江西官山乐昌含笑群落组成、种群动态与自然择优
2018-06-29宋述灵姚小华余泽平林国江杨清培
宋述灵,姚小华,余泽平,陈 琳,林国江,杨清培*
(1.江西农业大学 江西省竹子种质资源与利用重点实验室,江西 南昌 330045;2.江西官山国家级自然保护区管理局,江西 宜丰 336300)
珍稀植物是生物多样性的重要组成部分,对生态系统功能的发挥起着不容忽视的作用[1]。许多野生资源正急剧减少,加强天然林群落的长期定位监测,对珍稀濒危物种保护与恢复具有重要意义[2]。
乐昌含笑(Micheliachapensis)又名南方白兰花,木兰科含笑属乔木,喜温暖湿润气候[3],自然种群规模较小,是我国重点保护珍稀濒危植物[4]。一直以来,乐昌含笑作为优良的绿化和用材树种,野外资源过度开发,原始林面积快速下降,呈间断零散分布,野生种群逐年衰退[5]。前人在分子遗传、生理特性、种群分布等理论方面[6-8],以及引种栽培、品种选育等应用领域进行了大量研究[9,10],但自然状态下乐昌含笑群落生态学研究较少,缺乏长期跟踪[11]。由于受尺度变化影响,森林植被群落组成、结构及个体的空间分布格局不同[12],短时间尺度很难反映群落水平上的物种多样性格局和种群发育过程,不足以认识种群的发展规律和种间关系的形成机制[13-14]。
定位长期监测是最有效的生态学野外研究方法。“定位、定时、定身份”的特点能够保证研究的可重复性、长期跟踪性和综合性,能为解决相关科学问题提供最基本和最详尽的科学数据[15-16]。胸径、树高与多度等是植物群落监测的基础指标,能客观反映个体生长、种群更新、群落结构和发育过程[16-17]。官山自然保护区地处九岭山脉西段,位于鄱阳湖、江汉和洞庭湖3大平原的中心位置,是全球生物多样性优先保护地区[18]。优越的自然条件孕育了丰富的野生珍稀植物资源[19],该保护区也是全国重要的乐昌含笑种质资源建设基地,分布着群落结构完整、面积相对较大的天然林群落[18]。因此,为了解乐昌含笑天然林群落现状和今后种群整个发育过程,在江西官山建立乐昌含笑天然林长期监测样地,开展前期群落本底调查研究。本文依据第一次调查结果重点探讨:(1)物种组成及数量特征;(2)群落的空间结构;(3)木本植物多样性水平;(4)种群更新状况,拟为后期探讨乐昌含笑群落长期生态学研究奠定基础,也为该珍稀濒危物种保护提供科学依据。
1 材料和方法
1.1 研究区概况
江西官山自然保护区位于赣西北九岭山脉西段的宜丰、铜鼓两县境内(28°30′~28°40′N;14°29′~114°45′E),是我省首批建立的国家级自然保护区。总面积11 500.5 hm2,其中核心区3 621.1 hm2,缓冲区1 466.4 hm2,实验区6 413.0 hm2。该区属中亚热带温暖湿润气候区,具有四季分明、光照充足、无霜期长等特点[20]。年均气温16.2 ℃,年均降水量1 950~2 100 mm,其地带性土壤为红壤[19]。由于独特的地理位置和良好的生态环境,官山孕育了丰富的自然资源,素有九岭山翡翠之称,分布有典型的亚热带常绿阔叶林,其中珍稀物种资源种类繁多,具有国家重点保护野生植物21种和国家级珍稀濒危保护植物28种,分别占江西省的38.1%和51.8%[18]。同时,保护区内天然分布着较大面积的乐昌含笑群落,为长期动态监测研究提供了理想的实验场所。
1.2 样地设置与调查
2016年7月,经实地踏查,根据官山乐昌含笑群落分布特征和资源总量,在保护区内选择典型的乐昌含笑天然林,通过全站仪建立了3个永久性固定样地,编号分别为GS-MC1(20 m×20 m)、GS-MC2(20 m×30 m)、GS-MC3(50 m×50 m),开展群落的第一次本底调查并记录每个样地海拔、坡度、坡向、乔木层郁闭度等基本信息(表1)。用相邻格子法进行每木调查,以10 m×10 m为基本单位,将样地内所有胸径(DBH)≥1 cm的木本或藤本植物用铝制标牌进行标记,并记录植物种名、胸径、高度、枝下高、光照和生长状况等[11,21-23],DBH≥1 cm的萌枝、分枝也按照同样方法挂牌测量,并记录萌枝、分枝的母株编号[15]。

表1 样地基本情况Tab.1 Basic information of plots
1.3 数据处理与分析
1.3.1 物种重要值计算 重要值= (相对密度+相对频度+相对显著度+相对高度)/4
(1)
其中,相对密度为某个物种的个体数目与样方中全部物种的个体数目总和的比值;相对频度为某物种的频度占全部物种的频度之和的比值,它是反映某种植物分布均匀程度的一个指标;相对显著度为某个物种的胸高断面积总和与样方内所有物种的胸高断面积总和的比值;相对高度为某物种高度占所有物种高度总和的比值[24-25]。
1.3.2 群落结构测度 垂直结构根据群落植物个体高度分布范围,划分5个等级,每个等级间隔5 m[26]。第Ⅰ高度级为0
径级结构:将群落内植物径级按5 cm的组间距分成6个等级。第Ⅰ组为0
1.3.3 种群年龄结构及静态生命表编制方法 种群年龄结构以立木径级结构代替,按5 cm的组间距分为5个等级。以龄级为横坐标,各龄级个体数为纵坐标,绘制官山乐昌含笑种群年龄结构图。种静态生命表以大小级代替龄级编制,详细编制方法见参考文献[28-29]。以龄级为横坐标,以标准化存活数lx为纵坐标绘制存活曲线;以龄级为横坐标,死亡率qx为纵坐标,绘制死亡率与致死率曲线。
1.3.4 物种多样性测度[30-31]物种丰富度:R=S
(2)
(3)
(4)
(5)
(6)
(7)
其中,S为物种总数目,N为所有物种的个体数之和,ni为第i个种的个体数,Pi为第i个种的个体数占所有个体总数N的比例。
2 结果与分析
2.1 物种组成与数量特征
2.1.1 种类组成 乐昌含笑天然林群落的物种组成较丰富(表2)。3个样地共有1 085株121种,隶属72属43科。其中GS-MC1有85株16种14属12科;样地GS-MC2中分布有147株29种22属16科以及样地GS-MC3中包括853株115种67属42科。另外,群落中灌木层的物种数均大于乔木层,样地GS-MC1、GS-MC2和GS-MC3的乔木层种数分别为7、15和59种;灌木层分别包含14、19和92种。上述物种组成数量的差异主要原因在于样地面积不同。
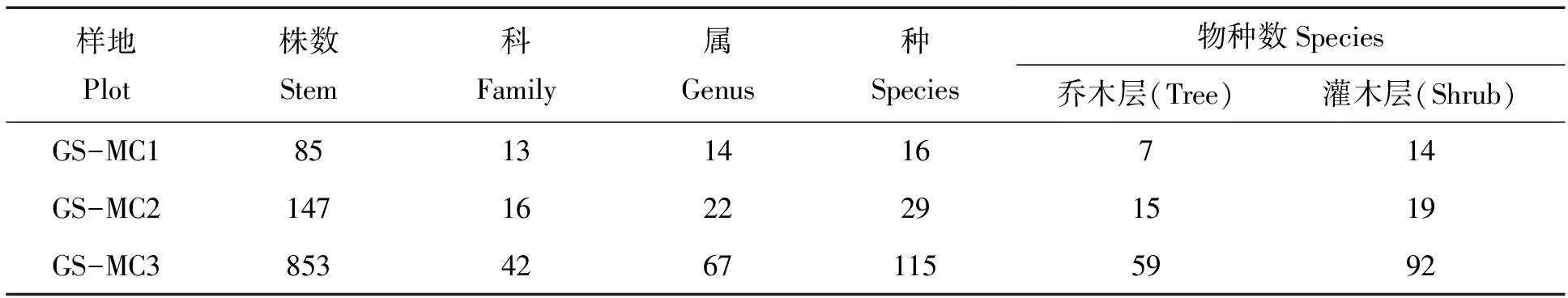
表2 乐昌含笑天然林群落科属种数量情况Tab.2 Number of species,genus,family and stems in Michelia chapensis community
2.1.2 重要值分析 乐昌含笑在3个群落中的优势地位较明显,是群落中的建群种。样地GS-MC1乔木层处于前3位的是乐昌含笑(31.03)、三峡槭(Acerwilsonii)(21.92)和钩栲(Castanopsistibetana)(19.21),重要值之和高达72.17,灌木层优势种为尾叶山茶(Camelliacaudat)和乐昌含笑,重要值分别是28.74和14.23。样地GS-MC2中乔木层优势种乐昌含笑重要值达27.46,伴生种包括钩栲(12.63)、木荷(Schimasuperba)(11.94)和多花泡花树(Meliosmamyriantha)(8.13),灌木层中茜树(Aidiacochinchinensis)、尾叶山茶、乐昌含笑的重要值位居前3,分别为21.62、16.90和11.59。样地GS-MC3是以乐昌含笑和麻栎(Quercusacutissima)群落乔木层共优种,其重要值为15.30和13.18,灌木层伴生种主要有乐昌含笑、拟赤杨、三峡槭、钩栲和百齿卫矛(Euonymuscentidens)等,麻栎在灌木层中没有出现,样地中乐昌含笑的最大胸径高达61.30 cm,最大高度为27 m。所有样地中胸径最大为麻栎,高达71 cm(表3)。
2.2 群落结构
2.2.1 垂直结构 3个样地群落垂直结构较相似,皆表现为随着高度级增加,树木个体数百分比呈逐渐下降(图1)。如0~5 m、5~10 m、10~15 m、15~20 m和大于20 m的高度区间,样地GS-MC1树木多度百分比分别为55.29%、27.06%、1.18%、10.59%和5.88%;样地GS-MC2中树木多度百分比分别为75.51%、10.20%、7.48%、4.08%和2.72%;样地GS-MC3中树木多度百分比分别为70.11%、16.88%、7.50%、3.17%和2.34%。以上说明,乐昌含笑群落具有相似的垂直分布结构,树木高度集中分布于0~10 m。
2.2.2 径级结构 同样,如同群落垂直结构,3个样地群落径级结构大体相似,树木多度百分比随着径级增加,大体呈逐渐下降趋势(图2)。在0~5 cm胸径区间的多度百分比均最大,3个样地分别为63.53%、73.47%和71.98%。在5~10 cm、10~15 cm、15~20 cm、20~25 cm、大于25 cm区间,样地GS-MC1中的树木多度百分比分别为16.47%、3.53%、7.06%、0.00%和9.41%,样地GS-MC2则分布为4.76%、7.48%、3.40%、2.72%和8.16%,样地GS-MC3中树木多度百分比分别为11.02%、6.92%、5.28%、1.76%和3.05%。
2.3 群落物种多样性
随样地面积增大,物种多样性呈上升趋势(表4)。乔木层Magalef指数分别为1.75、3.82和10.59,灌木层分别为3.26、3.84和14.17。乔木层的Simpson指数增幅较大,由样地GS-MC1中的5.41增加至样地GS-MC3 中的12.07,增幅123%,灌木层Simpson指数增幅为77%。样地GS-MC1、GS-MC2和GS-MC3的Shannon-Wiener指数,乔木层分别为1.69、2.37和3.32,灌木层为2.09、2.34和3.72。3个样地Pielou均匀度指数较高,范围在0.79~0.87,而生态优势度指数均较低,仅为0.04~0.21,且灌木层多样性皆大于乔木层。

表3 乐昌含笑天然林群落重要值排前10物种Tab.3 Importance value (IV) of the top 10 species in M. chapensis community
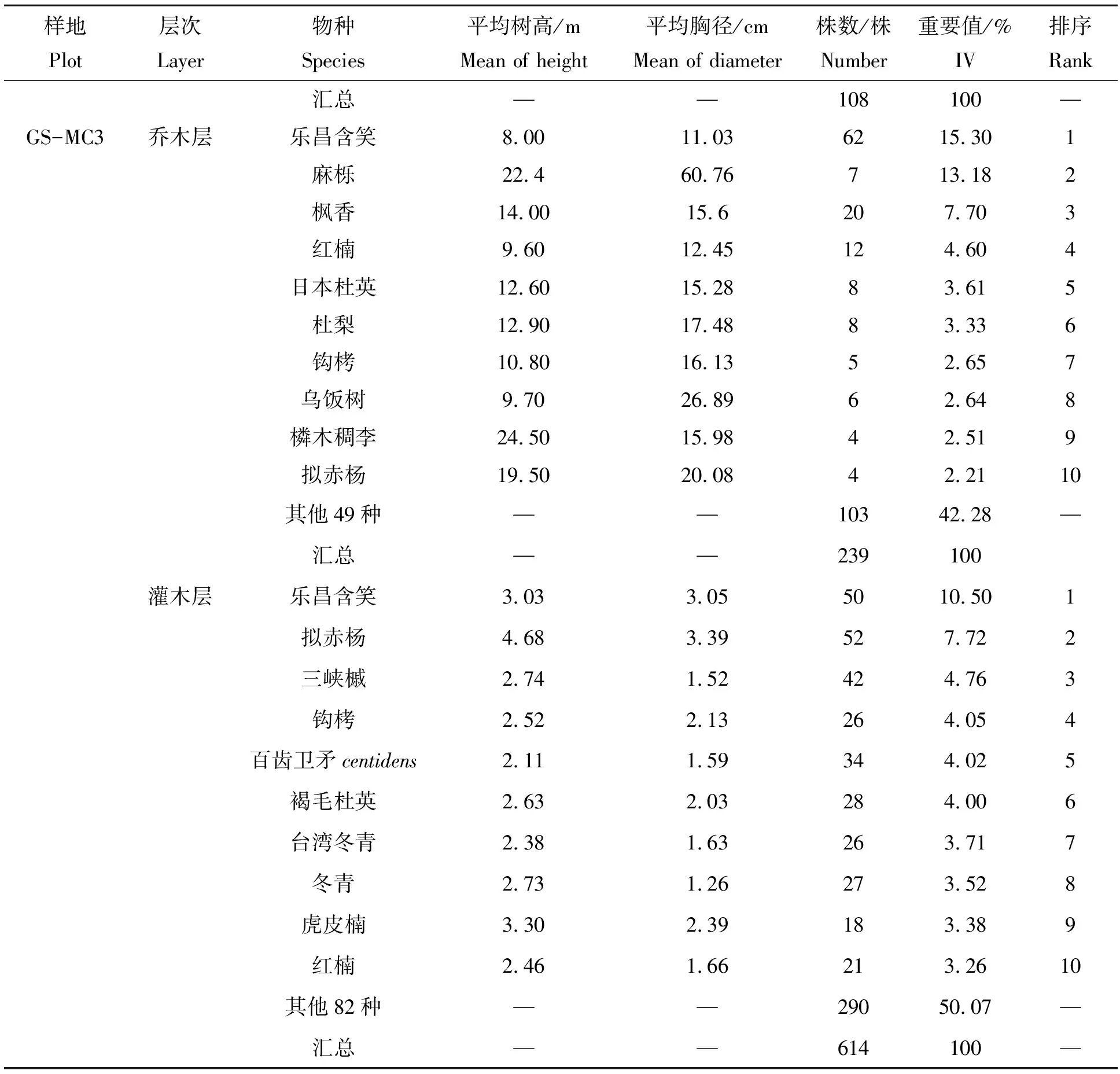
续表3
乐昌含笑Micheliachapensis;三峡槭Acerwilsonii;钩栲Castanopsistibetana;橉木稠李Padusracemose;中华卫矛Euonymusnitidus;榕叶冬青Ilexficoidea;尾叶山茶Camelliacaudat;茜树Aidiacochinchinensis;细枝柃Euryaloquaiana;多花泡花树Meliosmamyriantha;粗糠柴Mallotusphilippensis;三尖杉Cephalotaxusfortune;木荷Schimasuperba;枫香Liquidambarformosana;拟赤杨Alniphyllumfortune;南酸枣Choerospondiasaxillaris;鼠刺Iteachinensis;刺叶桂樱Laurocerasusspinulosa;日本杜英Elaeocarpusjaponicas;台湾冬青Ilexformosana;绿冬青Ilexviridis;麻栎Quercusacutissima;红楠Machilusthunbergii;杜梨Pyrusbetulifolia;乌饭树Vacciniumbracteatum;百齿卫矛Euonymuscentidens;褐毛杜英Elaeocarpusduclouxii;冬青Ilexchinensis;虎皮楠Daphniphyllumoldhami

图1 乐昌含笑群落垂直结构Fig.1 Vertical structure of species in M. chapsensis community

图2 乐昌含笑群落径级分布Fig.2 Diameter distribution of species in M. chapsensis community
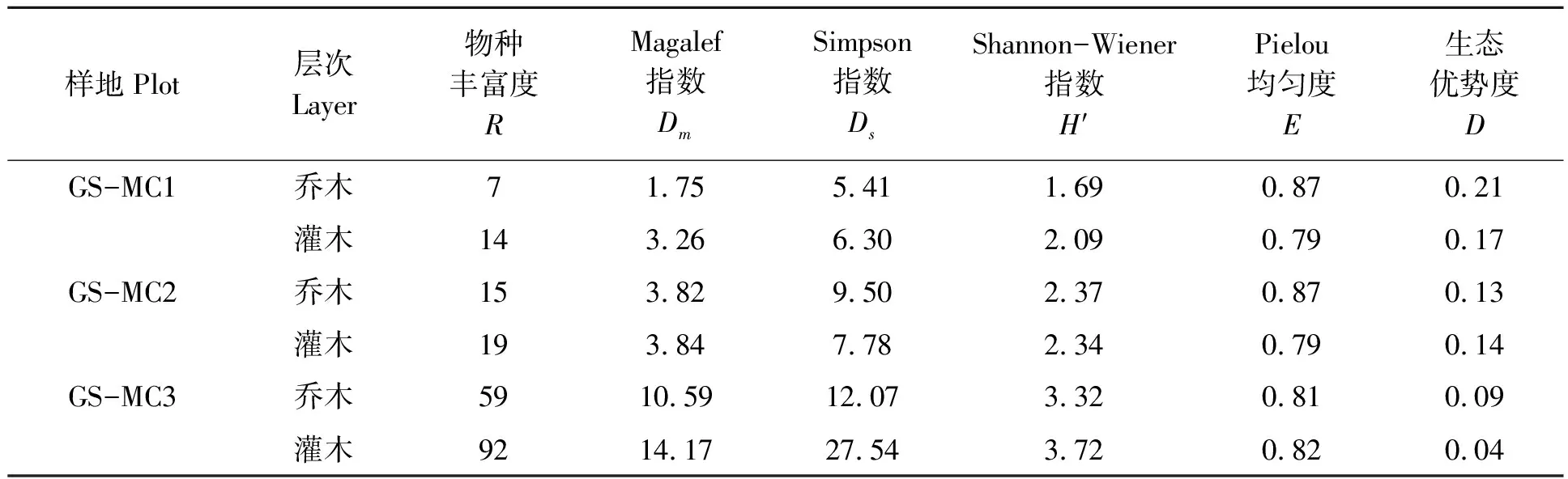
表4 乐昌含笑群落物种多样性指数分析Tab.4 Diversity indexes of M. chapsensis community
2.4 种群年龄结构

图3 乐昌含笑种群年龄结构Fig.3 Age structure of M. chapsensis population
2.4.1 乐昌含笑种群年龄结构 官山乐昌含笑种群3个样地中的个体总数为152株,其中最大胸径为61.70 cm,最高为37 m。从年龄结构图可看出官山乐昌含笑在各个龄段均有分布,种群结构大体呈“金字塔”型(图3)。随着龄级增加,乐昌含笑个体数大体呈下降趋势。其中第I龄级的个体数最多为74棵,其次是第Ⅱ龄级和第Ⅲ龄级,这前3龄级植株数量为131,占总数的86.2%,其他径级合计只有13.8%。第Ⅳ龄级到第Ⅵ龄级的个体数都较少,特别是在第Ⅴ龄级,种群个体数陡然下降,仅为1棵,占总数的1.0%。总的来说,胸径小于15 cm的个体数较多。

表5 江西官山乐昌含笑种群静态生命表Tab.5 Static life-table of M. chapsensis population
x为年龄级;Ax在x龄级的实际存活数;ax为匀滑后的x龄级的存活数;lx表示在x龄级开始时标准化存活数(以1 000为基数),lx=ax/ax-1×lx-1;dx表示从x到x+1龄级期间标准化死亡数,dx=lx-lx+1;qx表示从x到x+1龄级间隔期间的死亡率,qx=dx/lx;Lx表示从x到x+1龄级间隔期间平均存活的个体数,Lx=(lx+lx+1)/2;Tx表示从x龄级到超过x龄级的个体总数,Tx=Lx+Lx +1+Lx+2+…,即Tx∑Lx;ex表示进入x龄级个体的生命期望,ex=Tx/lx;Kx表示消失率(损失度),Kx=lnlx-lnlx+1
x:age;ax:the number of alive organisms at agex;lx:the number of survivors at the agex,supposing the number of population at the beginning is 1000;dx:the number of death individuals from agextox+1;qx:the proportion of death during the interval age (x,x+1);Lx:the number of individuals alive during unit time (x,x+1);Tx:total number of alive organisms from agex;ex:observed expectation of life at agex;Kx:killing power,Kx=lnlx-lnlx+1
2.4.2 种群静态生命表和存活曲线 从静态生命表(表5)可以看出,官山乐昌含笑种群随年龄增加,存活数lx逐渐减少;期望值ex在第Ⅱ龄级最大,为1.67,而在第Ⅵ龄级徒然降至最低,仅为0.79。从种群存活曲线看,乐昌含笑种群的Deevey存活曲线趋于第Ⅲ型曲线(“凹”形),小径级个体存活数较多,早期斜率较大,即小径级的个体在向中径级的发育过程中有极高的死亡率,但是一旦存活到第III径级以后,死亡率就降低并逐步稳定(图4)。种群的死亡率曲线和消失率曲线变化趋势一致,种群个体死亡率qx和消失率kx最大值都在第Ⅲ龄级(图5)。

图4 乐昌含笑种群存活曲线Fig.4 Survival curve of M. chapsensis population

图5 乐昌含笑种群死亡率及消失率曲线Fig.5 Mortality rate and hazard rate of M. chapsensis population
3 讨论与结论
3.1 群落复杂性与乐昌含笑更新
群落组成和结构是群落中各种生物在空间和时间上配置状况的表现,对研究群落原始生产力和环境条件资源都有重要的价值[32]。其中物种组成是群落结构和生物多样性的基础[33],与群落结构和生物多样性共同决定生态系统的功能。本研究中乐昌含笑3个样地共有1 085株121种,隶属72属43科,GS-MC1(400 m2)、GS-MC2(600 m2)和GS-MC3(2 500 m2)样地分别有85株16种14属12科、147株29种22属16科和853株115种66属42科。乐昌含笑重要值无论是在乔木层还是灌木层皆位于前列,在样地GS-MC1、GS-MC2和GS-MC3乔木层的重要值均最大,分别占31.05、27.96和15.94;在灌木层分别为14.91、11.44和9.59。前期的相关研究得出,湖南醴陵王村乐昌含笑群落(800 m2)有植物82种68属38科(含草本),乐昌含笑在乔木层中的重要值达159.55,而灌木层乐昌含笑重要值未排前列[4];广州南亚热带低丘平地的1 600 m2乐昌含笑样地内有维管束植物70种58属44科,其中乔木树种35种、灌木树种16种,乐昌含笑在乔木层的重要值为62.84,灌木层排名靠前的主要物种也未见乐昌含笑[11]。与以上研究相比,官山乐昌含笑群落在物种数量和重要值上的差异可能主要与面积、区域生物多样性水平以及与计算公式差异有关。马磊[11]和刘克旺[4]的研究通过相对多度、相对频度、相对显著度之和进行度量,而本文采用相对频度、相对密度和相对显著度、相对高度4者之和的平均值来度量。乐昌含笑群落结构分布图可以反映群落结构复杂,各级皆有物种分布。处在不同坡度与坡向的3个样地垂直结构与径级结构分布极为相似,皆表现为随高度级、径级增加,树木多度百分比逐渐下降,与杨萍等[25]对贵州黎平县乐昌含笑群落结构特征研究结果相似。群落中物种个体高度、径级大都分别处于0~10 m和0~5 cm,表明群落中、小径级个体较多,群落繁殖力强,更新良好。研究发现样地GS-MC3共优种麻栎仅乔木层有7株大径级个体,而灌木层并没有幼苗幼树分布。这可能是因为麻栎喜光,通常出现在演替前期,而在群落演替过程中,随物种数量增多,郁闭度增大,麻栎可能因光照不足,竞争能力弱于耐荫树[34],于是整个群落内没有其小径级个体,而成株的存在可能是群落演替过程中所遗留的树种,由此可推测该种可能面临自我更新困难。以上分析可以说明,本研究乐昌含笑群落物种组成丰富、结构复杂且乐昌含笑优势地位明显,是群落的优势种和建群种。
物种多样性既可以表明生物资源的丰富程度,又体现了生物与环境之间的复杂关系。多样性水平高低体现在2个方面:丰度和均匀度。物种丰富度和Margalef指数常用来衡量种类丰度,Pielou均匀度和生态优势度指数反映了物种之间多度分配的均匀程度,Simpson指数和Shannon-Wiener指数则是两者的综合,更好地度量群落物种多样性水平[35-36]。研究中,样地GS-MC1、GS-MC2和GS-MC3乔木层的物种数分别为7、15和59种,灌木层分别是14、19和92种;Shannon-Wiener指数在乔木层分别为1.69、2.37和3.32,灌木层为2.09、2.34和3.72;Simpson指数在5.41~27.54;Pielou均匀度指数范围在0.79~0.87,生态优势度指数仅为0.04~0.21,结果符合种与面积曲线变化规律[12,37]。与前期杨萍等[25]对贵州黎平县乐昌含笑研究相比,在面积(600 m2)相同条件下,官山样地GS-MC2乔木层Simpson多样性指数(9.50)、Pielou均匀度(0.87)均远高于黎平县乐昌含笑林(0.86和0.77);一般情况下,物种丰富,结构复杂而且物种之间分配较均匀的群落,其稳定性越大,生态服务功能和生产力水平越高[38],这表明官山乐昌含笑天然林群落稳定性较大,应对环境突变或外部干扰的能力较强。从该群落层次多样性格局来看,灌木层的物种多样性水平高于乔木层,可能是由于乔木层树种种类相对固定,同样发现灌木层除了灌木树种外,还包括了绝大多数乔木树种的幼苗、幼树,这都能造成灌木层的多样性指数高于乔木层[24]。
3.2 种群年龄结构与自然择优
从乐昌含笑年龄结构分布图可知,乐昌含笑种群结构呈“金字塔”型,种群属于增长型。乐昌含笑在各年龄段基本都有分布,随着龄级增加,种群数量逐渐下降,种群幼树、幼苗较多,结果与马磊等[11]对广州增城市乐昌含笑种群特征研究相反。造成这种龄级分布格局的原因可能是因为在小树阶段,官山乐昌含笑分布格局主要是由本身的生物生态学特性决定的,乐昌含笑的繁殖力强,在蔽荫环境中,乐昌含笑种子能正常发芽,幼苗较耐荫[25],在与其他树种的竞争过程中,经常处于优势的位置,并逐步扩展,加上受外界干扰较少,因此表现为幼树较多。随着种群的继续发育,密度制约因素引起种内和种间竞争加剧,增加种群个体的死亡[39],因而后期个体数逐渐减少。而种群存活曲线规律显示,小径级向中间径级发育过程中有极高的死亡率,直到第III径级以后,死亡率才逐渐降低而变得稳定。这说明虽然种群幼龄时期数量较多,有充足的数量为中间和大径级作为补充,但同样幼树有极高的死亡率,表明第I径级到第III径级这一时期种内或种间竞争激烈[40]。由于幼苗发育过程中过高的死亡率会对种群后续更新造成严重不良影响,为维持其自身在群落当中的竞争力和实现自身的良好更新,乐昌含笑种群在幼年向中老龄过度时期可能存在自然选优过程,使得在第III径级后,随着种群密度减小,个体逐渐到达主层林,竞争能力增强,生长空间加大,死亡率降低,种群进入相对稳定阶段[41]。期望值(ex)在第Ⅱ径级值达到种群的最大值为1.67,同样也说明小径级的活力很大,有大量小径级个体在短期内能够进行补充维持种群自我更新;大径级后期,期望值陡然下降到0.79,表明进入成熟期以后,乐昌含笑的种群活力会逐渐丧失。以上分析可以说明,乐昌含笑在组成复杂的群落背景中完全可以自然更新,且存在一个强烈的自然择优过程。
目前群落特征本底调查研究发现,官山国家级自然保护区乐昌含笑群落物种组成丰富,多样性水平较高,乐昌含笑种群自然更新良好,可对官山乐昌含笑天然林进行就地保护,这既是保护乐昌含笑种群的需要,也是保护其它种及复杂种间关系的必要。长期定位监测是探讨珍稀植物濒危机制的有效的途径,而以上的研究发现仅是对现阶段乐昌含笑生长状况的分析结果,因此,今后乐昌含笑种群动态变化、种间关系、群落演替方向等有待后期序列监测研究,从而为该种保护和可持续发展提供理论参考。
致谢:江西农业大学文仁权等同学及江西省宜春市林业科学研究所肖智勇工程师野外协助调查,官山国家级自然保护区和官山林业局对本试验给予了大力支持,谨致谢意。
参考文献:
[1] Mouillot D,Bellwood D R,Baraloto C,et al.Rare species support vulnerable functions in high-diversity ecosystems[J].Plos Biology,2013,11(5):e1001569.
[2] 方精云,王襄平,沈泽昊,等.植物群落清查的主要内容、方法和技术规范[J].生物多样性,2009,17(6):533-548.
Fang J Y,Wang R P,Sheng Z H,et al.Methods and protocols for plant community inventory[J].Biodiversity Science,2009,17(6):533-548.
[3] 祁承经,汤庚国.树木学[M].北京:中国林业出版社,2015:91-93.
Qi C J,Tang G G.Dendrology[M].Beijing:China Forestry Publishing House,2015:91-93.
[4] 刘克旺,苏勇,侯碧清.乐昌含笑群落特征及其种群动态[J].中南林学院学报,2004,21(4):47-50.
Liu K W,Su Y,Hou B Q.Research on the characteristics and dynamics ofMicheliachapensiscommunities[J].Journal of Centural South Forest University,2004,21(4):47-50.
[5] 李玉媛,李达孝.云南木兰科植物的保护价值与开发前景[J].北京林业大学学报,1999,21(3):29-35.
Li Y Y,Li D X.Conservation value and exploitation foreground of the magnoliaceae plants in Yunnan[J].Journal of Beijing Forestry University,1999,21(3):29-35.
[6] 邵文豪,姜景民,栾启福,等.乐昌含笑不同种源幼林生长性状变异和种源选择[J].江西农业大学学报,2011,33(4):701-706.
Shao W H,Jiang J M,Luan Q F,et al.A Study on variation of growth traits in youngMicheliachapensisforest and provenances selection[J].Acta Agriculturae Universitatis Jiangxiensis,2011,33(4):701-706.
[7] 姜景民,滕花景,袁金玲,等.乐昌含笑种群遗传多样性的研究[J].林业科学研究,2005,18(2):109-113.
Jiang J M,Teng H J,Yuan J L,et al.Genetic diversity ofMicheliachapensisDandy populations[J].Forest Research,2005,18(2):109-113.
[8] Cao X Y,Li C G,Miao Q,et al.Molecular cloning and expression analysis of a leaf-specific expressing 3-hydroxy-3 methylglutaryl-CoA (HMG-CoA) reductase gene fromMicheliachapensisDandy[J].Journal of Medicinal Plants Research,2011,5(16):3868-3875.
[9] 王润辉,张伟红,郑会全,等.乐昌含笑多点种源试验与优良种源选择[J].中南林业科技大学学报,2015,35(5):16-21.
Wang R H,Zhang W H,Zheng H Q,et al.Study on mutil-site provenance trials and superior provenances selection ofMicheliachapensis[J].Journal of Central South University of Forestry & Technology,2015,35(5):16-21.
[10] 姜景民.木兰科植物种质资源评价和乐昌含笑品种选育研究[D].北京:中国林业科学研究院,2006.
Jiang J M.Genetic resource evaluation of magnoliaceas species and cultivar breeding ofMicheliachapensisDandy[D].Beijing:Chinese Academy of Forestry Sciences,2006.
[11] 马磊,吴林芳,叶华谷,等.广州增城叶迳背乐昌含笑林群落特征研究[J].热带亚热带植物学报,2012,20(2):169-176.
Ma L,Wu L F,Ye H G,et al.Studies on community characteristics ofMicheliachapensisin Yejingbei,Zengcheng,Guangzhou[J].Journal of Tropical and Subtropical Botany,2012,20(2):169-176.
[12] 匡旭,邢丁亮,张昭臣,等.长白山北坡云冷杉林和落叶松林物种组成与群落结构[J].应用生态学报,2014,25(8):2149-2157.
Kuang X,Xing D L,Zhang S C,et al.Species composition and community structure of the spruce-fir forest and larch forest plots on the northern slope of Changbai Mountains,northeast China[J].Chinese Journal of Applied Ecology,2014,25(8):2149-2157.
[13] Anderson-Teixeira K J,Davies S J,Bennett A C,et al.CTFS-ForestGEO:a worldwide network monitoring forests in an era of global change[J].Global Change Biology,2015,21(2):528-549.
[14] Bongers F.Increasing liana abundance and biomass in tropical forests:emerging patterns and putative mechanisms[J].Ecology Letters,2011,14(14):397-406.
[15] 兰国玉.世界热带森林生态系统大样地定位研究进展[J].西北植物学报,2007,27(10):2140-2145.
Lan G Y.Research progress on large,long-term plot of tropical forest ecosystem in the world[J].Acta Bot Boreal,2007,27(10):2140-2145.
[16] 袁志良,王婷,朱学灵,等.宝天曼落叶阔叶林样地栓皮栎种群空间格局[J].生物多样性,2011,19(2):224-231.
Yuan Z L,Wang T,Zhu X L,et al.Patterns of spatial distribution ofQuercusvariabilisin deciduous broadleaf forests in Baotianman Nature Reserve[J].Biodiversity Science,2011,19(2):224-231.
[17] 杨庆松,马遵平,谢玉彬,等.浙江天童20 ha常绿阔叶林动态监测样地的群落特征[J].生物多样性,2011,19(2):215-223.
Yang Q S,Ma Z P,Xie Y B,et al.Community structure and species composition of an evergreen broadleaved forest in Tiantong’s 20 ha dynamic plot,Zhejiang province,eastern China[J].Biodiversity Science,2011,19(2):215-223.
[18] 刘信中,吴和平.江西官山自然保护区科学考察与研究[M].北京:中国林业出版社,2005.
Liu X Z,Wu H P.Scientific survey and study on the Guanshan nature reserve in Jiangxi province[M].Beijing:China Forestry Publishing House,2005.
[19] 曹岚,梁芳,邹红,等.江西官山国家级自然保护区珍稀植物多样性研究[J].安徽农业科学,2012,40(3):1696-1698.
Cao L,Liang F,Zou H,et al.Study on the diversity of rare and endangered plants in Guanshan Natural Reserve of Jiangxi Province[J].Journal of Anhui Agri Sci,2012,40(3):1696-1698.
[20] 姚振生,徐向荣,陈京,等.江西官山自然保护区药用植物资源[J].亚热带植物科学,2005,34(4):43-47.
Yao Z S,Xu X R,Chen J,et al.Medicinal plant resources of Guanshan Nature Reserve in Jiangxi Province[J].Subtropical Plant Science,2005,34(4):43-47.
[21] 闫淑君,洪伟,吴承祯,等.武夷山天然米槠林优势种群结构与分布格局[J].热带亚热带植物学报,2002,10(1):15-21.
Yan S J,Hong W,Wu C Z,et al.The structure and distribution pattern of dominant populations inCastanopsiscarlesiicommunity[J].Journal of Tropical and Subtropical Botany,2002,10(1):15-21.
[22] 丁晖,杨云芳,徐海根,等.武夷山典型常绿阔叶林动态监测样地:物种组成与群落结构[J].生态学报,2014,35(4):2-18.
Ding H,Yang Y F,Xu H G,et al.Species composition and community structure of the typicalevergreen broad-leaved forest in the Wuyi Mountains of southeastern China[J].Acta Ecologica Sinica,2014,35(4):2-18.
[23] 许涵,李意德,林明献,等.海南尖峰岭热带山地雨林60 ha动态监测样地群落结构特征[J].生物多样性,2015,23(2):192-201.
Xu H,Li Y D,Lin M X,et al.Community characteristics of 60 ha dynamics plot in the tropical montane rain forest in Jianfengling,Hainan island[J].Biodiversity Science,2015,23(2):192-201.
[24] 张志祥,刘鹏,刘春生,等.浙江九龙山南方铁杉(Tsugatchekiangensis)群落结构及优势种群更新类型[J].生态学报,2008,28(9):4547-4558.
Zhang Z X,Liu P,Liu C S,et al.The structure characteristics and dominant population regeneration types ofTsugatchekiangensiscommunities in the Jiulongshan national natural reserve of Zhejiang Province.[J].Actaec Ecologica Sinica,2008,28(9):4547-4558.
[25] 杨萍,吴宗慰,潘德权,等.贵州省黎平县楠竹林场乐昌含笑群落研究[J].种子,2012,31(12):54-58.
Yang P,Wu Z W,Pan D Q,et al.Studies onMichiliachapensiscommunity bamboo forest management area of Liping county in Guizhou[J].Seed,2012,31(12):54-58.
[26] 彭重华,巫涛,王海峰,等.乐昌含笑群落的分布格局分析[J].河南农业科学,2011,40(7):113-117.
Peng C H,Wu T,Wang H F,et al.The distribution pattern analyses ofMicheliachapensiscommunities[J].Journal of Henan Agricultural Sciences,2011,40(7):113-117.
[27] 谭一波,申文辉,郑威,等.广西容县红锥天然林物种组成与群落结构特征[J].中南林业科技大学学报,2015,35(5):98-102.
Tan Y B,Shen W H,Zhen W,et al.Species composition and community structure characteristics ofCastanopsishystrixnatural forest in Rongxian county,Guangxi Province[J].Journal of Central South University of Forestry & Technology,2015,35(5):98-102.
[28] 易雪梅,张悦,姬兰柱.长白山黄檗种群结构[J].生态学杂志,2013,32(9):2257-2262.
Yi X M,Zhang Y,Ji L Z.Population structure ofPhellodendronamurensein Changbai Mountains of northeast China[J].Chinese Journal of Ecology,2013,32(9):2257-2262.
[29] 李先琨,苏宗明,向悟生,等.濒危植物元宝山冷杉种群结构与分布格局[J].生态学报,2002,22(12):2246-2253.
Li X K,Su Z M,Xiang W S,et al.Study on the structure and spatial pattern of the endangered plant population ofAbiesyuanbaoshanensis[J].Actaec Ecologica Sinica,2002,22(12):2246-2253.
[30] 马克平.生物群落多样性的测度方法Ⅰα多样性的测度方法(上)[J].生物多样性,1994,2(3):162-168.
Ma K P.A method for biological communinity diversity(Ⅰ)[J].Chinese Biodiversity,1994,2(3):162-168.
[31] 马克平,刘玉明.生物群落多样性的测度方法Ⅰα多样性的测度方法(下)[J].生物多样性,1994,2(4):231-239.
Ma K P,Liu Y M.A method for biological communinity diversity(Ⅱ)[J].Chinese Biodiversity,1994,2(4):231-239.
[32] Miller R L,Fujii R.Plant community,primary productivity,and environmental conditions following wetland re-establishment in the Sacramento-San Joaquin Delta,California[J].Wetlands Ecology and Management,2010,18(1):1-16.
[33] 欧阳明,杨清培,陈昕,等.毛竹扩张对次生常绿阔叶林物种组成、结构与多样性的影响[J].生物多样性,2016,24(6):649-657.
Ouyang M,Yang Q P,Chen X,et al.Effects ofPhyllostachysedulisexpansion on species composition,structure and diversity of secondary evergreen broad-leaved forest[J] Chinese Biodiversity,2016,24(6):649-657.
[34] 徐飞,郭卫华,徐伟红,等.不同光环境对麻栎和刺槐幼苗生长和光合特征的影响[J].生态学报,2010,30(12):3098-3107.
Xu F,Guo W H,Xu W H,et al.Effects of light intensity on growth and photosynthesis of seedlings ofQuercusacutissimaandRobiniapseudoacacia[J].Acta Ecologica Sinica,2010,30(12):3098-3107.
[35] 李禄军,曾德慧.物种多样性与生态系统功能的关系研究进展[J].生态学杂志,2008,27(11):2010-2017.
Li L J,Zeng D H.Relationships between species diversity and ecosystem function:a review[J].Chinese Journal of Ecology,2008,27(11):2010-2017.
[36] 康永祥,康博文,刘建军,等.陕北黄土高原文冠果群落结构及物种多样性[J].生态学报,2010,30(16):4328-4339.
Kang Y X,Kang B W,Liu J J,et al.Structure and species diversity ofXanthocerassorbifoliacommunity in the Loess Plateau of North Shanxi[J].Actaec Ecologica Sinica,2010,30(16):4328-4339.
[37] 兰国玉,吴志祥,谢贵水.海南橡胶林植物多样性特征[J].生物多样性,2014,22(5):658-666.
Lan G Y,Wu Z X,Xie G S.Characteristics of plant species diversity of rubber plantation in Hainan island[J].Biodiversity Science,2014,22(5):658-666.
[38] 杨清培,钟安建,金志农,等.江西武夷山南方铁杉群落分类及更新能力评价[J].江西农业大学学报,2014,36(6):1275-1283.
Yang Q P,Zhong A J,Jin Z N,et al.A community classification of nature forest ofTsugatchekiangensisand its regeneration capacity[J].Acta Agriculturae Universitatis Jiangxiensis,2014,36(6):1275-1283.
[39] 列志旸,李洁,周彤彤,等.密度对乐昌含笑幼苗养分积累的影响[J].湖南林业科技,2016,43(1):44-47.
Lie Z C,Li J,Zhou T T,et al.Density effect on nutrient content and accumulation ofMicheliachapensisplantations[J].Hunan Forestry Science & Technology,2016,43(1):44-47.
[40] 吴显坤,南程慧,汤庚国,等.珍稀濒危植物浙江楠种群结构分析[J].安徽农业大学学报,2015,42(6):980-984.
Wu X K,Nan C H,Tang G G,et al.The population structure of the endangered plantPheobechekiangensis[J].Journal of Anhui Agricultural University,2015,42(6):980-984.
[41] 陈洁,周年兴,李灿,等.集合种群竞争机制在庐山森林演替模拟中的应用[J].生态学杂志,2017,36(3):862-868.
Chen J,Zhou N X,Li C,et al.Application of meta-population competition mechanism in forest succession simulation of Mount Lushan[J].Chinese Journal of Ecology,2017,36(3):862-868.
