基于DNA条形码对浙南岛屿日本花棘石鳖的遗传特征分析
2018-06-29吴晓雯张华伟张永普
吴晓雯, 张华伟, 余 海, 彭 欣, 张永普
(1. 温州大学生命与环境科学学院,浙江温州 325035;2. 浙江省海洋水产养殖研究所, 浙江温州 325005;3.苍南县水产研究所,浙江苍南 325802)
日本花棘石鳖(LiolophurajaponicaLischke)隶属于软体动物门(Mollusca),多板纲(Polyplacophora),石鳖科(Chitonidae),花棘石鳖属,栖息于潮间带中、低潮区,常附着于岩石上或石缝间。在中国东海、南海及朝鲜半岛、日本均有分布。该种环带肌肉肥厚,可食用,福建和海南等沿海居民常采捕食其肉,故有一定的经济价值[1]。日本花棘石鳖在浙江南部沿海的洞头列岛、北麂列岛、大北列岛、南麂列岛、七星列岛均为常见种[2-4]。多板纲软体动物的研究大多集中于形态特征[5- 6]、种类分布和群落结构[1,7-8]、生活习性[9]、分类[10-14]以及齿舌的物理学特征[15]等方面。近年来,通过分子手段对石鳖的系统分类和进化研究也有报道[16-17],但未涉及石鳖的种群结构和遗传多样性等。
线粒体DNA(mitochondrial DNA,mtDNA)具进化速率较快,母性遗传且基因重组率低等优点,是研究种群遗传结构和进化历史的理想分子标记[18-20]。mtDNA可以较好的揭示种内和种间的变化[21],其中细胞色素氧化酶亚基I(COⅠ),基因变异较大、进化速率较快,在软体动物腹足类[22]、双壳类[23]、头足类[24]等分子系统和种群遗传结构等领域有广泛的应用。16S rRNA基因在一级结构和二级结构上都相对保守,进化速率比COⅠ基因慢,在种群研究中也有一定的应用[25]。
本研究以浙江南部沿海的洞头列岛(洞头岛、大竹峙岛、东策岛、南爿山岛、鹿西岛、大瞿岛),北麂列岛(北麂岛),大北列岛(北龙山),南麂列岛(南麂岛、后麂山岛、大檑山屿、柴峙岛)和七星列岛(东星仔岛)岩相潮间带的13个日本花棘石鳖自然群体为研究对象,构建系统进化树,旨在探讨该种的种群结构、遗传多样性、遗传背景等特征,推测不同自然群体的系统进化关系,为其资源保护和遗传学研究提供理论依据。
1 材料与方法
1.1 样品采集
本研究所用的样品于2017年采自浙南沿海的洞头列岛(洞头岛、大竹峙岛、东策岛、南爿山岛、鹿西岛、大瞿岛),北麂列岛(北麂岛),大北列岛(北龙山),南麂列岛(南麂岛、后麂山岛、大檑山屿、柴峙岛)和七星列岛(东星仔岛)的岩相潮间带(图1,表1),采集个体足部肌肉并标记分装后,置于液氮中带回实验室后-80 ℃保存。

图1 日本花棘石鳖采样点分布图Fig.1 Map of sampling sites of L. japonica

区域Region群体Population缩写Abbreviation东经East longitude北纬North latitude样本数Sample size洞头列岛洞头岛DT121°10'19.13″27°49'57.44″13 大竹峙岛 DZS121°12'48.74″27°49'12.87″13 东策岛DC121°09'01.38″27°45'32.04″13 南爿山岛 NPS121°15'33.62″28°00'15.29″10 大瞿岛DQ121°05'23.97″27°47'29.92″10 鹿西岛LX121°12'50.70″27°59'33.43″5北麂列岛北麂岛BJ121°11'47.82″27°37'08.82″13大北列岛北龙山BL120°58'51.56″27°40'08.56″12南麂列岛南麂岛NJ121°03'06.28″27°27'30.57″3 后麂山岛 HJS121°07'40.71″27°28'26.96″6 大檑山屿 DLS121°05'24.50″27°29'39.48″8 柴峙岛CS121°04'54.45″27°25'40.36″8七星列岛东星仔岛DX120°02'47.98″27°02'40.36″11
1.2 DNA提取
取足部肌肉,用双蒸水洗去杂物,再用滤纸吸干,按照全式金EasyPure Marine Animal Genomic DNA Kit试剂盒(北京全式金生物技术有限公司)步骤提取总DNA,于-20 ℃分管保存。
1.3 PCR扩增和测序
实验使用无脊椎动物COⅠ通用引物[26]扩增:LCOⅠ1490(5′-GGTCAACAAATCATAAAGATATTGG-3′)和 HCOⅠ 2198(5′-TAAACTTCAGGGTGACCAAAAAATCA-3′),16SA(5′-CGCCTGTTTATCAAAAACAT-3′)和16SB(5′-CCGGTCTGAACTCAGATCATGT-3′)[27],引物由上海生工(Sangon biotech)生物有限公司合成。
PCR反应为25 μL体系,其中包括:超纯水9.5 μL,上下游引物各1 μL,DNA模板1 μL, 2×EasyTaq PCR SuperMix(+dye)(北京全式金生物技术有限公司)12.5 μL。PCR程序设置:94 ℃预变性4 min,94 ℃变性30 s,51~53 ℃退火30 s,72 ℃延伸1 min(变性延伸35个循环),最后72 ℃再延伸10 min。每次反应均设置阴性对照以检验是否有污染。PCR产物经质量分数为1%的琼脂糖凝胶电泳检测,确定PCR产物为目的条带后,送至上海生工生物有限公司进行纯化及双向测序,以确保序列可靠性。
1.4 序列比对与分析
所得序列用DNAstar 5.0软件包中的Seqman软件处理,结合峰图进行人工校对,确保每一个位点准确。将两组数据(COⅠ和16S rRNA)通过NCBI的Blast比对分析,验证所得的序列均为目的基因。利用MEGA 6.0[28]中无脊椎动物线粒体遗传密码子把所有的COⅠ编码基因序列翻译成氨基酸,以确认此蛋白编码基因片段是否正常翻译。在所有序列中未发现终止密码子,可确认获得的序列是线粒体而不是核假基因拷贝[29]。
利用DNAsp 5.0[30]分析多态位点数(Singleton polymorphic sites),单倍型多样性(Haplotype diversity,Hd)和核苷酸多样性(Nucleotide diversity,Pi),统计单倍型。利用MEGA 6.0分析碱基组成,计算保守位点(Conserved sites)、变异位点数(Variable sites)、简约信息位点数(Parsim-information sites)、单突变位点(Singleton sites),根据Kimura两参数模型(Kimura-2-Parameter,K2P)[31]计算群体间的遗传距离以及单倍型遗传距离。采用最大拟然法(Maximum Likehood,ML法)[28]和贝叶斯法(Bayesian Inference,BI法)[32]构建系统发生树,进行1 000次自检。此外,利用Network 5.0[33]中Median-network法构建单倍型网络图,来探讨日本花棘石鳖的单倍型谱系关系。
2 结果与分析
2.1 碱基组成
本研究分析的125条日本花棘石鳖COⅠ基因片段经比对最终获得的长度为675 bp。由表2可知,A+T(62.5%)含量明显大于C+G(37.5%)含量,存在明显的A或T碱基偏向,符合线粒体组成特征。共有保守位点634个,变异位点41个,其中包括简约信息点22个,单突变位点19个。
125条日本花棘石鳖16S rRNA基因片段经比对长度为517 bp,由表2可知,A+T (64.9%)含量明显大于C+G(35.1%)含量,符合线粒体组成特征。共有保守位点508个,变异位点9个,其中简约信息点4个,单突变位点5个。
2.2 序列多态性分析
13个群体总体的COⅠ基因片段多态位点有41个,核苷酸多样度指数(Pi)为0.006 01±0.000 57,单倍型多样度指数(Hd)为0.847±0.025,平均核苷酸差异度为4.057 8。由表3可知,各群体间单倍型多样度在0.756(南爿山岛)~1.000(南麂岛、柴峙岛)之间,核苷酸多样度在0.003 0(大檑山屿)~0.014 8(南麂岛)之间,平均核苷酸差异数在2.036(大檑山屿)~10.000(南麂岛)之间。南麂岛群体的核苷酸多样度、单倍型多样度相对较高,而大檑山屿和南麂岛的核苷酸多样度、单倍型多样度都相对较低。
注:DT-洞头岛、DZS-大竹峙岛、DC-东策岛、NPS-南爿山岛、DQ-大瞿岛、LX-鹿西岛、 BJ-北麂岛、NJ-南麂岛、HJS-后麂山岛、BL-北龙山、DLS-大檑山屿、CS-柴峙岛、DX-东星仔岛。以下类同
Note: DT-Dongtou Island, DZS-Dazhushi Island, DC-Dongce Island, NPS-Nanpanshan Island, DQ-Daqu Island, LX-Luxi Island, BJ-Beiji Island, NJ-Nanji Island, HJS-Houjishan Island, BL-Beilongshan Island, DLS-Daleishan Island, CS-Chaishi Island, DX-Dongxizai Island. The same as following
13个群体总体的16S rRNA基因片段多态位点有9个,核苷酸多样度指数(Pi)为0.001 69 ± 0.003 24,单倍型多样度为指数(Hd)为0.665±0.029,平均核苷酸差异度为0.871 2。由表4可知,各群体间单倍型多样度在0.536(大檑山屿)~0.987(南麂岛)之间,核苷酸多样度在0.001 0(大檑山屿)~0.002 9(后麂山岛),平均核苷酸差异数在0.536(大檑山屿)~1.533(后麂山岛)之间。后麂山岛群体的核苷酸多样度最高,大檑山屿群体的核苷酸多样度最低;南麂岛群体的单倍型多样度最高,大檑山屿群体单倍型多样度最低。
此外,COⅠ基因片段共检测到41个变异位点,定义了30个单倍型(Hap1~30)(表5),其中,21个单倍型仅在单个群体一个个体中检测到,Hap17在东策岛群体的两个个体中检测到;Hap6是主体单倍型,基因序列表达为Hap6的个体占总个体数的34.1%,Hap5和Hap1各占总个体数的12.0%和11.2%。说明各群体内具有丰富的多态性,但单倍型共享率不高。
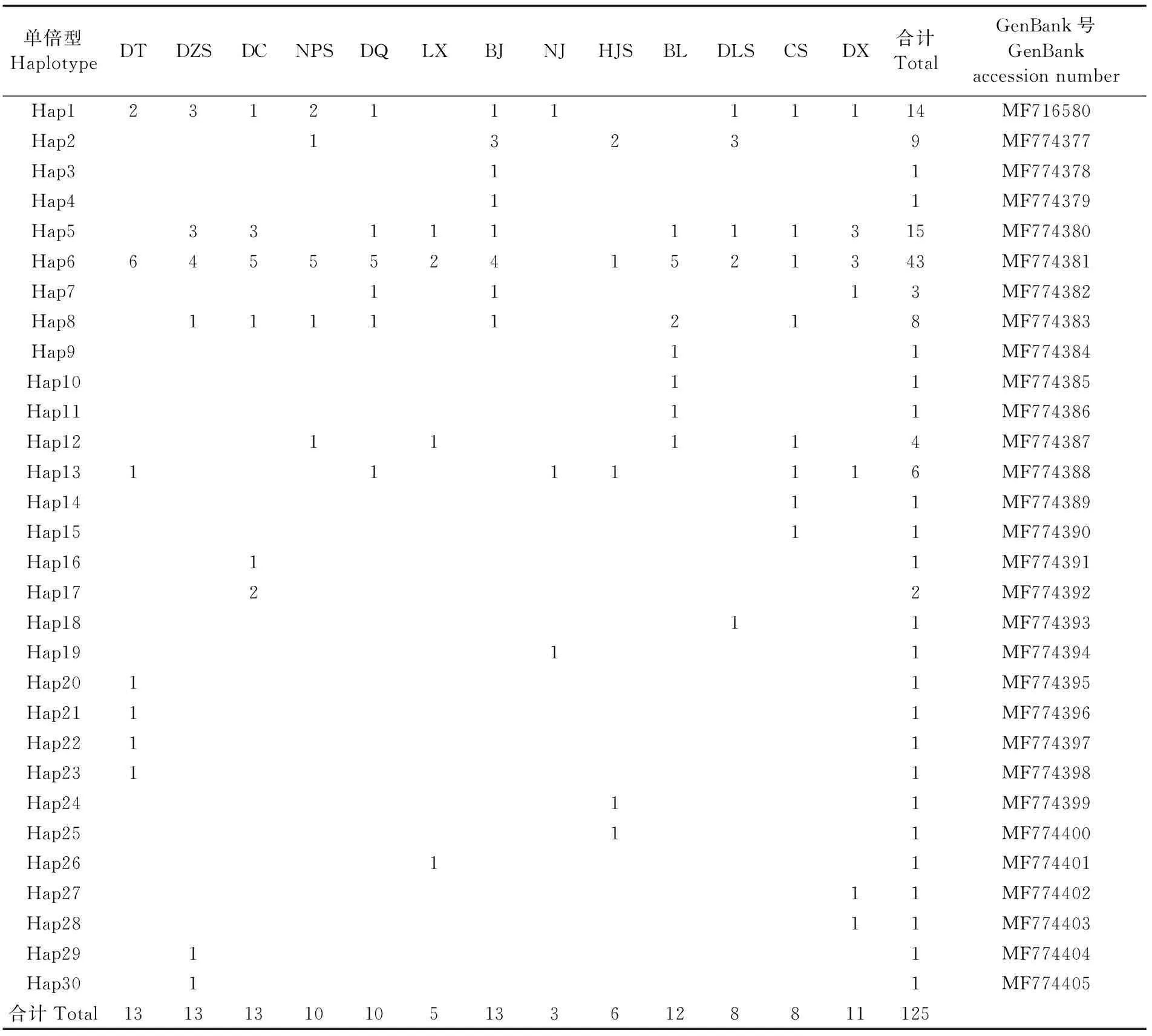
表5 各群体间 COⅠ基因片段单倍型分布情况Tab.5 Distribution of haplotypes of COⅠ gene fragments among 13 populations of L. japonica
16S rRNA基因片段共检测到9个变异位点,定义了10个单倍型(表6),其中,Hap1为所有群体共有,基因序列表达为Hap1的个体占总个体数的48%。Hap2占总个体数的31.2%,Hap3占总个体数的9.6%。Hap4、Hap5、Hap9、Hap10为单个群体所拥有。
2.3 遗传距离分析
13个群体间基于COⅠ基因片段的群体内遗传距离见表3,基于16S rRNA基因片段的遗传距离见表4;13个群体间的遗传距离见表7。基于COⅠ基因片段分析,13个日本花棘石鳖群体间遗传距离在0.003 1(大檑山屿-北麂岛,大檑山屿-南爿山岛)~0.011 0(大檑山屿-柴峙岛),群体内遗传距离在0.003 4(大檑山屿)~0.015 0(南麂岛);基于16S rRNA基因片段分析,群体间遗传距离在0.001 1(南麂岛-柴峙岛)~0.002 5(后麂山岛-柴峙岛),群体内遗传距离在0.001 0(大檑山屿)~0.003 0(后麂山岛)。日本花棘石鳖不同群体内和群体间的遗传距离处于同一水平,说明群体间遗传分化程度低。
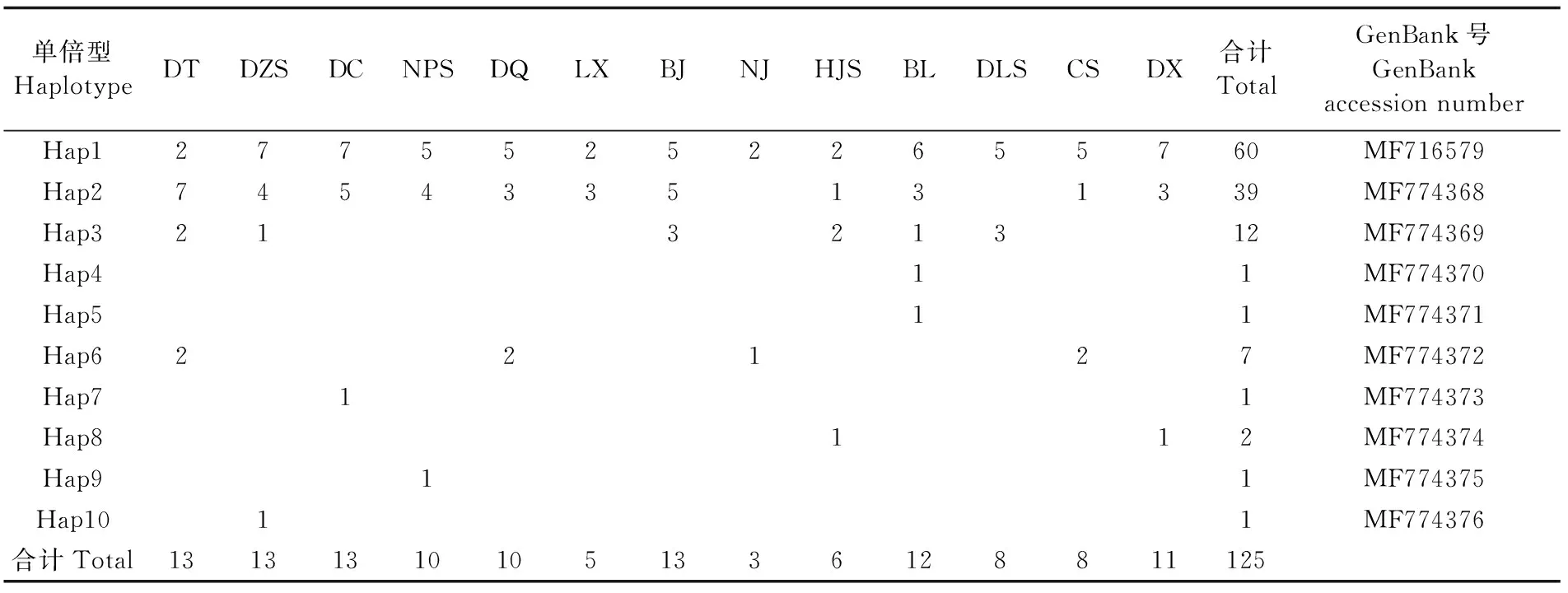
表6 各群体间16S rRNA基因片段单倍型分布情况Tab.6 Distribution of haplotypes of 16S rRNA gene fragment among 13 populations of L. japonica

表7 基于 COⅠ和16S rRNA片段的13个种群间遗传距离Tab.7 Genetic distances of COⅠ gene and 16S rRNA fragments among 13 populations of L. japonica
注:对角线下方为COⅠ遗传距离,对角线上方为16S rRNA遗传距离
Note: Genetic distances ofCOⅠ genes are below the diagonal, genetic distances of 16S rRNA are above the diagonal
2.4 系统发生分析
以Acanthopleurabrevispinosa(COⅠ:KX537628.1;16S: KX537613.1)和A.granulata(COⅠ: AY377719.1;16S: AY377608.1)为外群,以K-2-P为模型构建单倍型ML树,使用Modeltest 3.7选出最优模型构建单倍型 BI树。基于COⅠ基因的30个单倍型构建的ML树拓扑结构(图2)和BI树(图3)相吻合,日本花棘石鳖群体内Hap13、Hap15和Hap20聚合在一起,其它单倍型先后聚合一支,主要存在两个明显的单倍型类群,最后和外群类聚一起;基于16S rRNA基因的10个单倍型构建的ML树拓扑结构(图4)与BI树(图5)相吻合显示,10个单倍型先类聚为一支,再与外群聚合。部分单倍型置信值较低(置信值低于50%),且13个不同地理群体单倍型散乱分布,没有出现明显的地理结构和遗传谱系结构。
利用中介网络邻接法构建单倍型网络图,COⅠ基因网络图如图6,显示4个单倍型谱系分支,与ML系统树的拓扑结构相吻合。主要的单倍型为Hap1,Hap2和Hap6。其它单倍型通过一步或者多步突变分别于主要的相连。其中,Hap1突变为Hap5和Hap24,出现一个过渡单倍型,但在此研究中未检测到,可能与样本数量有关。16S rRNA基因片段的网络邻接法构建单倍型网络图(图7),比较单一。

图2 基于COⅠ基因13个群体日本花棘石鳖30个单倍型的ML系统发育树Fig.2 ML phylogenetic trees based on COⅠ gene fragments among 13 populations of L. japonica

图3 基于COⅠ基因13个群体日本花棘石鳖30个单倍型的BI系统发育树Fig.3 BI phylogenetic trees based on COⅠ gene fragments among 13 populations of L.japonica
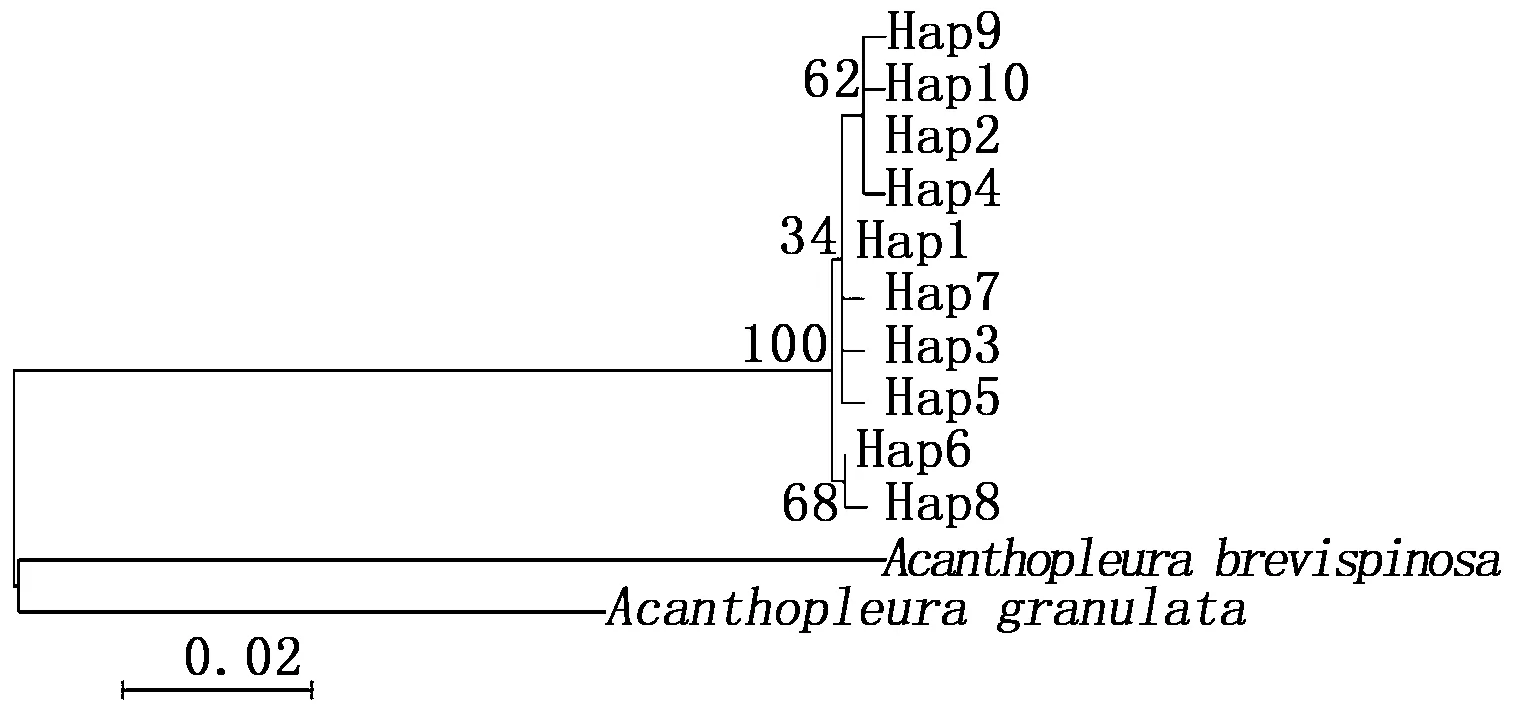
图4 基于16S rRNA基因13个群体日本花棘石鳖单倍型的ML系统发育树Fig.4 ML phylogenetic trees based on 16S rRNA gene fragments among 13 populations of L. japonica
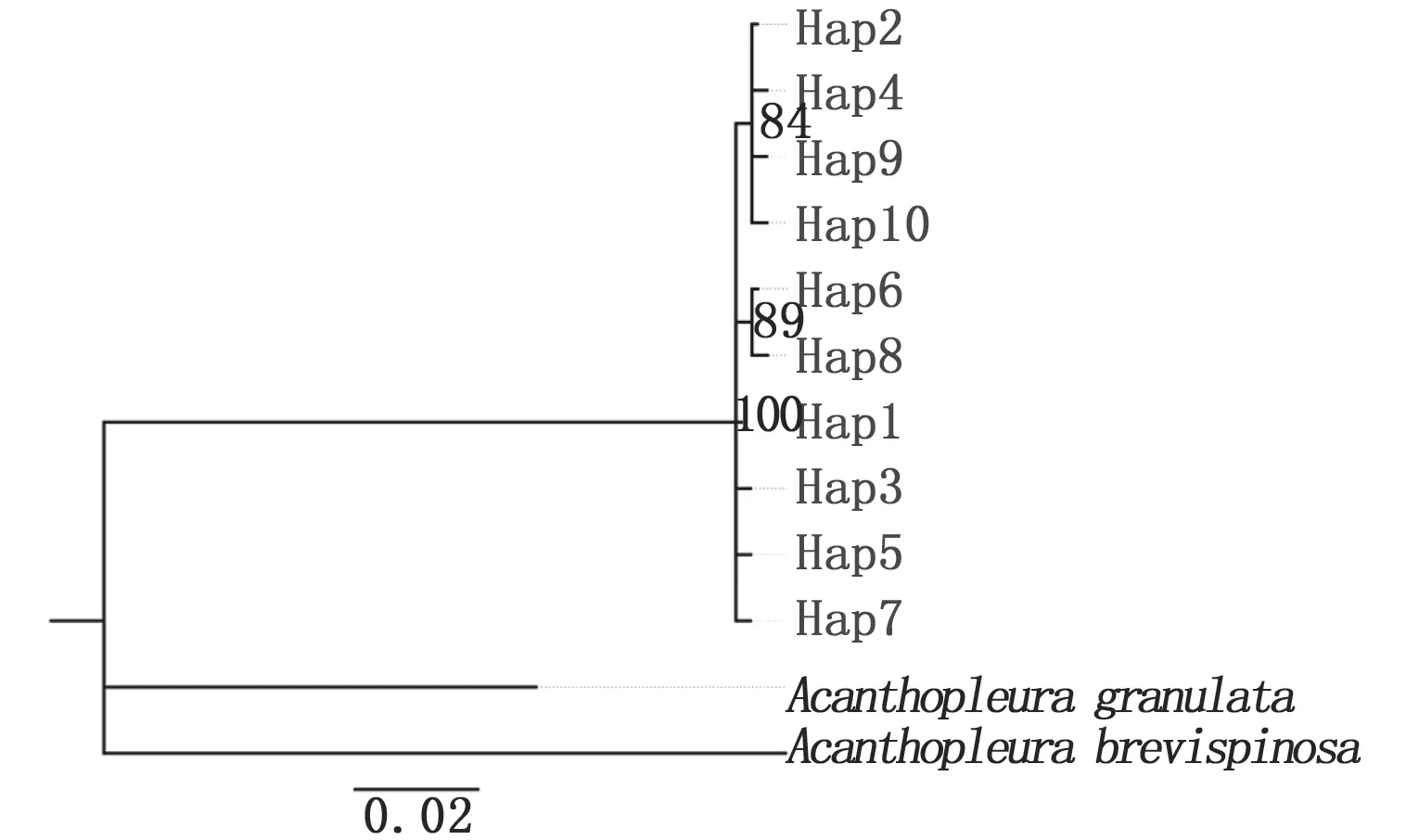
图5 基于16S rRNA基因13个群体日本花棘石鳖单倍型的BI系统发育树Fig.5 BI phylogenetic trees based on 16S rRNA gene fragments among 13 populations of L. japonica

图6 基于日本花棘石鳖COⅠ基因片段的Median-network网络图Fig.6 Median-network showing phylogenetic relationships among haplotypes based on COⅠ gene of L. japonica注:黄色圆圈表示某一具体单倍型,红色圆圈表示缺失单倍型;红色数字反应两两单倍型之间的变异位点Note: Yellow circle represents unique haplotypes, red circle represents a deletion of haplotypes, and the red number represents mutation steps

图7 基于日本花棘石鳖16S rRNA基因片段的Median-network网络图Fig.7 Median-network showing phylogenetic relationships among haplotypes based on 16S rRNA gene of L. japonica注:黄色表示某一具体单倍型,红色数字反应两两单倍型之间的变异位点Note: Yellow circle represents unique haplotypes, and the red number represents mutation steps
3 讨论
FOLMER等[26]认为,线粒体基因中A+T含量较高占有优势。无论是基于COⅠ基因还是16S rRNA基因,13个群体的日本花棘石鳖碱基组成显示了较高的A+T含量,与中国沿海16种帘蛤目的研究结果一致[34]。
COⅠ基因是线粒体氧化呼吸链重要的组成成分,基因变异较大;16S rRNA基因是非编码蛋白基因,不受密码子编码的选择压力,进化速度较为适中[35]。日本花棘石鳖COⅠ基因序列变异程度高于16S rRNA,表明16S rRNA基因比COⅠ基因更保守,COⅠ基因更适合用于种群内遗传研究。这一现象与双壳类的贻贝属Mytilidae[36]、黑龙江河蓝蛤(Potamocorbulaamurensis)[37],腹足类的的方斑东风螺(Babyloniaareolata)和台湾东风螺(Babyloniaformosae)[38]、头足类乌贼目(Sepioidea)[39]研究类似。
生物体进化的潜力和抵抗恶劣环境的生存能力,取决于种群内遗传变异的范围和种群遗传结构的分化程度[40]。遗传多样性常用核苷酸多样性和单倍型多样性等参数来衡量[41]。基于COⅠ基因片段分析,日本花棘石鳖整体核苷酸多样度(Pi)为0.006 01,单倍型多样度(Hd)为0.847,略高于双壳类的文蛤(Meretrixmeretrix)(Pi=0.002 39,Hd= 0.763)[42]和头足类的曼氏无针乌贼(Sepiellajaponica)(Pi=0.002 72,Hd=0.576)[24]。物种对环境变化的适应能力与遗传多样性呈正相关关系,适应能力越强,物种分布范围越广[43]。日本花棘石鳖属于广盐广布种[1],需要有足够的变异来适应复杂的环境。13个群体的核苷酸多样度和单倍型多样度分别在0.003 0 ~ 0.014 8和0.756~1.000之间,其中南麂岛群体的核苷酸多样度和平均核苷酸差异最高,表明南麂岛群体的遗传背景和遗传多样性较丰富;基于16S rRNA基因片段分析,日本花棘石鳖整体核苷酸多样度为0.001 69,单倍型多样度为0.665,13个群体的核苷酸多样度和单倍型多样度分别在0.001 0~0.002 9和0.536~0.987之间,其中后麂山群体的核苷酸多态度和平均核苷酸差异高于其它群体,表明南麂岛群体较其它群体有丰富的遗传多样性。南麂岛和后麂山岛同属于南麂列岛,地处台湾暖流与江浙沿岸流交汇和交替消涨的海区,属亚热带海洋季风气候,浪击度大,盐度高,气候环境较其它海岛复杂多变,导致该地区日本花棘石鳖遗传多样性较高。
浙江南部沿海的日本花棘石鳖遗传特征总体呈现出单倍型多态度高,核苷酸多态度低的特点,这与李玉龙等[44]对辽东湾和韩国3个不同海蜇地理群体研究结果相似,推测日本花棘石鳖可能经历了种群扩张[45-46]。日本花棘石鳖单倍型多样性可能在短时间内积累变异而快速提高,而核苷酸多态性在短时间内尚未达到这种程度,从而出现高单倍型多样性,低核苷酸多态度的现象[47]。不同地理群体单倍型散乱分布,没有出现明显的地理结构和遗传谱系结构,表明各岛屿的日本花棘石鳖为近期分化并存在基因交流。大多数的海洋生物群体在广泛的空间内存在广泛的基因流。虽然此研究中的13个岛屿呈现隔离状态,但是海洋环境影响群体基因交流的阻碍较小,而且洋流可以作为日本花棘石鳖卵、浮游期幼体运动的媒介,从而加强不同地理群体之间的基因交流。多板纲软体动物是软体动物中较为原始的一个门类,在进化上外形和生活习性上相对保守[48-49];另外,采样的地理尺度跨越较小,这些可能是导致日本花棘石鳖遗传差异出现较小积累的原因。综上所述,浙南不同岛屿分布的日本花棘石鳖遗传变异较小,尚未达到种间分化。
致谢:感谢浙江师范大学张加勇博士在数据处理过程中的支持和帮助。
参考文献:
[1] 张永普, 应雪萍, 黄象栋, 等. 浙南岛屿岩相潮间带石鳖的种类组成与数量分布 [J]. 动物学杂志, 2001, 36 (3): 5-9.
ZHANG Y P, YING X P, HUANG X D,etal. The species composition and quantity distribution of the chitons in the rocky intertidal zone of the islands in the south Zhejiang [J]. Chinese Journal of Zoology, 2001, 36 (3): 5-9.
[2] 卢 周, 陆京明, 周化斌, 等. 洞头南北爿山海洋特别保护区潮间带大型底栖生物群落的时空分布 [J]. 温州大学学报 (自然科学版), 2016, 37 (3): 38-48.
LU Z, LU J M, ZHOU H B,etal. Spatial and temporal distribution of macro-benthos communities in the intertidal zone of special marine protection area in Nanbeipan mountain, Dongtou district [J]. Journal of Wenzhou University (Natural Sciences), 2016, 37 (3): 38-48.
[3] 彭茂潇, 钱培力, 张永普, 等. 洞头无居民海岛岩相潮间带夏季大型底栖动物群落格局[J]. 生态学杂志, 2013, 32 (9): 2469-2479.
PENG M X, QIAN P L, ZHANG Y P,etal. Community structure of macrozoobenthos in rocky intertidal zone of uninhabited islands in Dongtou County, Zhejiang Province of East China in summer [J]. Chinese Journal of Ecology, 2013, 32 (9): 2469-2479.
[4] 余 海, 阙伟伟, 陆京明, 等. 七星列岛软体动物种类组成与区系 [J]. 温州大学学报 (自然科学版), 2016, 37 (4): 33-39.
YU H, QUE W W, LU J M,etal. Species composition and fauna of mollusca in Seven Star Islands [J]. Journal of Wenzhou University (Natural Sciences), 2016, 37 (4): 33-39.
[5] 张均龙, 史振平, 王 承, 等. 基于壳板和齿舌形态对中国沿岸几种常见多板纲软体动物的分类研究 [J]. 海洋科学, 2015, 39 (11): 96-107.
ZHANG J L, SHI Z P, WANG C,etal. Taxonomic study on some common species of Polyplacophora in the coast of China based on valves and rudular morphology [J]. Marine Sciences, 2015, 39 (11): 96-107.
[6] 陈道海, 孙世春. 9种石鳖壳板的形态研究 [J]. 中国海洋大学学报 (自然科学版), 2010, 40 (6): 53-60.
CHEN D H, SUN S C. Observations on the shell plates of 9 chiton species (Polyplacophora) [J]. Periodical of Ocean University of China (Natural Sciences), 2010, 40 (6): 53-60.
[7] 张永普, 刘德庆. 浙南岛屿岩相潮间带石鳖的群落结构 [J]. 动物学杂志, 2002, 37 (4): 5-9.
ZHANG Y P, LIU D Q. Community structure of chitons in the rocky intertidal zone of the islands in South Zhejiang [J]. Chinese Journal of Zoology, 2002, 37 (4): 5-9.
[8] SIRENKO B I. Report on present state of our knowledge with regard to the chitons of the Magellan Strait and Falkland Islands [J]. Venus, 2006, 65 (1-2): 81-89.
[9] 赵青松, 金 珊, 陈寅儿. 石鳖的结构与习性 [J]. 水利渔业, 2007, 27 (3): 46-47.
ZHAO Q S, JIN S, CHEN Y E. The structure and habits of chitons [J]. Reservoir Fisheries, 2007, 27 (3): 46-47.
[10] GÜLLER M, LIUZZI M G, ZELAYA D G. A new species ofLeptochiton(Polyplacophora: Leptochitonidae) from the southwestern Atlantic [J]. Malacologia, 2015, 58 (1-2): 147-155.
[11] OWADA M. A new cryptic species distinguished fromIschnochitoncomptus(Gould, 1859) (Polyplacophora: Ischnochitonidae) in central Honshu, Japan [J]. Molluscan Research, 2016, 36 (4): 255-263.
[12] 张均龙, 徐凤山, 张素萍. 多板纲软体动物系统分类学研究进展 [J]. 海洋科学, 2013, 37 (4): 111-117.
ZHANG J L, XU F S, ZHANG S P. Progress of the systematics of Polyplacophora (Mollusca) [J]. Marine Sciences, 2013, 37 (4): 111-117.
[13] CHEN D H, SUN S C. Analysis of diversity ofCOⅠ gene and system generation of nine species of chitons [J]. Wuhan University Journal of Natural Sciences, 2013, 18 (2): 171-177.
[14] BORIS S O,JAVIER S.Update of the genusLeptochiton(Mollusca: Polyplacophora) in Chilean deep waters three new reports and description of two newspecies [J]. Zootaxa, 2016, 4173 (3): 259-279.
[15] 钱 霞, 刘 维, 赵见高. 红条毛肤石鳖齿舌牙齿内的纳米磁性矿物质 [J]. 科学通报, 2002, 47 (1): 10-13.
QIAN X, ZHAO W, ZHAO J G. Magnetic anisotropy study of the radula of chitonAcanthochtonrubrolinestusLischke [J]. Chinese Science Bulletin, 2002, 47 (1): 10-13.
[16] IRISARRI I, EERNISSE D J, ZARDOYA R. Molecular phylogeny of Acanthochitonina (Mollusca: Polyplacophora: Chitonida): Three new mitochondrial genomes, rearranged gene orders and systematics [J]. Journal of Natural History, 2014, 48 (45-48): 2825-2853.
[18] BONATTO S L, SALZANO F M. A simple and early migration for control region sequences in a marine mammal: Implications for conservation and biogeography of Steller sea lions (Eumetopiasjubatus) [J]. Journal of Mammalogy, 1997, 77 (1): 95-108.
[19] STANLEYET H F, CASEY S, CARNAHAM J M,etal. Worldwide patterns of mitochondrial DNA differentiation in the harbor seal Phocavitulina [J]. Molecular Biochemical Evolution, 1996, 13 (2): 368-382.
[20] VILA C, SAVOLAINEN P, MALDONADO J E,etal. Multiple and ancient origins of the domestic dog [J]. Science, 1997, 276 (5319): 1687-1689.
[21] AVISE J C. Molecular markers: Natural history and evolution [M]. New York: Chapman & Hall, 1993.
[22] KUMAR R, JAISWAR A K, PAVAN-KUMAR. A. A ,etal. DNA barcoding ofThaisspecies (Family: Muricidae) from west coast of India [J]. Indian Journal of Fisheries, 2015, 62 (2): 92-97.
[23] 田吉腾, 侯 丫, 刘志鸿, 等. 基于线粒体COⅠ基因的毛蚶群体遗传多样性 [J]. 海洋科学, 2016, 40 (1): 1- 9.
TIAN J T, HOU Y, LIU Z H,etal. Genetic diversity of different populations of Scapharca subcrenata based on mitochondrialCOⅠ gene [J]. Marine Sciences, 2016, 40 (1): 1-9.
[24] LI J J, YE Y Y, WU C W,etal. Genetic diversity and population structure ofSepiellajaponica(Mollusca: Cephalopoda: Decapoda) inferred by mitochondrial DNA (COⅠ) variations [J]. Biochemical Systematics and Ecology, 2014(56): 8-15.
[25] QI P Z, GUO B Y, XIE C X,etal. Assessing the genetic diversity and population structure ofCulteralburnusin China based on mitochondrial 16S rRNA andCOⅠ gene sequences [J]. Biochemical Systematics and Ecology, 2013(50): 390-396.
[26] FOLMER O, BLACK M, HOEH W,etal. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoaninvertebrates [J]. Molecular Marine Biology and Biotechnology, 1994, 3 (5): 294-299.
[27] PALUMBI S R, MARTIN A P, ROMANO S,etal. The simple fool's guide to PCR version2.0 [M]. Honolulu: Department of Zoology, University of Hawaii, 1991.
[28] TAMURA K, STECHER G, PETERSON D,etal. MEGA6: Molecular evolutionary genetics analysis version 6.0 [J]. Molecular Biochemical Evolution. 2013, 30 (12): 2725 -2729.
[29] 韦筱媚, 刘 凡, 颜琳妙, 等. 基于线粒体基因的广西变色树蜥种群分子系统关系与遗传多样性 [J]. 四川动物, 2017, 36 (5): 519-530.
WEI X M, LIU F, YAN L M,etal. Phylogenetic relationship and genetic diversity ofCalotesversicolorin Guangxi [J]. Sichuan Journal of Zoology, 2017, 36 (5): 519-530.
[30] LIBRADO P, ROZAS J. DNASP v5: A software for comprehensive analysis of DNA polymorphism data [J]. Bioinformatics, 2009, 25 (11): 1451-1452.
[31] KIMURA M.A simple method for estimating evolutionary rate of base substitutions through comparative of studies of nucleotide sequence [J]. Journal of Molecular Evolution, 1980(16): 111-120.
[32] HUELSENBECK J P, RONDQUIST F. MRBAYES: Bayesian inference of phylogenetic trees [J].Bioinformatics Applications Note, 2001, 17 (8): 754-755.
[33] FORSTER P,TORRONI A,RENFREW C. Phylogenetic star contraction applied to Asian and Papuan mtDNA evolution [J]. Molecular Biochemical Evolution, 2001, 18 (10): 1864-1881.
[34] 王琳楠, 闫喜武, 秦艳杰, 等. 中国帘蛤目16 种经济贝类DNA 条形码及分子系统发育的研究 [J]. 大连海洋大学学报, 2013, 28 (5): 431-437.
WANG L N, YAN X W, QIN Y J,etal. DNA barcoding and molecular phylogeny of 16 species of economical shellfish in Veneroida along China Coast [J]. Journal of Dalian Ocean University, 2013, 28 (5): 431-437.
[35] 王 超, 曹 奕, 吴杨平, 等. 6个不同海域文蛤地理群体的亲缘关系分析 [J]. 海洋渔业, 2016, 38 (3): 262-272.
WANG C, CAO Y, WU Y P,etal. Genetic relationship analysis of sixMeretrixmeretrixpopulations from different sea areas [J]. Marine Fisheries, 2016, 38 (3): 262-272.
[36] 毛阳丽, 蔡厚才, 李成久, 等. 基于线粒体COⅠ与16S rRNA基因序列探讨贻贝属的系统发育 [J]. 南方水产, 2010, 6 (5): 27-36.
MAO Y L, CAI H C, LI C J,etal. Molecular phylogenetics of genusMytilusbased onCOⅠ and 16S rRNA sequences [J]. South China Fisheries Science, 2010, 6 (5): 27-36.
[37] 孙 超, 刘志鸿, 杨爱国, 等. 基于线粒体COⅠ和16S rRNA基因研究3个地理群体黑龙江河蓝蛤的遗传多样性 [J]. 海洋科学, 2015, 39 (1): 39-45.
SUN C, LIU Z H, YANG A G,etal. The genetic diversity of three populations ofPotamocorbulaamurensisbased on mitochondrialCOⅠ and 16S rRNA gene [J]. Marine Sciences, 2015,39(1): 39-45.
[38] 苏天凤, 黄建华, 吴进锋, 等. 两种东风螺线粒体基因序列多态性研究 [J]. 中国水产科学, 2007, 14 (3): 369-376.
SU T F, HUANG J H, WU J F,etal. Genetic polymorphism of mitochondrial DNA sequences inBabyloniaareolataandBabyloniaformosae[J]. Journal of Fishery Sciences of China, 2007, 14 (3): 369-376.
[39] LIN X Z, ZHANG X D, XIAO S,etal. Phylogeny of the cuttlefishes (Mollusca: Cephalopoda) based on mitochondrialCOⅠ and 16S rRNA gene sequence data [J]. Acta Oceanologica Sinica, 2004, 23 (4): 699-707.
[40] GRANT P R, GRANT B R. Hybridization of bird species [J]. Science, 1992, 256 (5054): 193-197.
[41] 王 旭. 黄口荔枝螺和疣荔枝螺遗传多样性研究 [D]. 浙江海洋学院, 2013.
WANG X. Research on genetic diversity ofThaisluteostomaandThaisclavigera[D]. Zhejiang Ocean University, 2013.
[42] 李宏俊, 张晶晶, 袁秀堂, 等. 利用线粒体COⅠ和微卫星标记分析文蛤7个地理群体的遗传变异 [J]. 生态学报, 2016, 36 (2): 499-507.
LI H J, ZHANG J J, YUAN X T,etal. Genetic diversity and differentiation of seven geographical populations of hard clam (Meretrixmeretrix) assessed byCOⅠ and microsatellite markers [J]. Acta Ecologica Sinica, 2016, 36 (2): 499-507.
[43] 陈灵芝. 中国的生物多样性现状及其保护对策 [M]. 北京: 科学出版社, 1993: 99-113.
CHEN L Z. Biodiversity status and protection countermeasures in China [M]. Beijing: Science Press, 1993: 99-113.
[44] 李玉龙, 董 婧, 王 彬, 等. 基于线粒体COⅠ基因的海蜇不同地理群体遗传特征 [J]. 应用生态学报, 2016, 27 (7): 2340-2347.
LI Y L, DONG J, WANG B,etal. Genetic characterization of different populations ofRhopilemaesculentumbased on the mitochondrialCOⅠ sequence [J]. Chinese Journal of Applied Ecology, 2016, 27 (7): 2340-2347.
[45] 朱 叶, 章 群, 李贵生, 等. 中近海角木叶鲽(Pleuronichthyscornutus)种群遗传多样性研究[J]. 海洋通报, 2012, 31 (5): 552-556.
ZHU Y, ZHANG Q, LI G S,etal. Genetic diversity of fourPleuronichthyscornutuspopulations in coastal waters of China [J]. Marine Science Bulletin, 2012, 31 (5): 552-556.
[46] FRANKHAM R, BALLOU J D, BRISCOE D A. Introduction to Conservation Genetics [M]. Cambridge: Cambridge University Press, 2002.
[47] AVISE J C, NEIGEL J E, ARNOLD J. Demographic influences on mitochondrial DNA lineage survivorship in animal population [J]. Journal of Molecular Evolution, 1984, 20 (2): 99-105.
[48] CARLTON J T. The Light and Smith Manual:Intertidal Invertebrates from Central California to Oregon [M]. California: University of California Press, 2007: 701-713.
[49] SIGWART J D. Morphological cladistic analysis as amodel for character evaluation in primitive livingchitons (Polyplacophora, Lepidopleurina) [J]. American Malacological Bulletin, 2009, 27 (1-2): 95-104.
