濒危植物狭叶罗伞形态特征及其群落和种群学研究
2018-06-25杨锦超
杨锦超 杜 凡 石 明 代 俊 宋 放
(西南林业大学林学院,云南 昆明 650224)
狭叶罗伞 (Brassaiopsisangustifolia) 为五加科 (Araliaceae) 灌木。武素功于1958年初在云南元江三区小竹箐采到标本 (武素功827),由冯国楣在1979年建立新种[1]。近60年来,其分布区有过多次调查,但都未发现该物种,也没有任何相关研究的报道。文军等[2]在研究东南亚罗伞属 (Brassaiopsis) 植物进化关系时,也未将此种列入其中。狭叶罗伞花序侧生于无叶老枝 (茎花)、花序休眠等特征,与同属其他种有显著区别[1],在五加科中也极为特殊,是研究五加科系统学的重要材料。这是一个未被关注、知之甚少,但形态特别的狭域特有种。
2006年项目组在进行元江自然保护区科考时,再次发现狭叶罗伞。近年来因在其周围建风电,威胁加大,对其研究迫在眉睫。本研究根据近2年来的实地调查、文献整理、标本研究等途径,对其分布现状、种群、群落及繁殖等特征进行研究,为狭叶罗伞的深入研究及有效保护提供科学依据。
1 研究区概况
研究区位于元江县咪哩乡甘岔村,元江河谷西岸山地,东经101°49′~101°52′,北纬23°32′~23°34′,海拔2 100~2 250 m。年平均温度9.7~11.7 ℃,年降水量1 646~1 803 mm,生境湿润;土壤为山地黄棕壤[3]。山体中下部已被广泛开垦成耕地。
研究区所属的元江县地处滇中高原南部边缘,是西南季风的控制范围。其西南面的哀牢山阻挡西南暖湿气流北上,在迎风坡形成降雨区,在背风坡——元江河谷形成 “焚风效应”[4],河谷地带形成半萨王纳植被。其独特的地理条件形成了元江山柑 (Cappariswui)、瘤果三宝木 (Trigonostemontuberculatus) 等8个狭域特有植物[5],近年又发现希陶木属 (Tsaiodendron) 等狭域特有植物类群[6]。
2 研究方法
2.1 资料收集
查阅了CVH中国数字植物标本馆 (http://www.cvh.ac.cn/)、NSII国家标本资源共享平台 (http://www.nsii.org.cn/2017/)、GlobalPlants (http://plants.jstor.org/)、昆明植物所、北京植物所和广西植物所的标本;查阅了 《Flora of China》、《中国植物志》、《云南植物志》 及 《西藏植物志》 等文献。
2.2 野外调查
调查时间为2016年11月、2017年1月、2017年6月、2017年11月,采用典型取样法,选取5个20 m × 25 m的样方。记录样方海拔、坡度、坡向和人为干扰程度等;乔木层设置5 m × 5 m的网格,胸径大于5 cm的乔木进行每木调查;在样方4个角及中心各设置1个5 m × 5 m和1 m × 1 m的小样方,分别调查灌木层 (包括胸径小于5 cm的乔木幼树) 和草本层,记录物种株数、平均高度、平均胸径、盖度等。另外实测样方内每株狭叶罗伞的株高、基径、冠幅及以水沟为中心线记录狭叶罗伞的分布位置,调查区域内的狭叶罗伞植株,并观察狭叶罗伞不同时期的物候特征。在狭叶罗伞模式标本产地 (元江三区小竹箐) 调查,未发现狭叶罗伞植株。
2.3 分析方法
按照惯例,将树高 (H) 大于15 m的植株计入乔木上层,树高5 m至15 m的计入乔木下层。5 m以下的木本植物计入灌木层,包括乔木幼树和真正的灌木。攀援灌木、藤本、附生植物计入层间层。草本植物与蕨类植物一起计入草本层。计算乔木层、灌木层及草本层的相对密度、相对频度、相对优势度和重要值[7-8]。
根据狭叶罗伞的树高,除更新苗外,以0.5 m划分高度级,2.5 m以上的植株较少,划为单独一个高度级,共7个高度级,分别是:Ⅰ级为更新苗,即2年生以下实生苗 (茎木质化低、光滑柔嫩)H< 0.2 m;Ⅱ级为H< 0.5 m,除更新苗外;Ⅲ级为0.5 m ≤H< 1 m;Ⅳ级为1 m ≤H< 1.5 m;Ⅴ级为1.5 m ≤H< 2 m;Ⅵ级为2 m ≤H< 2.5 m;Ⅶ级为H≥ 2.5 m。
3 结果与分析
3.1 狭叶罗伞的记录情况
经查,国外无狭叶罗伞标本;国内除模式标本外,尚有2份标本鉴定为狭叶罗伞,其基本信息见表1。

表1 狭叶罗伞标本Table 1 The specimens identified of B.angustifolia
表1中,采自广西的付雨节0097号标本,其花序顶生、总花梗较细,与狭叶罗伞模式标本花序侧生于无叶老枝上、总花梗粗壮区别明显;采自西藏墨脱的程树志、李渤生00214号标本,其叶片6~7裂、先端渐尖、叶脉弧形、上面不显及叶下面光滑无毛,与狭叶罗伞模式标本叶片小于等于5裂、先端长渐尖、叶脉较直边缘分叉、上面明显及叶下面疏背锈色星状绒毛区别明显。因此,这两号标本都是错误鉴定,狭叶罗伞标本仅有采自元江县的模式标本。
《云南种子植物名录》 记录狭叶罗伞的产地和 《云南植物志》 一致,均为云南南部 (元江)[1,9];《Flora of China》 记录狭叶罗伞产云南南部及越南[10]。作者查阅大量资料未见到越南的标本,也没有从越南的同行中获得相关信息,因此狭叶罗伞在越南的分布尚需进一步考证。
3.2 狭叶罗伞的群落特征
狭叶罗伞分布在海拔2 100~2 250 m,群落类型属于中山湿性常绿阔叶林。群落高达20~30 m,树干较通直,胸径可达70 cm,树干上有厚达5 cm的苔藓和地衣,附生种子植物和蕨类植物,林下有明显的宁南方竹 (Chimonobambusaningnanica) 层片。这些群落学特征均反映其生境的高度湿润。
3.2.1物种组成
狭叶罗伞群落物种组成见表2。5个样方内共计维管植物52科90属107种,未发现裸子植物。种类较多的科有樟科 (Lauraceae) (7属10种)、山茶科 (Theaceae) (4属7种)、蔷薇科 (Rosaceae) (3属6种)、壳斗科 (Fagaceae) (3属5种)、百合科 (Liliaceae) (5属5种)。

表2 狭叶罗伞群落物种组成Table 2 Community species composition of B.angustifolia
3.2.2数量特征
狭叶罗伞样方内植物的重要值见表3~5。

表3 狭叶罗伞样方乔木层植物的重要值Table 3 Important value of woody plants in the B.angustifolia plot
群落乔木层盖度85%~90%,物种计31种,占物种总数的29%。其中重要值前10的见表3,作为乔木层优势种的7种,平均高度15.7 m,重要值之和65.32,分别为疏齿栲 (Castanopsisremotidenticulata)、壶壳柯 (Lithocarpusechinophorus)、截果柯 (Lithocarpustruncatus)、硬斗柯 (Lithocarpushancei)、红花木莲 (Manglietiainsignis)、黄牛奶树 (Symplocoslaurina)、小花檬果樟 (Caryodaphnopsishenryi),前4种为乔木上层优势种,后3种为乔木下层优势种。
灌木层盖度15%~35%,平均高1.7 m,物种计41种,占物种总数的38.32%。乔木幼树较少,计10种 (表4),其中重要值最大是刀把木为2.86。真正的灌木中重要值前10的见表4,重要值较大的有宁南方竹和狭叶罗伞,其中狭叶罗伞的重要值为16.17。

表4 狭叶罗伞样方灌木层植物的重要值Table 4 Important value of the shurbs in the B.angustifolia plot
草本层盖度8%~25%,平均高0.3 m,物种计23种,占物种总数的21.50%。其中重要值前10的见表5,重要值之和71.13。这些物种多为耐阴湿植物。
层间植物较发达,藤本植物有叉须崖爬藤 (Tetrastigmahypoglaucum)、扶芳藤 (Euonymusfortunei)、丛林素馨 (Jasminumduclouxii) 等;附生植物丰富,主要有苔藓、蕨类及杜鹃花科 (Ericaceae) 的一些种类,如树平藓 (Homaliodendronflabellatum)、滇线蕨 (Colysiselliptica)、假友水龙骨 (Polypodiodessubamoena)、带叶瓦韦 (Lepisorusloriformis)、白花树萝卜 (Agapetesmannii) 等。
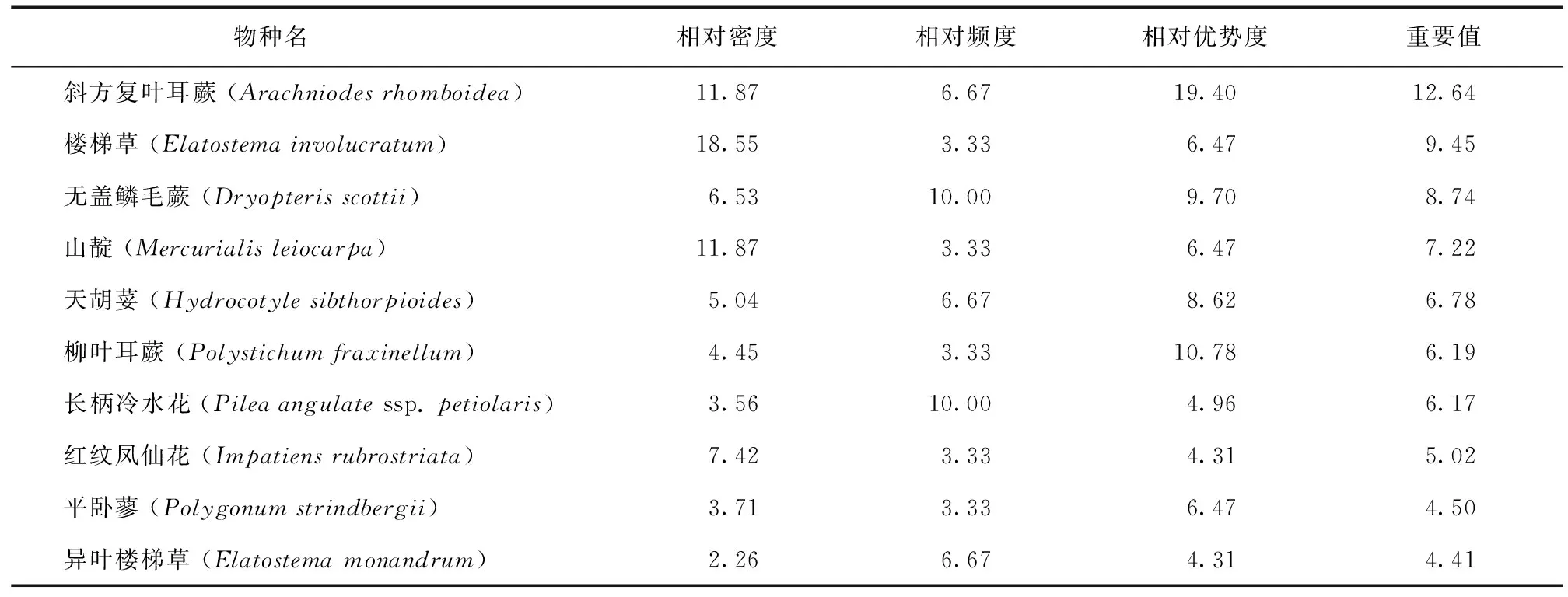
表5 狭叶罗伞样方草本层植物的重要值Table 5 Important value of the herbs in the B.angustifolia plot
3.3 狭叶罗伞的种群特征
狭叶罗伞的种群结构特征见表6。狭叶罗伞种群面积不足3公顷,逐一记录到152株。5个样方中,样方1位于山坡上,仅有1株植株,生长于树基凹处,其余4个样方位于沟箐边。样方内植株共计70株,平均高度0.80 m,平均基径1.52 cm,平均冠幅0.75 cm × 0.73 cm,无自然枯死植株,最高植株5.5 m,主干死亡,在基部又萌发出新枝。狭叶罗伞茎上大多有明显折断过的痕迹。
3.3.1狭叶罗伞分布
种群的空间分布格局是由种群生物学特性、种内种间关系及环境因素综合影响形成的。5个样方内的狭叶罗伞,绝大多数零星分布于水沟边上5 m之内,山坡少见,偶有植株生于乔木基部的凹塘内。

表6 狭叶罗伞的种群结构特征Table 6 The population structure of B.angustifolia
3.3.2狭叶罗伞种群结构
狭叶罗伞作为1种生长缓慢的灌木,径级变化不大,很难用胸径或基径的粗度推断年龄,故用高度划分代替龄级进行种群结构特征分析,第Ⅰ高度级对应第Ⅰ龄级,依次类推,得到狭叶罗伞种群的年龄结构比例:Ⅰ龄级 (更新苗) 占2.86%;Ⅱ龄级 (幼树) 38.57%;Ⅲ龄级 (小树) 占27.14%;Ⅳ龄级 (成年树) 占10%;Ⅴ龄级 (中树) 占11.43%;Ⅵ龄级 (大树) 占7.14%;Ⅶ龄级 (老树) 占2.86%。图1以龄级比例作为纵坐标,以龄级作为横坐标,分析种群动态变化。

图1狭叶罗伞种群年龄结构
Fig. 1 Population age structure ofB.angustifolia
根据种群的年龄结构,一般将种群划分为增长型、稳定型和衰退型。狭叶罗伞植株高度超过1 m时,开始开花 (即Ⅳ龄级),样地内Ⅰ、Ⅱ、Ⅲ龄级植株还不具备繁殖能力,个体数比例占总株数的68.57%,表明狭叶种群属于增长型。
3.3.3狭叶罗伞种子散布及更新
狭叶罗伞主要分布于沟箐边,种子主要靠重力传播,掉落的种子顺水移动,在沟边萌发长大;成熟果实上有明显的啮齿类动物啃食痕迹;部分植株生于乔木基部凹塘内,凹塘内的土壤松软,往里为动物巢穴,在其周围未发现其他狭叶罗伞植株,其明显为动物搬运储存的种子萌生长大。5个样方内调查到狭叶罗伞实生苗2株(H为0.13 m和0.15 m),占样方内总株数的2.86%,种群密度8株/hm2。样方内外调查到的152株植株中,实生苗仅5株。这表明狭叶罗伞更新能力较低。
3.4 狭叶罗伞的形态特征
狭叶罗伞生长情况见图2。
3.4.1干形及叶形
狭叶罗伞主干一般不分支,茎最粗可达4 cm,最高可达5.5 m,茎上有圆锥形稍扁的皮刺。狭叶罗伞开花时周围叶脱落,呈老茎生花状。野外观察大量植株,叶从幼苗阶段的单叶,逐渐向掌状深裂、掌状全裂变化 (图2),裂片狭长披针形,具有发育异形叶性现象。叶片基部和叶柄顶端始终没有形成关节,叶片不会从叶柄顶端脱落,所以应为单叶而非指状复叶。

A. 群落外貌;B. 林下沟箐;C. 生长于树基凹处的植株;D. 叶片掌状深裂;E. 花;F. 气生根;G. 叶片掌状全裂和老茎生花;H. 果实;I. 狭叶罗伞幼苗。
图2狭叶罗伞生长情况
Fig.2 The growth status ofB.angustifolia
3.4.2开花特征
狭叶罗伞常绿,叶聚生枝顶。花序生于叶下方老茎上,具有老茎生花特性 (图2)。从出现花蕾到开花需3~4 a时间。花序为3~8个 (而非3~4) 伞形花序排列成总状或窄圆锥状,每个伞形花序约30朵小花。在同一植株上,第1年、第2年、第3年和第4年的花序并存,第1年生花序长约2 cm,伞形花序球形,直径约1 cm;第2年花序轴稍伸长,总花梗变化较小;第3年花序轴和总花梗变化不明显,第4年花序轴、总花梗和花柄明显伸长,总花序达10 cm。花期从11月到次年3月。样方内的70株狭叶罗伞,有花序的14株,占总株数的20%;结实的仅6株,占总株数的8.6%。
3.4.3果实及种子特征
《云南植物志》 中未记录狭叶罗伞果实及种子的特点。调查表明,狭叶罗伞果期为3—6月,果实为浆果,味道微苦带甜,果皮薄,成熟后紫黑色,长圆形,长7~11 mm,宽5~9 mm,柱头宿存约1.5 mm;果柄长7~12 mm;外果皮肉质,内果皮膜质;种子通常侧扁、肾形,多数果实每室2粒种子,部分种子发育不完整,仅1枚种子。
4 结论与讨论
本研究发现狭叶罗伞生长在2 100~2 250 m的中山湿性常绿阔叶林林下沟箐山坡呈零星分布,是罗伞属分布海拔最高的种之一;狭叶罗伞的叶为不裂、掌状深裂、掌状全裂的单叶,而非 《云南植物志》 描述的指状复叶 (掌状复叶);狭叶罗伞长达3~4 a孕花期的现象,笔者将其称为花序休眠,这种现象在有花植物中未见报道,其花序休眠的生物学意义有待进一步研究。花序萌发于二年生茎,为与老茎生花现象区别,笔者将其称为茎花现象。
狭叶罗伞分布区狭窄,种群数量稀少,处于高度濒危状态,其原因可能是狭叶罗伞分布点唯一,只有云南元江县甘岔村,且分布面积小,加之人们开垦耕地、修建风电基站使狭叶罗伞适宜生境更加稀少;也可能是狭叶罗伞孕花期长,果实和种子被动物取食,数量少,严重影响了狭叶罗伞的更新;还有可能是附近村民采笋放牧、人畜践踏使得狭叶罗伞部分植株死亡,尤其是幼苗的死亡。
狭叶罗伞的历史起源、生存繁殖特性及人类影响决定了种群分布特点。虽然其种群年龄结构属于增长型,但其分布狭域、种群数量少、更新困难、原生境面积减少及人畜影响大,使其成为一个狭域脆弱的特有种。根据 《IUCN物种红色名录濒危等级和标准》 3.1版 (第二版)[11],狭叶罗伞符合濒危标准 (EN),即种群未达到极危标准,且推断种群的成熟个体数少于250 (成熟个体数为58),建议纳入IUCN物种红色名录。同时,也符合 《云南省极小种群物种拯救保护规划纲要 (2010—2020年)》 评定标准的第二条,即生境独特、生态幅狭窄、地理分布范围日益缩小而长期处于脆弱状态的物种,建议将其收录为云南极小种群物种中,给予重点保护。
致谢:本研究外业调查得到元江国家级自然保护区管护局工作人员的大力协助,同时得到喻丁香、赵宣武、李娟、马晨晨、肖之强、肖明昆、吕亚媚等同学的帮助,在此一并致以最衷心的感谢。
[1] 中国科学院昆明植物研究所. 云南植物志: 第二卷[M]. 北京:科学出版社, 1979: 471.
[2] 文军, 朱昱苹, Lee C, 等. 东南亚地区五加科植物进化关系的初步研究[J]. 云南植物研究, 2008, 30(4): 391-399.
[3] 哀牢山自然保护区综合考察团. 哀牢山自然保护区综合考察报告集[M]. 昆明: 云南民族出版社, 1988.
[4] 刘方炎, 朱华. 元江干热河谷植被数量分类及其多样性分析[J]. 广西植物, 2005, 25(1): 22-25, 92.
[5] 李海涛, 杜凡. 元江自然保护区种子植物区系特有现象[J]. 林业调查规划, 2008, 33(2): 87-91.
[6] Zhou Z, Gu B J, Sun H, et al. Molecular phylogenetic analyses of Euphorbiaceae tribe Epiprineae, with the description of a new genus, Tsaiodendron gen. nov., from south-western China[J]. Botanical Journal of the Linnean Society, 2017, 184(2): 167-184.
[7] 孙儒泳. 普通生态学[M]. 北京: 高等教育出版社, 1993: 134.
[8] 马晨晨, 代俊, 肖之强, 等. 极小种群物种云南肉豆蔻的群落结构及其种群现状[J]. 广西植物, 2017, 37(6): 783-790, 782.
[9] 中国科学院昆明植物研究所. 云南种子植物名录: 上册[M]. 昆明: 云南人民出版社, 1984: 887.
[10] Flora of China Editorial Committee. Flora of China: Vol.12[M]. Beijing: Science Press and Missouri Botanical Garden Press, 2007.
[11] IUCN. IUCN Red List Categories and Criteria, Version3.1. 2nd edn[S]. Gland, Switzerland and Cambridge, UK: IUCN, 2012.
