微生境尺度下有蹄类动物空间季节利用互作关系研究
2018-06-26胡素贤王晶晶姜广顺
杨 帆 胡素贤 王晶晶 郭 楷 姜广顺*
(1.东北林业大学野生动物资源学院,哈尔滨,150040;2.东北林业大学图书馆,哈尔滨,150040)
群落生态学研究中一个主要目标是了解物种共存的条件。竞争排斥原则指出,生态位相似的物种竞争激烈,难以共存,往往导致生态位分离[1]。因此,共存物种的分布区域必须发生一定程度的差异,这可能发生在时间、营养或生境选择水平上。异质景观促使相同营养级别的物种共存现象尤为明显,不同生境选择和利用策略是促进同域分布的主要原因之一[2]。对微生境尺度下物种间生境利用互作关系的分析至今尚未得到充分研究,需要更小尺度的生境信息才能更准确地对适宜生境进行区分和质量评估,并在更大尺度上发现不具明显效应的重要生境利用特征和规律[3]。
随着群落物种数量的增加,其中任何一个物种的生境选择和利用都可能受到影响。物种和物种之间的相互作用可能取决于群落内其他物种的种类和丰度。这种分析可能适合于评估任何特定生境中目标物种的动态,但它本身不可能解释所有群落成员的共存[4]。目前,群落物种共存的研究多集中于热带地区,而在高纬度地区,由于天气寒冷,植物生长周期长,所以动物会依赖环境特点选择有不同的生存策略,空间关系也有所变化。而森林火烧会对生境环境进行重塑,影响物种群落的更替[5-6],2002年和2003年间大兴安岭地区曾发生过小规模火烧,因此,也将森林火烧作为生境选择影响的因素之一。本研究比较了大兴安岭双河国家级自然保护区同域分布的4种有蹄类动物(西伯利亚狍Capreolus pygargus、野猪Sus scrofa、马鹿Cervus elaphus、驼鹿Alces alces)在微生境尺度下空间利用的互作关系,并假设植被和景观结构等生境因子是驱动有蹄类集团微生境利用互作关系的主要因素。
1 材料与方法
1.1 研究区域概况
黑龙江双河国家级自然保护区处于中国北部边疆大兴安岭山地北部,北部及东北部与俄罗斯隔黑龙江相望。保护区地理坐标为 E124°52'48″~125°32'03″,N52°54'25″~ 53°12'08″,总面积 88849 hm2。地形南高北低,坡度较缓,相对高度较小,海拔在200~515 m之间。冬季漫长而寒冷,年均气温为-4.3℃,极端最低气温-45.8℃,极端最高气温38℃,≥10℃积温为1500~1800℃。有霜期从9月上旬至翌年5月下旬,冰冻期可达7个月之久,全年平均积雪期为165~175 d,平均冻土深为2.5~3.0 m左右。在局部低洼沼泽地带有岛状永冻层分布。年平均日照时数2527 h,植物生长期为110 d左右。
图1 双河国家级自然保护区位置图Fig.1 Map of Shuanghe National Nature Reserve
保护区分布着大兴安岭地区地带性植被,主要是混有阔叶树的寒温带针叶林,外貌与东西伯利亚针叶林相近,是以兴安落叶松为单优势种的针叶林,混有一些东北植物区系成分的阔叶树种,林下灌木和草本植物也同样是具有东北植物区系成分的种类,垂直分布现象不明显;部分低地、河漫滩分布着非地带性植被灌丛、草甸、沼泽和草塘等湿地植被。动物主要以适应大兴安岭寒冷气候条件的寒温带种类为主[7]。
1.2 数据采集与提取
1.2.1 自动相机数据
2013年6月~2015年12月,按照1对相机/每10 km2的密度布设,每3个月取1次数据,并更换SD卡和电池,共架设124点次。相机的架设地点一般选择山脊与水源附近等动物偏爱的兽道边。布设相机时,对周边的环境信息进行记录,包括火烧情况、隐蔽级等。
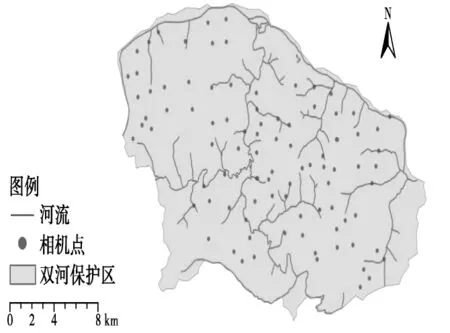
图2 双河国家级自然保护区自动相机布设图Fig.2 Locations of camera traps in Shuanghe National Nature Reserve
获取照片后进行人工筛选,在进行表格录入时记录拍摄的动物物种、行为、人为干扰类型、相机编号、相机点坐标、日期时间等。在30 min以内拍到的同一物种按照拍到1次计算。每个相机位点作为1行(row),每天为1列(column),有发现物种的点,按照时间在对应的相机位点和时间的格内填“1”,没有发现的位点都用 “0”表示[8-9]。
1.2.2 微生境变量数据
从中国科学院计算机网络信息中心地理空间数据(http:∥www.gscloud.cn)处下载双河地区 GDEM DEM 30 m分辨率数字高程数据,并从中国地理数据云下载保护区最近2010年MODIS影像,提取了15个变量,并加入记录的火烧情况和隐蔽级(表1)。

表1 微生境变量的定义和描述Tab.1 Definitions and descriptions of microhabitat variables
1.2.3 生境变量相关性分析
所有生境变量进行空间自相关关系检验。若2个变量相关系数的绝对值大于0.6,则认为二者间存在较强的空间相关性,二者不能同时进入同一个模型[10]。使用R软件(V3.4.1)中的Spearman相关分析筛选,相关系数大于0.6的变量合理删除。最后去掉了海拔和落叶松桦林2个变量,将剩余的15个生境变量作为协变量放入到占有模型中。
2 数据分析
2.1 冷季和暖季有蹄类微生境利用
根据大兴安岭地区的气候特征以及研究需求,将每年的5月到10月划分为暖季,11月至次年4月划分为冷季,同时相机数据按暖季和冷季分别处理。使用单季节占有模型(single season model occupancy model)并根据Backward筛选法[11]建立最优模型,△AIC值越小,模型拟合效果越好,模型权重越大,则越重要。
(1)首先建立空模型(null model),占有变量中不带入任何环境协变量,再拟合一个全模型,将全部环境协变量作为占有变量带入模型中。这2个模型拟合后的AIC值,作为即将开始拟合占有变量的参考。
(2)全模型中通过不断增加和减少方程中的环境协变量找到最小△AIC值的方程。
2.2 有蹄类物种空间利用互作因子
物种间单季节模型(two-species single season model)可以计算2个物种不同情况下的占有概率[12]。其中物种互作因子(SIF,species interaction factor)则被认为是反映物种间空间分布模式相关性的一个指标,SIF等于1时表明物种间的空间分布是相互独立,大于1时物种间的空间分布模式趋向于重合,而小于1时表明物种间的空间分布模式趋向于分离[13]。将2个物种的数据矩阵放入Presence软件,选择Two species-Single season model。
3 结果
3.1 冷暖季微生境变量利用
3.1.1 狍
研究结果显示,暖季狍的最优占有概率模型包含坡向、坡度、河流、居民点、公路、季节道、农田、草原草甸、阔叶林、阔叶灌木林、针叶林、针叶灌木林、隐蔽级13个变量(表2)。与坡向、居民点、公路、季节道、阔叶林、针叶灌木林呈正相关,即坡向越大,距居民点、公路、季节道、阔叶林、针叶灌木林越远,占有概率越大。与坡度、河流、农田、草原草甸、阔叶灌木林、针叶林、隐蔽级为负相关,即坡度越缓,距河流、农田、草原草甸、阔叶灌木林、针叶林距离越近,隐蔽级越小,占有概率越大(表3)。冷季最优占有概率模型包含坡度、河流、居民点、公路、季节道、阔叶灌木林、针叶林、针叶灌木林8个变量(表2)。与坡度、居民点、公路、季节道、针叶灌木林呈正相关,坡度越陡,距居民点、公路、季节道、针叶灌木林越远,占有概率越大。与河流、阔叶灌木林、针叶林为负相关,即距河流、阔叶灌木林、针叶林越近,占有概率越大(表3)。
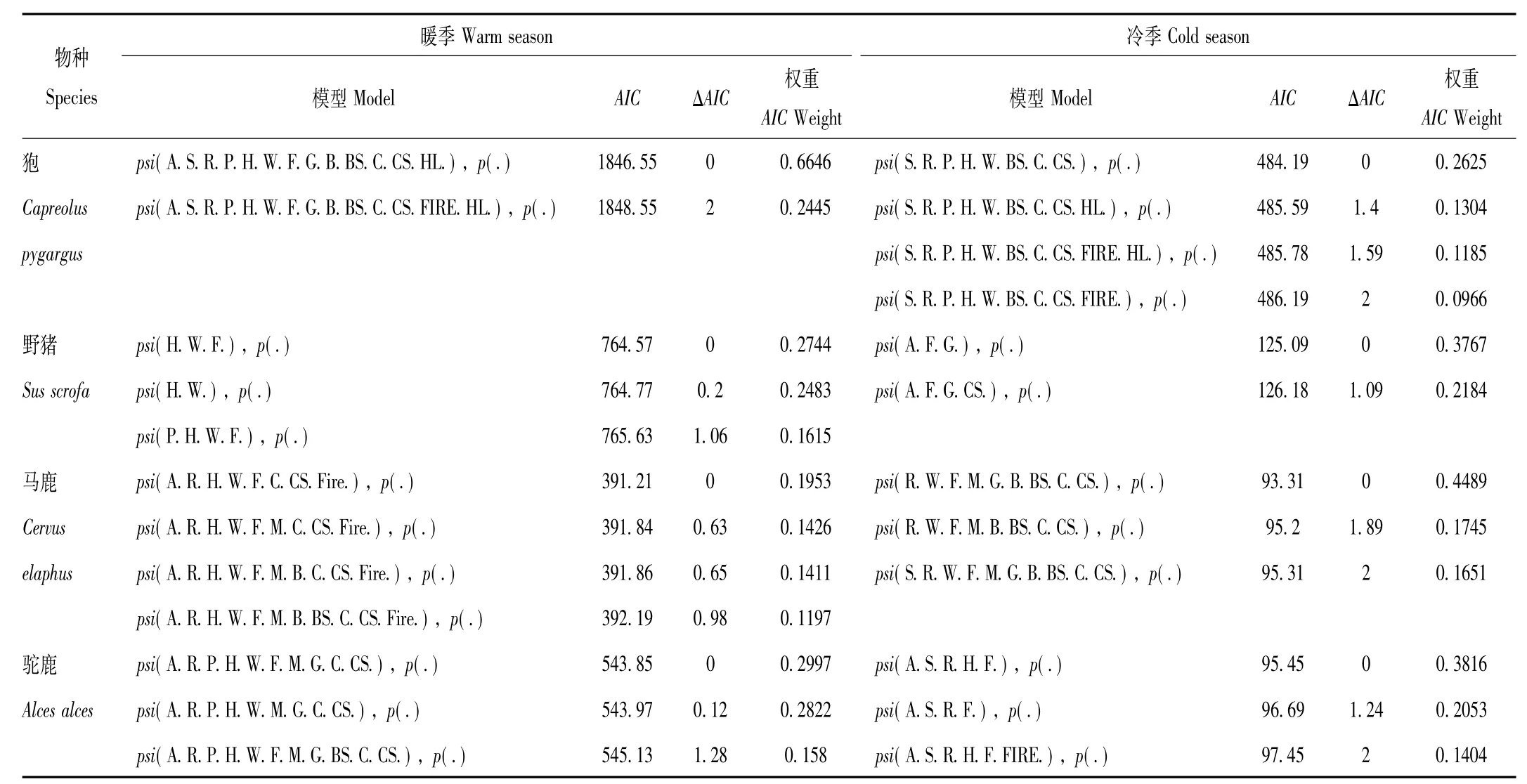
表2 双河国家级自然保护区4种有蹄类占有概率的顶级模型Tab.2 The top models for predicting site occupancy of four ungulates in Shuanghe National Nature Reserves
3.1.2 野猪
研究结果显示,暖季野猪的最优占有概率模型包含公路、季节道、农田3个变量(表2)。与公路、季节道呈正相关,距公路、季节道越远,占有概率越大。与农田为负相关,离农田越近,占有概率越大(表3)。冷季最优占有概率模型包含坡向、农田、草原草甸3个变量(表2)。与坡向呈正相关,坡向越大,占有概率越大,与农田、草原草甸为负相关,选择离农田、草原草甸近的生境(表3)。
3.1.3 马鹿
研究结果显示,暖季马鹿的最优占有概率模型包含坡向、河流、公路、季节道、农田、针叶林、针叶灌木林和火烧迹地8个变量(表2)。与公路、季节道、针叶林、针叶灌木林呈正相关,即距公路、季节道、针叶林、针叶灌木林距离越远,占有概率越大。与坡向、河流、农田、火烧迹地为负相关,即坡向越小,距河流、农田越近,距火烧迹地越远,占有概率越大。选择坡向较小,距离公路、季节道远,干扰大的,离农田、火烧迹地近,食物丰富的生境,回避针叶林和针叶灌木林。冷季最优占有概率模型包含河流、季节道、农田、草原草甸、湿地沼泽、阔叶林、阔叶灌木林、针叶林、针叶灌木林9个变量(表2)。与季节道、农田、草原草甸、阔叶灌木林、针叶林呈正相关,距季节道、农田、草原草甸、阔叶灌木林和针叶林距离越远,占有概率越大。与河流、湿地沼泽、阔叶林、针叶灌木林为负相关,即距河流、湿地沼泽、阔叶林、针叶灌木林距离越近,占有概率越大。选择距水源近的阔叶林和针叶灌木林,回避草原草甸、阔叶灌木林和针叶林(表3)。
3.1.4 驼鹿
研究结果显示,暖季驼鹿的最优占有概率模型包含坡向、河流、居民点、公路、季节道、农田、草原草甸、湿地沼泽、针叶林、针叶灌木林10个变量(表2)。与居民点、公路、季节道呈正相关,距居民点、公路、季节道距离越远,占有概率越大,与坡向、河流、农田、草原草甸、湿地沼泽、针叶林、针叶灌木林为负相关,即坡向越小,距河流、农田、草原草甸、湿地沼泽、针叶林、针叶灌木林越近,占有概率越大。选择坡向小,距离人为干扰远的草原草甸、湿地沼泽、针叶林和针叶灌木林生境(表3)。冷季最优占有概率模型包含坡向、坡度、河流、公路、农田5个变量(表2)。与农田呈正相关,距离农田越远,占有概率越大。与坡向、坡度、河流、公路为负相关,坡向越小,坡度越缓,距河流、公路越近,占有概率越大(表3)。
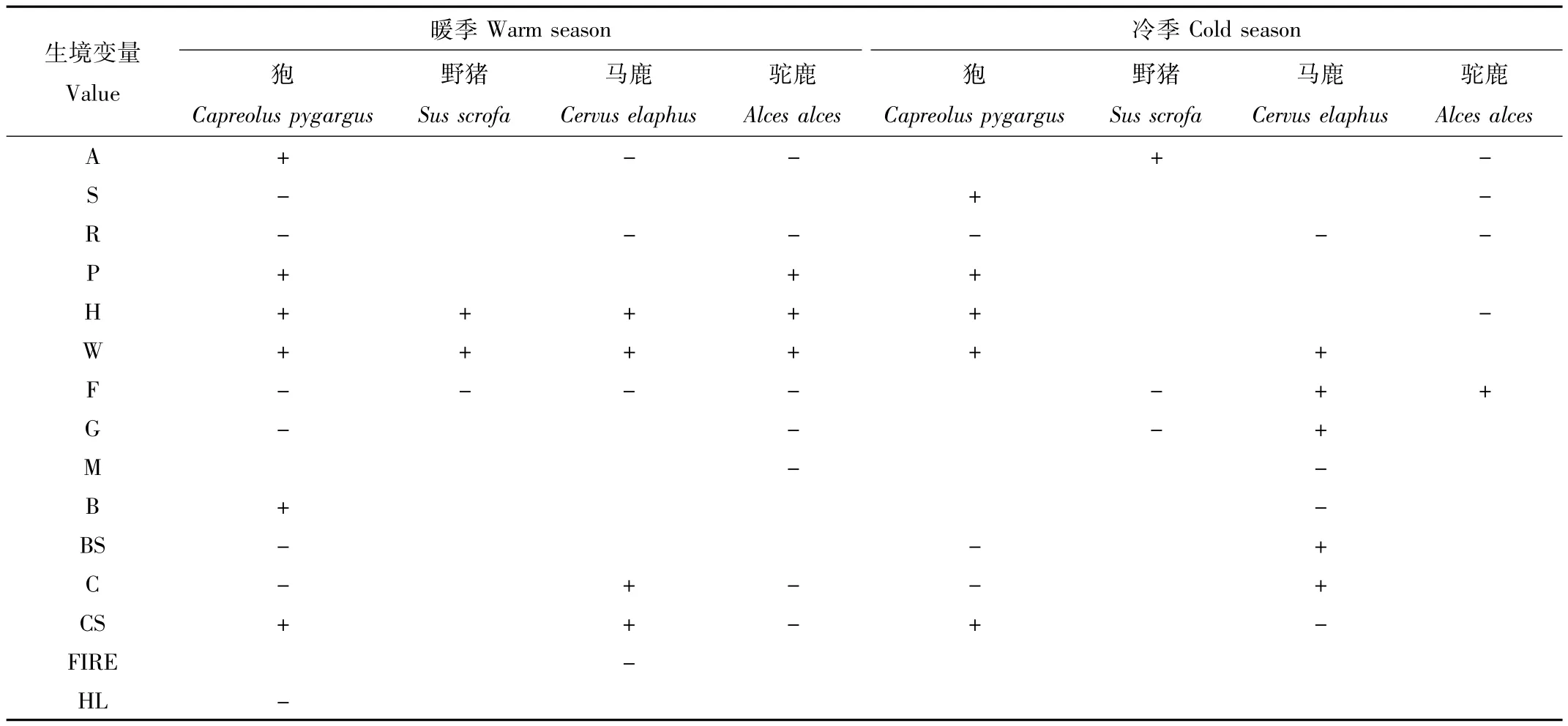
表3 不同季节顶级模型的β系数方向Tab.3 β coefficient directions of the top model for different seasons
3.2 4种有蹄类空间利用互作关系
暖季时,研究结果显示狍与野猪的空间关系呈不明显的共存关系,马鹿与狍的空间关系呈不明显的共存关系,马鹿与野猪呈回避关系,驼鹿与狍呈不明显的回避关系,驼鹿与野猪呈明显回避关系,驼鹿与马鹿呈明显共存关系(表4,图3)。冷季时,狍与野猪的空间关系呈回避关系,马鹿与狍的关系呈不明显的回避关系,马鹿与野猪呈明显的回避关系,驼鹿与狍呈独立关系,驼鹿与野猪呈明显回避关系,驼鹿与马鹿呈明显回避关系(表4,图3)。
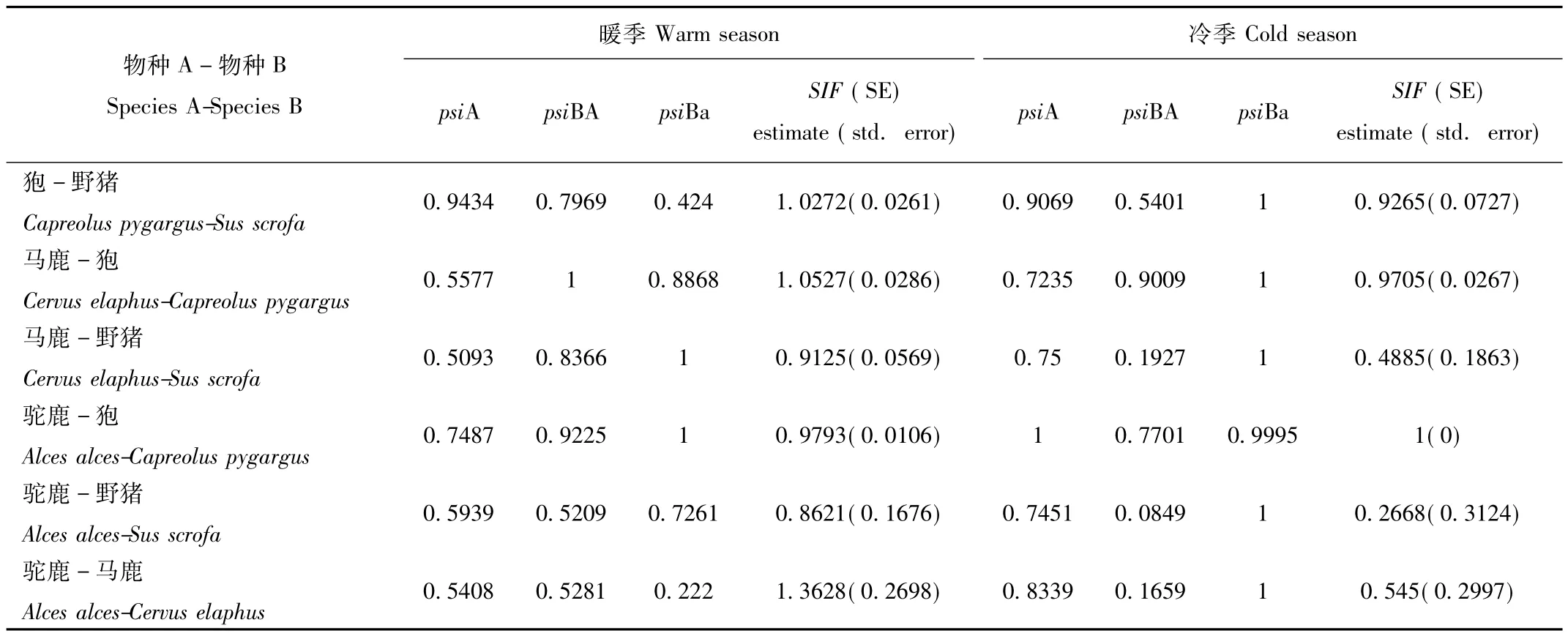
表4 不同季节4种有蹄类物种相互作用结果Tab.4 Species interaction factors of four ungulates in two seasons
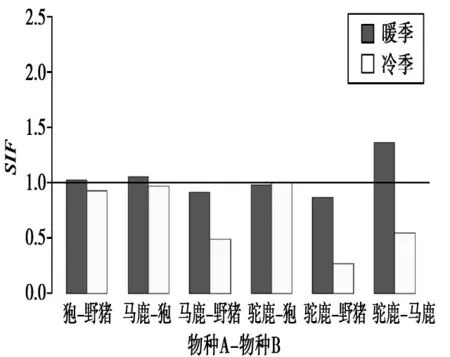
图3 有蹄类不同季节物种相互作用因子变化Fig.3 Species interaction factors of ungulates in different seasons
4 讨论
4.1 4种有蹄类不同季节生境变量利用
有蹄类对于生境的选择和利用,主要是在食物和遮蔽这两个因素间权衡的结果[14]。本研究表明,狍在两个季节,共同利用了距河流近,阔叶灌丛和针叶林的生境,阔叶灌丛的枝叶多,适口性好,可以被狍取食,针叶林又提供了比较干爽的地方便于休息[14]。回避了居民点、道路这两种人为干扰,但在暖季又选择了离农田近的地区,可能是农田能产生一定食物,使得狍并不完全回避此区域。野猪在两个不同的季节,共同选择距农田近的区域,可能是在农田较易获得食物,尤其是冷季。但暖季更多是回避居民点和道路的干扰,而在冷季,草原草甸中的水被冻住,便于行走,且所处位置是大兴安岭地区海拔低的地方,这里有较多野猪可食用苔草,所以野猪冷季主要选择了食物相关的生境。马鹿在两个季节共同利用了河流、季节道和针叶林3个影响其生存的生境变量,回避季节道和针叶林,趋向于河流的生境。针叶林适口性差,且下可吃的食物较少,对于马鹿,不是优良的生境[15]。驼鹿偏向利用坡向大,距河流距离近的地区。在暖季,驼鹿利用了农田、草原草甸、湿地沼泽、针叶林、针叶灌丛多个生境类型,草原草甸、湿地沼泽可以提供食物和补充微量元素,而针叶林和针叶灌丛则提供了休息的地方[16]。
总之,在暖季,4种动物都对道路有着回避现象,而对农田又有着喜好,并不完全排斥人为干扰。在冷季,4种动物生境变量选择各有差异,但3种鹿科动物(狍、马鹿、驼鹿)两个季节都偏爱距河流近的生境。
4.2 影响4种有蹄类动物物种空间利用互作的生境因子
物种的空间分布是受多种因素影响的,一个是受物种间的相互关系影响,也就是物种间内部的作用,包括捕食、竞争、寄生、协同共生等,另外还会受到外部作用力即生境变量的影响[17-18]。
物种空间分布模式相关性分析结果,可以看出物种空间分布模式相关性存在着季节上的差异,而季节差异代表的是生境变量的差异,这说明物种空间分布模式相关性可能不仅受物种间关系的影响,而且还会受到生境变量的影响。另外总体上来看冷季物种空间相关性指标相较于夏季都会有明显的下降趋势,可能因为冷季食物丰富度下降[19],物种在环境利用上的压力更大,物种空间分布模式的分离有利于动物群落对有限环境资源的高效利用从而缓解资源短缺的压力。另外,物种生境选择的模型数据也能很好地解释这些变化。例如,暖季时驼鹿和马鹿空间分布都受坡向、河流、公路、季节道、农田这5种生境变量的影响且受影响的方向一致,所以马鹿和驼鹿的空间分布模式趋向于重叠,但是到冷季,两者利用生境变量的模型却出现了巨大的差异,只有河流和农田两种生境变量仍会对马鹿和驼鹿有相同的影响,此时马鹿和驼鹿的空间分布模式呈现相互回避的现象;又如,马鹿和野猪,暖季时虽然两者都受公路、季节道、农田的同向影响,但是公路、季节道、农田是影响野猪空间分布的主要生境变量,而马鹿还会受到坡向、河流、针叶林、针叶灌丛、火烧迹地等生境变量的影响,所以两者对生境变量的选择是一个包含关系,从而导致了空间分布模式有着轻微的分离。冷季时马鹿和野猪对生境变量的需求出现了完全的错位,所以两者空间分布模式的回避效应更加显著;驼鹿和野猪空间分布模式关系的变化也是相同的。
总之,双河国家级自然保护区4种有蹄类动物冷暖季在微生境利用上存在着显著差异,生境变量利用上的差异影响物种空间利用的互作关系,为该地区有蹄类动物集团的保护提供了科学参考。
致谢:感谢国家林业局野生动物与自然保护区管理司“虎、东北豹资源调查技术研究”和“东北虎、东北豹种群及栖息地调查评估标准制定及信息汇总”项目的资助。感谢研究过程中,实验室的师兄、师姐给予的很多帮助,感谢他们的付出。野外工作得到了项目区双河国家级自然保护区工作人员的大力支持和帮助,在此表示感谢。
[1] Hardin G.The competitive exclusion principle [J].Science,1960,131(3409):1292-1297.
[2] Rosenzweig M L,Abramsky Z,Brand S.Estimating species interactions in heterogeneous environments [J].Oikos,1984,43(3):329-340.
[3] Fernández N,Delibes M,Palomares F.Landscape evaluation in conservation:molecular sampling and habitat modeling for the Iberian lynx[J].Ecological Applications,2006,16(3):1037-1049.
[4] Morris D W.Toward an ecological synthesis:a case for habitat selection [J].Oecologia,2003,136(1):1-13.
[5] Kotliar N B,Hejl S J,Hutto R L,et al.Effects of fire and post-fire salvage logging on avian communities in conifer-dominated forests of the western United States[J].Studies in Avian Biology,2002,25:49-64.
[6] 安森鹏,高振岭,王磊.林火对森林生态系统的影响[J].科技信息,2013(6):90-91.
[7] 郭楷.黑龙江双河保护区猞猁与猎物冬季生境相关性研究[D].哈尔滨:东北林业大学,2016.
[8] MacKenzie D I,Nichols J D,Lachman G B,et al.Estimating site occupancy rates when detection probabilities are less than one [J].Ecology,2002,83(8):2248-2255.
[9] 张鹏.基于自动相机技术的兽类监测及狍栖息地选择研究[D].哈尔滨:东北林业大学,2015.
[10] Soto C,Palomares F.Coexistence of sympatric carnivores in relatively homogeneous Mediterranean landscapes:functional importance of habitat segregation at the fine-scale level[J].Oecologia,2015,179(1):223-35.
[11] Akaike H.A new look at the statistical model identification [J].IEEE Transactions on Automatic Control,1974,19(6):716 -723.
[12] MacKenzie D I,Bailey L L,Nichols J D.Investigating species cooccurrence patterns when species are detected imperfectly [J].Journal of Animal Ecology,2004,73(3):546-555.
[13] Steen D A,McClure C J W,Brock J C,et al.Snake co-occurrence patterns are best explained by habitat and hypothesized effects of interspecific interactions[J].Journal of Animal Ecology,2014,83(1):286-295.
[14] Mysterud A,stbye E.Cover as a habitat element for temperate ungulates:effects on habitat selection and demography[J].Wildlife Society Bulletin(1973-2006),1999,27(2):385-394.
[15] Stephens P A,Zaumyslova O Y,Hayward G D,et al.Analysis of the long-term dynamics of ungulates in Sikhote-Alin Zapovednik,Russian Far East[R].The USDA Forest Service and Sikhote-Alin Zapovednik,2006.
[16] Jiang Guangshun,Ma Jianzhang,Zhang Minghai,et al.Assessing microhabitat use by roe deer and moose in China[J].Wildlife Research,2009,36(2):134-142.
[17] Pulliam H R,Danielson B J.Sources,sinks,and habitat selection:a landscape perspective on population dynamics[J].The A-merican Naturalist,1991,137:S50 -S66.
[18] Matthiopoulos J,Fieberg J,Aarts G,et al.Establishing the link between habitat selection and animal population dynamics[J].Ecological Monographs,2015,85(3):413 -436.
[19] Suttie J M,Goodall E D,Pennie K,et al.Winter food restriction and summer compensation in red deer stags(Cervus elaphus) [J].British Journal of Nutrition,1983,50(3):737-747.
