是动植物的MAPK在进化过程中都高度保守,具有相似的保守结构。与动物的细胞外调节蛋白激酶(Extracellular regulated protein kinases,ERK)相似,植物MAPK活性环上同样具有TDY或TEY基序[21]。同样,依据活性环上特殊基序TXY的种类(TEY/TDY),将水稻MAPK划分为A-F 7组,本研究通过RT-PCR技术克隆的OsMPK15序列与文献推测的序列[22]一致,预期编码的蛋白包含498个氨基酸,属于E组成员。与之同属E组的OsMPK13已被证实参与苯并噻二唑(Benzothiadiazole,BTH)诱导的水稻抗病反应[23],OsMPK14在水稻的非生物胁迫与激素应答调控过程中具有重要作用[22],但对OsMPK15的研究尚未见报道。
不良的土壤条件如干旱、高盐等是制约水稻产量的主要因素。已有研究显示,MAPK在植物应答多种非生物胁迫的过程中扮演了重要的角色。例如,在拟南芥中,低温、高盐、机械损伤等可以显著提高AtMEKK1的转录水平[24];低温、干旱、高渗、创伤等可以激活AtMPK4和AtMPK6的瞬时表达[25]。水 稻 中 OsMPK4、OsMPK5、OsMPK7、OsMPK8和OsMPK12等都可以被干旱、盐或者温度的变化而诱导表达[26-28]。为了研究非生物胁迫对水稻OsMPK15基因表达的影响,本研究模拟了干旱和高盐胁迫,并利用实时荧光定量PCR技术进行了检测。发现在干旱和盐胁迫下,OsMPK15在根中的表达量均出现上调。相对于盐胁迫,OsMPK15对干旱胁迫更敏感。有报道显示,与OsMPK15同源性极高的OsMPK14和小麦TaMPK2均参与了非生物胁迫和激素的应答调控过程[22]。已有研究显示,外源ABA影响了多种植物MAPK基因的表达、蛋白的积累以及酶的活性,MAPK还参与了ABA介导的多种信号通路,包括氧化防御、保卫细胞信号转导和种子的萌发[29-32]。在拟南芥中,JA和SA可以诱导AtMPK6和AtMPK3的表达[33-34]。本研究发现,OsMPK15对激素ABA、JA和SA均有响应,其中以ABA诱导OsMPK15表达上调幅度最大。鉴于ABA是植物逆境胁迫相关激素,OsMPK15对ABA处理产生高度的应答反应,同样暗示该基因可能参与了水稻的逆境胁迫应答过程。本研究还发现,OsMPK15的表达与实验室前期报道的OsMPK14基因的表达模式十分相似,但也存在显著差异,如OsMPK14的表达在一定程度上受到干旱胁迫的抑制作用[18],这些结果说明这两种水稻MAPK与水稻的非生物胁迫应答有密切联系。此外,OsMPK15在幼穗期的水稻各组织中在叶和叶鞘中优势表达,OsMPK14在水稻地上部分的表达受到光的负调控[18],提示它们也可能与水稻的光信号通路存在关联,值得进一步的深入研究。
本研究结果提示,水稻OsMPK15可能与水稻的非生物胁迫应答、激素信号通路以及光信号通路有所关联,但其具体功能尚未见报道,应答机制也有待研究。目前,实验室已通过CRISPR-Cas9基因组编辑技术构建了OsMPK15敲除水稻,后续将开展对OsMPK15与水稻非生物胁迫应答、激素应答、光信号通路关系的研究,以期对其功能开展进一步的分析鉴定,为揭示其在体内的作用机制提供理论依据。
4 结论
克隆了水稻OsMPK15的cDNA编码区,发现该基因在不同组织中表达存在差异,能够被ABA、SA、JA高度诱导,同时受干旱和盐胁迫影响,推测该基因可能与水稻的抗逆应答过程密切相关。
[1]Nam-Soo J. The rice MAPKK-MAPK interactome:the biological significance of MAPK components in hormone signal transduction[J]. Plant Cell, 2013, 32:923-931.
[2]Huang HJ, Fu SF, Tai YH, et al. Expression of Oryza sativa MAP kinase gene is developmentally regulated and stressresponsive[J]. Physiology Plant, 2002, 114:572-580.
[3]Krishna M, Narang H. The complexity of mitogen-activated protein kinases(MAPKs)made simple[J]. Cellular and Molecular Life Science, 2008, 65(22):3525-3544.
[4]De Smet I, Voss U, Jürgens G et al. Receptor-like kinases shape the plant[J]. Cell Biology, 2009, 11 :1166-1173. .
[5]Li J, Tax FE. Receptor-like kinases:key regulators of plant development and defense[J]. Plant Biol, 2013, 55:1184-1187.
[6]Gish LA, Clark SE. The RLK/Pelle family of kinases[J]. Plant J,2011, 66:117-127.
[7]Nishihama R, Ishikawa M, et al. The NPK1 mitogen-activated protein kinase kinase kinase is a regulator of cell-plate formation in plant cytokinesis[J]. Genes Dev, 2001, 3 :352-363.
[8]Soyano T, Nishihama R, et al. NQK1/NtMEK1 is a MAPKK that acts in the NPK1 MAPKKK-mediated MAPK cascade and is required for plant cytokinesis[J]. Genes Dev, 2003, 17:1055-1067.
[9]Wen JQ, Oono K, Imai R. Two novel mitogen-activated protein signaling componets, OsMEK1 and OsMAP1, are involved in a moderate low-temperature signaling pathway in rice[J]. Plant Physiology, 2002, 129(4):1880-1891.
[10]Hadiarto T, Nanmori T, Matsuoka D, et al. Activation of Arabidopsis MAPK kinase(AtMEKK1)and induction of AtMEKK1-AtMEK1 pathway by wounding[J]. Planta, 2006, 223(4):708-713.
[11]Teige M, et al. The MKK2 pathway mediates cold and salt stress signaling in Arabidopsis[J]. Mol Cell, 2004, 1 :141-152.
[12]Pitzschke A, Djamei A, Bitton F, et al. A major role of the MEKK1-MKK1/2-MPK4 pathway in ROS signaling[J]. Molecular Plant,2009, 2(1):120-137.
[13]Yuan B, Shen X, Li X, et al. Mitogen-activated protein kinase OsMPK6 negatively regulates rice disease resistance to bacterial pathogens[J]. Planta, 2007, 226(4):953-960.
[14]Shen X, Yuan B, Liu H, et al. Opposite functions of a rice mitogenactivated protein kinase during the process of resistance against Xanthomonas oryzae[J]. Plant J, 2010, 64(1):86-99.
[15]Xiong L, Yang Y. Disease resistance and abiotic stress tolerance in rice are inversely modulated by an abscisic acid-inducible mitogenactivated protein kinase[J]. Plant Cell, 2003(3):745-759.
[16]Agrawal GK, et al. Isolation of novel rice(Oryza sativa L.)multiple stress responsive MAP kinase gene, OsMSRMK2, whose mRNA accumulates rapidly in response to environmental cues[J].Biochem Biophys Res Commun, 2002, 5:1009-1016.
[17]Wen JQ, Oono K, Imai R. Two novel mitogen-activated protein signaling components, OsMEK1 and OsMAP1, are involved in a moderate low-temperature signaling pathway in rice[J]. Plant Physiology, 2002, 129(4):1880-1891.
[18]梁卫红, 等. 水稻促分裂原活化蛋白激酶基因OsMPK14的克隆及表达分析[J]. 中国水稻科学, 2010(2):125-130.
[19]Yoshida S, Forno DA, Cock JH, et al. Laboratory manual for physiological studies of rice[M]. Manila:International Rice Research Institute, 1976.
[20]张静, 梁卫红. 2种非生物胁迫和7种激素对水稻OsAQP基因表达的影响[J]. 河南师范大学学报:自然科学版, 2016,44(1):105-109.
[21]Jiang M, Wen F, Cao JM, et al. Genome-wide exploration of the molecular evolution and regulatory network of mitogen-activated protein kinase cascades upon multiple stresses in Brachypodium distachyon[J]. BMC Genomics, 2015, 16:228.
[22]Reyna NS, Yang Y. Molecular analysis of the rice MAP kinase gene family in relation to Magnaporthe grisea infection[J]. Molecular Plant-Microbe Interactions, 2006, 19(5):530-540.
[23]Song D, et al. A novel rice MAPK gene, OsBIMK2, is involved in disease-resistance responses[J]. Plant Biol, 2006, 5:587-596.
[24]Mizoguchi T, Irie K, et al. A gene encoding a mitogen-activated protein kinase kinase kinase is induced simultaneously with genes for a mitogen-activated protein kinase and an S6 ribosomal protein kinase by touch, cold, and water stress in Arabidopsis thaliana[J]. Proc Natl Acad Sci USA, 1996, 93:765-769.
[25]Ichimura K, Mizoguchi T, Yoshida R, et al. Various abiotic stresses rapidly activate Arabidopsis MAP kinases ATMPK4 and ATMPK6[J]. Plant J, 2000, 24:655-665.
[26]Jeong M, Park S, Lee S, et al. A rice(Oryza sativa L.)MAP kinase gene, OsMAPK44, is involved in response to abiotic stresses[J].Plant Cell Tissue Organ Cult, 2006, 85(2):151-160.
[27]Agrawal GK, Agrawal SK, Shibato J, et al. Novel rice MAP kinases OsMSRMK3 and OsWJUMK1 involved in encountering diverse environmental stresses and developmental regulation[J].Biochem Biophys Res Commun, 2003, 300(3):775-783.
[28]Agrawal GK, Tamogami S, Iwahashi H, et al. Transient regulation of jasmonic acid-inducible rice MAP kinase gene(OsBWMK1)by diverse biotic and abiotic stresses[J]. Plant Physiology and Biochemistry, 2003, 41(4):355-361.
[29]Jammes F, Song C, Shin D, et al. MAP kinases MPK9 and MPK12 are preferentially expressed in guard cells and positively regulate ROS-mediated ABA signaling[J]. Proc Natl Acad Sci USA,2009, 106:20520-20525.
[30]Xing Y, Jia W, Zhang J. AtMKK1 mediates ABA-induced CAT1 expression and H2O2production via AtMPK6-coupled signaling in Arabidopsis[J]. Plant J, 2008, 54 :440-451.
[31]Zhang A, Zhang J, Ye N, et al. ZmMPK5 is required for the NADPH oxidase-mediated self-propagation of apoplastic H2O2in brassinosteroid-induced antioxidant defence in leaves of maize[J]. J Exp Bot, 2010, 61:4399-4411.
[32]Zong XJ, Li DP, Gu LK, et al. Abscisic acid and hydrogen peroxide induce a novel maize group C MAP kinase gene, ZmMPK7, which is responsible for the removal of reactive oxygen species[J].Planta, 2009, 229:485-495.
[33]Zhang S, Klessig DF. Salicylic acid activates a 48-kD MAP kinase in tobacco[J]. Plant Cell, 1997, 9(5):809-824.
[34]Seo S, Katou S, Seto H, et al. The mitogen-activated protein kinases WIPK and SIPK regulate the levels of jasmonic and salicylic acids in wounded tobacco plants[J]. Plant J, 2007, 49(5):899-909.
