植物-土壤反馈作用对2种引进树种的菌根侵染率和生物量的影响
2018-06-22陈璐璐雷妮娅
陈璐璐,雷妮娅
(北京林业大学 林学院,北京100083)
现代植物生态学视植物与其生存的环境为一个彼此依赖且相互影响的整体,其中,植物-土壤反馈关系最具代表性。植物体扎根土壤并从中获取水肥资源用于生长,直接影响到土壤微生物群系种类组成及其动态变化,此类变化反过来又作用于植物根系,通过影响根系吸收从而作用于植物体的生长发育,形成一个相互影响的反馈作用环[1]。目前,植物-土壤反馈研究主要集中在植物对不同来源土壤微生物的反馈响应,供试树种、试验方法及测定指标均是影响植物-土壤反馈作用结果的重要因素[2-3]。KULMATISKI等[3]对2008年以前的329个反馈试验进行总结和分析,发现反馈作用多以供试种生物量、株高及种子存活率等进行衡量,其中近70%的反馈试验表明土壤微生物对植物为负反馈作用,仅28%为正反馈作用,少量试验为中立反馈即无影响,且试验树种是影响反馈结果的一个重要因素。土壤微生物接种方法是影响植物-土壤反馈结果的另一个重要因素。近年来,科学家们发现不同径级的土壤微生物在反馈体系中的作用差别显著[4-5]。关于不同径级土壤微生物群对植物菌根真菌的影响情况则少见报道。丛枝菌根真菌(arbuscular mycorrhizal fungi,AM)是土壤中对植物生长有重要有益作用的微生物群,促进植物根系对水分、营养元素等的吸收[6],在植物生长、定植乃至群落演替中扮演重要角色[7-8]。目前,反馈试验中关于丛枝菌根真菌的报道不少,肯定了丛枝菌根真菌在植物-土壤反馈作用中具有重要作用[9-11]。研究表明:丛枝菌根真菌促进植物生物量积累,而该种有益作用在不同的反馈体系中未得到表现[9,12],它们在植物-土壤相互作用关系中的响应情况仍需进一步研究。近百年来,中国大量引进复叶槭Acer negundo和美国白蜡Fraxinus americana作为庭院及 “四旁”绿化树种,2个树种在中国生长良好,并在华北地区广泛分布。复叶槭扩散能力强,作为园林树种引进欧洲后被不少地区视为入侵树种,人们认为复叶槭入侵成功源于逃离了本土专性有害土壤微生物的抑制作用,也即天敌释放假说[13-14]。目前,对美国白蜡在不同土壤反馈体系下的响应研究不多,关于它是否发展为入侵物种及其引入机制尚未见报道。本研究以复叶槭和美国白蜡2个引进树种为对象,研究两者对不同径级及不同来源土壤微生物的反馈响应,采用少量接种土壤与基质土壤混合的盆栽育苗方法,针对供试种构建不同的植物-土壤反馈体系。通过比较幼苗在不同植物-土壤反馈体系中感染丛枝菌根真菌和积累生物量的情况,来判定其植物-土壤反馈作用的方向与强度,从而初步确定2个引进树种在中国的扩散机制。
1 材料与方法
1.1 试验材料
供试树种为美国白蜡和复叶槭,2013年底分别从美国Louisiana Forest Seed Company和匈牙利Juhasz&Co.Ltd购进当地原产的成熟种子。于2014年4月以营养草炭土为基质育苗,7月选取相同规格的幼苗移植到口径为14 cm,高16 cm的塑料花盆,进行盆栽试验。育苗和移植过程中所用器具、材料(含种子及幼苗根系)均经过体积分数为75%的乙醇擦拭或浸泡进行表面消毒处理,并以纯净水冲洗后使用。育苗期及幼苗移植后缓苗期采用人工喷洒纯净水进行水分管理。制备接种剂所用到野外土壤,分别从中国林木种子公司大兴苗圃内自留的美国白蜡片林和复叶槭片林林下采集。采集的土壤过2 mm筛分成4份备用。
1.2 盆栽试验设计
1.2.1 不同径级微生物类群下植物-土壤反馈试验 将从野外采集的美国白蜡和复叶槭片林下土壤过2.00 mm筛(保留大型土壤动物及大型真菌等)作为原状土壤处理(field soil,FS2.00,<2.00 mm),过2.00 mm筛并用无菌水浸泡24 h后经快速定量滤纸(0.12 mm)滤出上层澄清液制备土壤浸提液(去除大型动物及大型真菌等),作为土壤浸提液处理(soil wash,SW0.12,<0.12 mm),灭菌土壤为对照(ck),灭菌方法参考文献[15]。其中浸提液处理添加与其他处理等量的灭菌土壤以保持各处理之间土壤养分等的均一性。接种土壤与基质按V(土壤)∶V(基质)=1∶2,充分混合均匀。分别在各处理土壤中移栽前期培育良好的美国白蜡和复叶槭幼苗,10株·处理-1,共60株。
1.2.2 不同来源土壤微生物类群下植物-土壤反馈试验 将从野外采集的美国白蜡和复叶槭片林下土壤分别过2.00 mm筛,与无菌草炭土按V(林下土)∶V(草炭土)=1∶2充分混匀,互相作为同种树下土壤(home soil,HS)及异种树下土壤处理(away soil,AS),即美国白蜡的异种树下土壤处理以复叶槭片林下取得的土壤接种获得,复叶槭异种树下土壤处理同理。10株·处理-1,共计40株。试验采用完全随机区组设计,花盆摆放在透光帐篷内的高脚木架上,每周按电脑临时生成的随机表进行一次重新排位。木架脚用塑料薄膜包覆,包覆高度为15 cm,以防止蚯蚓Pheretima等土壤动物爬上高架进入花盆。缓苗期内浸提液处理喷洒土壤浸提液,其他处理喷洒纯净水;缓苗期过后统一进行常规管护。经过2个生长季后,于2015年9月底将试验苗木带回实验室收集相关数据。
1.3 苗木收获与数据采集
1.3.1 生物量测定 逐株收获苗木,轻轻抖落根系土壤,将基质从根系上剥离,必要时借助水冲。将收获的苗木用自来水冲洗干净后用纸巾吸干表面水分,分叶、茎干、粗根(直径>2 mm)、细根(直径<2 mm)等4个部分分别称取并记录鲜质量,然后各取一定质量的样品于60℃烘箱中烘干至恒量,进行各部分含水率测定,并将鲜质量换算为生物量(干质量)。
1.3.2 菌根结构特征观察 在对细根进行取样烘干处理之前留取少量乳白色根端,切成约1 cm的根段,清洗干净后用甲醛-乙醇-醋酸混合固定液[FAA, V(体积分数为 50%乙醇)∶V(甲醛水)∶V(冰醋酸)=90∶5∶5]固定保存,并加入5 mL甘油长期固定[16],用于丛枝菌根菌侵染率测定样。丛枝菌根菌侵染率测定参考PHILLIPS等[17]的方法。从固定液中取出根段样本并放入质量浓度为10%氢氧化钾溶液中,90℃水浴加热1.5 h至根系透明;洗去碱液后用体积分数为10%的过氧化氢漂白15 min;清洗干净并用0.2 mol·L-1的盐酸酸化1 h;酸化根段用台盼蓝染色15 min后用乳酸酚脱色30 min;最后将脱色处理过的根段清洗干净后压片进行显微镜镜检。随机取幼苗根段10条·株-1,共100条·处理-1进行镜检。镜检采用十字交叉法计数[18],对视野内被十字丝所搭上的菌丝、泡囊和丛枝结构分别进行计数并记录在同一个交叉点下,通过等距移动玻片使每组根段形成不少于150个观察视野,用佳能PC1591对2个树种幼苗的丛枝菌根结构特征进行拍摄记录[19]。泡囊侵染率=(泡囊侵染的交叉点数和/总交叉数)×100%,丛枝侵染率=(丛枝侵染的交叉点数和/总交叉数)×100%。菌丝侵染率=(菌丝侵染的交叉点数和/总交叉数)×100%,总侵染率=(总交叉点-无侵染点数)/总交叉点×100%。
1.4 数据处理与分析
利用Excel 2007对数据进行平均值、标准误等初步处理,生物量和菌根侵染率采用SPSS 17.0进行单因素方差分析(one-way ANOVA),Duncan多重比较及Pearson相关分析。相关图表在SigmaPlot 12.0中完成。
2 结果与分析
2.1 供试种根系菌根结构与类型
2个供试树种的菌根结构组成差异明显。美国白蜡根系菌根为典型的A型(arum-type)丛枝结构,复叶槭的菌根结构为典型的P型(paris-type)。美国白蜡根系菌根菌丝除少量观测胞内菌丝圈(图1 A)外,多为胞间菌丝和根围菌丝,且多为无隔菌丝,少见有隔菌丝;泡囊结构的形态则主要有椭圆形、球形及棒状,偶见不规则状泡囊(图1C);丛枝结构为典型的二叉分支结构(图1D)。复叶槭根系菌根的菌丝则以胞内菌丝和菌丝圈为主,少量观测到胞间菌丝(图2A,B,C);各处理中泡囊均不常见,形状多为椭圆形(图2D);丛枝结构多为消解状态,仅在细胞皮层中形成点状。
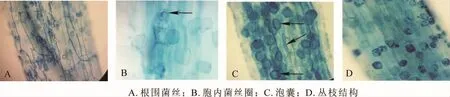
图1 美国白蜡根系丛枝菌根结构(800倍)Figure 1 Arbuscular mycorrhizal fungi structure of Fraxinus americana(800 ×)

图2 复叶槭根系丛枝菌根结构(800倍)Figure 2 Arbuscular mycorrhizal fungi structure of Acer negundo(800 ×)
2.2 不同径级土壤微生物接种处理下供试种反馈响应情况
2.2.1 不同径级土壤微生物接种下2个树种幼苗生物量 由图3可见:美国白蜡在SW0.12处理下总生物量最高,地上部分茎、叶生物量积累均受到促进作用;美国白蜡在FS2.00处理下总生物量最低,地下部分生物量显著低于对照(P<0.05),FS2.00处理抑制了地下部分生物量的积累;美国白蜡在SW0.12处理下各部分生物量显著高于FS2.00处理下各部分生物量(P<0.05)。复叶槭在FS2.00处理下总生物量最高,且该处理下细根、粗根、茎、叶等各部分生物量也是最高,SW0.12与FS2.00处理下复叶槭各部分生物量无显著差异。
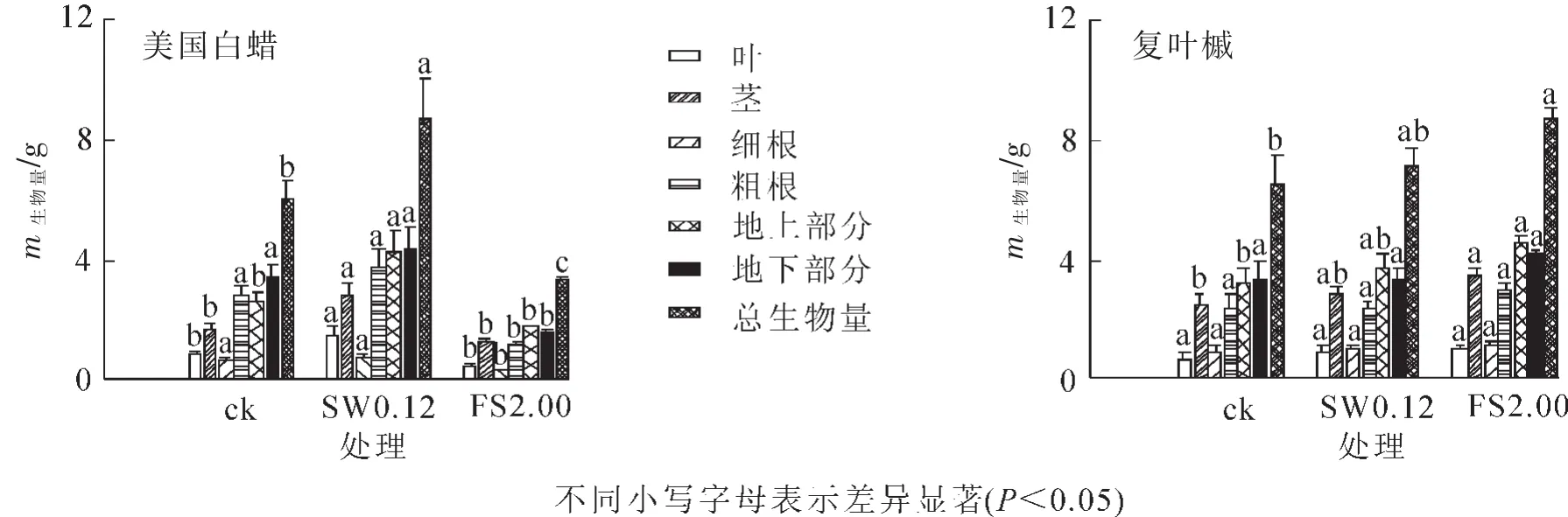
图3 不同径级土壤微生物接种下美国白蜡和复叶槭各部分生物量Figure 3 Effects of different size class of soil biota on the biomass of Fraxinus americana and Acer negundo
2.2.2 不同径级土壤微生物接种下2个树种幼苗菌根侵染 由图4可见:不同土壤处理对美国白蜡菌根各结构发育有影响,丛枝结构定殖率在SW0.12处理下最高,而在FS2.00处理下最低;菌丝侵染率在FS2.00处理下最高,而在SW0.12处理下最低,SW0.12处理抑制美国白蜡菌丝侵染;就总侵染率而言,SW0.12处理下最高,其次是FS2.00处理,对照(ck)处理最低。不同土壤处理对复叶槭各菌根结构发育无影响,菌根总侵染率在FS2.00处理下最高,其次是SW0.12处理,ck处理下最低。

图4 不同径级微生物接种下美国白蜡和复叶槭菌根侵染率Figure 4 Effects of different size class of soil biota on the mycorrhizal colonization of Fraxinus americana and Acer negundo
2.3 不同来源土壤对供试种反馈作用差异
2.3.1 不同来源土壤微生物接种下2个树种生物量 由图5可见,美国白蜡在AS处理下叶、茎、细根、粗根等各部分生物量均显著高于HS处理,分别高出133.2%,75.0%,106.5%,167.7%,总生物量高出量为119.3%(P<0.05)。相反,复叶槭在HS处理下叶、茎、细根、粗根等各部分生物量显著高于AS处理(P<0.05),分别高出为144.1%,95.0%,72.7%,112.9%,HS处理下总生物量比AS处理高出102.2%。2个树种地上部分和地下部分生物量均受AS和HS处理影响显著。

图5 不同土壤来源微生物接种下美国白蜡和复叶槭生物量Figure 5 Effects of soil source on the biomass of Fraxinus americana and Acer negundo
2.3.2 不同土壤来源微生物接种下2个树种菌根侵染率 从图6可见:美国白蜡总侵染率在AS处理下显著高于HS处理(P<0.05),高出81.4%,各菌根结构中,除丛枝结构定殖率在HS和AS之间无差异外,其他结构均为AS处理高于HS处理(P<0.05)。不同土壤处理下复叶槭各菌根侵染率及总侵染率无显著差异(P>0.05)。

图6 不同土壤来源微生物处理下美国白蜡和复叶槭的菌根侵染率Figure 6 Effects of soil source on the mycorrhizal colonization of Fraxinus americana and Acer negundo
2.4 2个树种菌根侵染与生物量相关关系分析
总体来看,各处理下2个树种总生物量与总侵染率无显著相关性,不同处理影响2个树种生物量与各菌根结构的相关关系。在特定植物-土壤反馈体系内,幼苗生物量与菌根侵染率间表现出显著或极显著相关性。表1可见:对照处理下,美国白蜡幼苗叶生物量、地上部分生物量及总生物量均与泡囊侵染率有显著正相关关系(P<0.05);HS处理下美国白蜡叶生物量、茎生物量与泡囊侵染率显著负相关(P<0.05);AS处理下美国白蜡粗根生物量与泡囊侵染率显著负相关(P<0.05)。由表2可见:HS处理下复叶槭幼苗泡囊定殖率与其细根生物量、茎生物量显著正相关(P<0.05),与地上部分生物量和总生物量极显著正相关(P<0.01)。在植物-土壤反馈体系中,2个树种根系泡囊侵染率是一个相对比较敏感的指标。
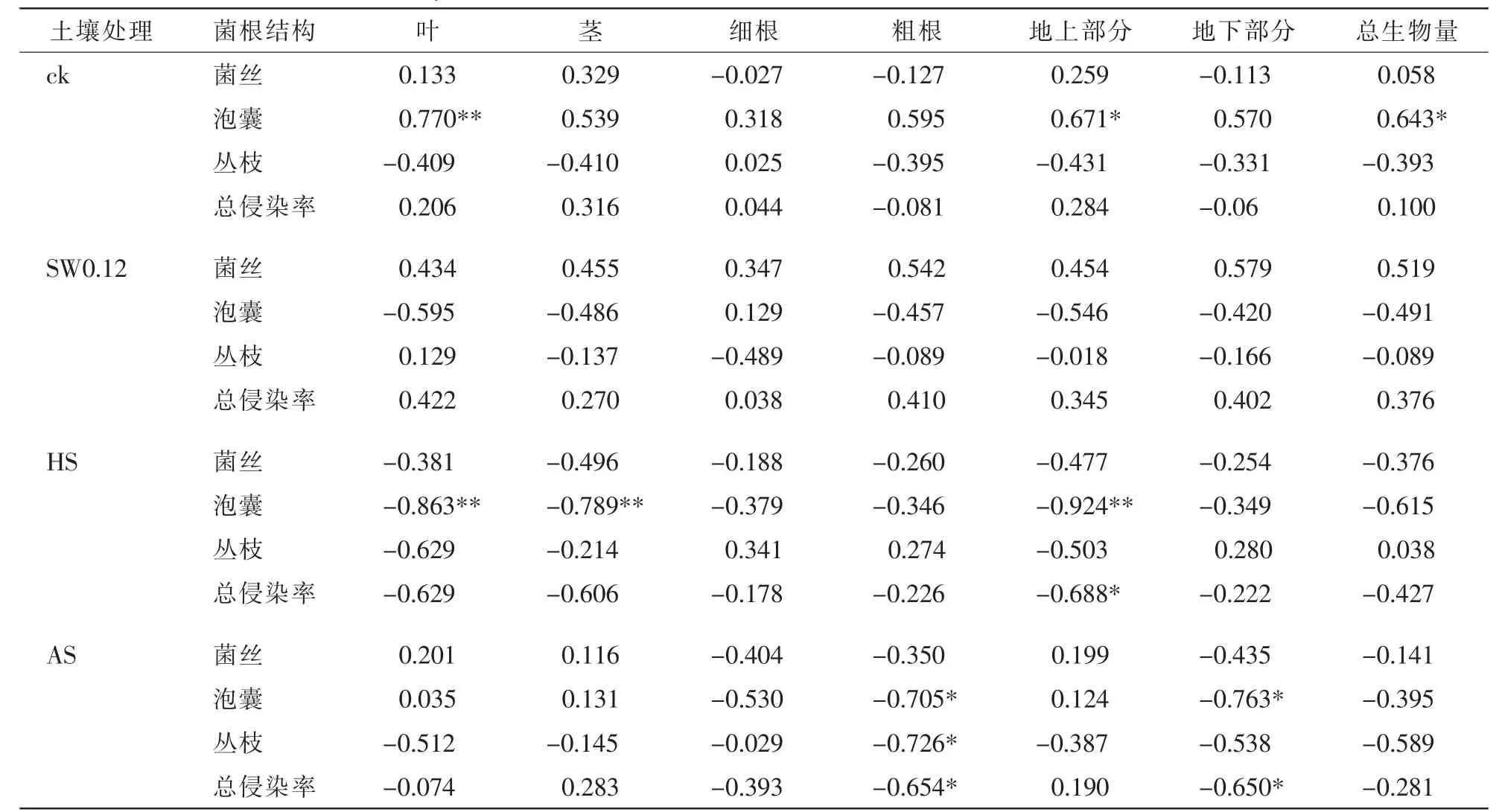
表1 不同处理下美国白蜡各菌根结构侵染率与各部分生物量相关性分析Table 1 Correlation analysis of AMF colonization and biomass of Fraxinus omericana under different treatments

表2 不同处理下复叶槭各菌根结构侵染率与各部分生物量的相关性分析Table 2 Correlation analysis of AMF colonization and biomass of Acer negundo under different treatments
3 讨论
3.1 同树种林下不同径级土壤微生物类群对幼苗的反馈作用
目前,判定植物-土壤反馈作用强度和方向主要通过测定供试种生物量、株高及种子萌发等情况。从原状土处理与对照组的对比来看,美国白蜡林下土壤不利于其幼苗生长,即表现为植物-土壤反馈体系中的负反馈作用;而复叶槭林下土壤则是利于其幼苗生长,表现为正反馈作用。2个树种幼苗的生长均受土壤微生物影响较大,美国白蜡林下土壤存在抑制其同种幼苗生长的专性真菌或病原菌,可能抑制自身的林下更新,能有效地规避同种内部代际竞争,维持群落物种多样性[20-22]。复叶槭林下的土壤正反馈作用则可能有利于其幼苗在林下的更新以及持续生存,从而增加树种在群落中的丰度,也可能增强该物种在某地的入侵能力[3,13-14]。对比不同径级土壤微生物类群(<2.00 mm,<0.12 mm)处理下2种植物生长表现可知,去除大型动物及大型真菌等的土壤浸提液更有利于美国白蜡幼苗生长,表明美国白蜡林下土壤中大型土壤动物抑制幼苗更新及生长,且该类大型动物可能抑制径级相对较小的微生物类群的有益作用;而就复叶槭而言,浸提液处理中(<0.12 mm)幼苗各部分生物量较原状土壤接种处理的略低但无显著差异,表明不同径级土壤微生物类群对复叶槭植物-土壤反馈作用不产生影响。不同的土壤处理方式或土壤接种方式对反馈作用的强度和方向影响较大[4-5],但这种影响也因物种而异。
对照处理下2种幼苗根系侵染率均不为0,可能是前期育苗基质中含有一定量的真菌类群,致使幼苗根系有一定的菌根侵染,用体积分数为75%的乙醇消毒基质并不能抑制其生长或将其消灭。从不同径级土壤微生物类群对2种植物菌根真菌的反馈作用来看,无论是浸提液还是原状土壤接种中微生物类群较对照均有益于2种植物菌根真菌侵染,但这2种土壤处理方式对供试种菌根真菌定殖率影响差异不大。
3.2 不同土壤来源微生物对植物的反馈作用
对比同种树下土壤及异种树下土壤处理下的植物生长表现,美国白蜡受到同种树下土壤微生物类群的抑制作用,而在异种树下土壤中则受到促进作用;复叶槭则受到同种树下土壤微生物类群的促进作用,而在异种树下土壤中则受到抑制作用。NIJJER等[23]进行了与本研究相似的试验,即同种树下土壤较异种树下土壤不利于乌桕Sapium sebiferum幼苗定植和生长,并在进一步的反馈试验中认为同种树下土壤中抑制幼苗生长的是土壤中的真菌。本研究发现美国白蜡在同种树下土壤菌根侵染率及生物量均显著低于异种树下土壤,该种抑制作用是否也来源于同种成年树林下土壤中具有抑制作用的真菌则需要进一步的实验验证。美国白蜡在异种土壤接种中有较高的菌根侵染率及生物量积累,而相关性分析却表明菌根侵染率与生物量无显著相关关系。复叶槭菌根侵染率在2种土壤处理下无显著差异,而生物量同种树下土壤处理显著高于异种树下土壤处理,菌根真菌的有益作用并未在生物量积累中得到显现。不少反馈试验也证实供试种生物量与菌根真菌并无显著相关关系[9,24-25],菌根真菌对植物的促进作用无法与土壤中发挥负反馈作用的微生物类群抗衡,其正反馈作用可能受到抑制[10,23,26]。
美国白蜡植物-土壤反馈关系符合JANZEN-CONNELL模型假说[20,27],即植物-土壤反馈作用抑制成年树林下同种幼苗生长从而维持较高物种多样性;该种植物-土壤反馈模式有利于森林群落维持较高的物种丰富度,从而避免单一物种结构,促进森林群落健康发展。复叶槭植物-土壤反馈关系则符合物种入侵机制中逃离天敌假说[28]:物种引入区域内自然天敌远少于本土区域,从而促进该物种大量定植。REINHART等[13]研究表明:复叶槭在美国本土区域受到同种树下和异种树下土壤负反馈作用,本土区域土传病原菌对其扩散有严重抑制作用。本研究中,复叶槭在中国引种区则受到正反馈作用,表明复叶槭在中国有较强扩散能力和入侵潜力,鉴于复叶槭在北美及欧洲部分地区成功入侵并带来严重后果,建议对其(大苗、幼树、成年树)植物-土壤反馈作用进行跟踪研究,并对其入侵能力进行全面评估。
3.3 菌根结构及类型在植物-土壤反馈体系中的响应
供试种美国白蜡和复叶槭丛枝菌根类型分别为A型和P型。丛枝P型结构较A型结构发育更慢,2种类型菌根结构吸收养分的能力也有差异[29-30]。根据本研究中美国白蜡和复叶槭在不同反馈体系中的响应,推断不同菌根类型可能影响供试种在植物-土壤反馈体系中的表现。目前没有对这一观点进行阐明的研究,需进一步验证。不同的土壤接种方式对美国白蜡菌根各结构的组成有显著影响,其中不同径级土壤微生物及不同来源土壤微生物处理均对美国白蜡泡囊及菌丝结构定殖率有显著影响;而与此不同的,复叶槭在各土壤处理下各菌根结构定殖率无显著差异;不同反馈体系下土壤接种剂对2种植物菌根泡囊与生物量的相关关系有显著影响,在不同反馈体系下泡囊结构表现相对敏感。2个树种表现出明显的营养吸收方式的差异,在不同处理中美国白蜡菌根侵染率可达70%,复叶槭仅45%~55%。同时,在相同土壤接种下复叶槭细根及粗根生物量均高于美国白蜡细根及粗根生物量,表明2个树种对菌根的依赖性确有差异;结合供试种在不同土壤接种中反馈响应差异,推断寄主植物菌根依赖强度在一定程度上可能影响植物与土壤微生物之间的耦合关系[31]。
4 结论
本研究在一定程度上阐明了美国白蜡和复叶槭在中国引种区的反馈模式,并探讨它们的不同反馈机理。得到以下结论:①试验方法影响植物-土壤反馈作用结果。不同的反馈体系的设计影响植物物种生物量及菌根侵染率表现,土壤中大型动物及大型真菌对美国白蜡幼苗生长有重要影响,研究植物-土壤反馈作用机制时应考虑土壤中不同径级土壤微生物类群的功能差异性。②试验植物种类影响植物-土壤反馈作用结果。美国白蜡和复叶槭表现出不同反馈模式,美国白蜡林下土壤抑制其幼苗生长的植物-土壤反馈模式有利于森林群落规避同种内部代际竞争,而复叶槭林下土壤促进其幼苗生长的反馈模式则提示其具有较强的入侵潜能。③不同菌根类型植物反馈作用差异明显,菌根特性是植物-土壤反馈作用评估的重要指标。菌根类型差异导致植物根系吸收养分的方式有很大差异,从而可能引起不同植物在植物-土壤反馈作用中的差异,并且在本研究的反馈作用中,泡囊结构对植物种类差异及反馈体系差异的响应较为敏感。因此,在今后的植物-土壤反馈研究中,与植物互利共生作用的菌根特性应考虑在内。
5 参考文献
[1]BEVER J D,DICKIE I A,FACELLI E,et al.Rooting theories of plant community ecology in microbial interactions[J].Trends Ecol Evol,2010,25(8):468 - 478.
[2]HARRISON K A,BARDGETT R D.Influence of plant species and soil conditions on plant-soil feedback in mixed grassland communities [J].J Ecol,2010,98(2):384 - 395.
[3]KULMATISKI A,BEARD K H,STEVENS J R,et al.Plant-soil feedbacks:a meta-analytical review [J].Ecol Lett,2008,11(9):980 - 992.
[4]BRINKMAN E P,van der PUTTEN W H,BAKKER E J,et al.Plant-soil feedback:experimental approaches,statistical analyses and ecological interpretations [J].J Ecol,2010,98(5):1063 - 1073.
[5]van de VOORDE T F J,van der PUTTEN W H,BEZEMER T M.Soil inoculation method determines the strength of plant-soil interactions [J].Soil Biol Biochem,2012,55(1):1 - 6.
[6]刘永俊.丛枝菌根的生理生态功能[J].西北民族大学学报(自然科学版),2008,29(1):54-59.LIU Yongjun.The physiological and ecological functions of arbuscular mycorrhiza [J].J Northwest Univ Natl Nat Sci,2008,29(1):54 - 59.
[7]梁倩倩,李敏,刘润进,等.全球变化下菌根真菌的作用及其作用机制[J].生态学报,2014,34(21):6039-6048.LIANG Qianqian,LI Min,LIU Runjin,et al.Function and functioning mechanisms of mycorrhizal fungi under global changes [J].Acta Ecol Sin,2014,34(21):6039 - 6048.
[8]BUNN R A,RAMSEY P W,LEKBERG Y.Do native and invasive plants differ in their interactions with arbuscular mycorrhizal fungi?a meta-analysis [J].J Ecol,2015,103(6):1547 - 1556.
[9]YANG Qiang,CARRILLO J,JIN Hongyue,et al.Plant-soil biota interactions of an invasive species in its native and introduced ranges:implications for invasion success [J].Soil Biol Biochem,2013,65(5):78 - 85.
[10]CALLAWAY R M,BEDMAR E J,REINHART K O,et al.Effects of soil biota from different ranges on Robinia invasion:acquiring mutualists and escaping pathogens [J].Ecology,2011,92(5):1027 - 1035.
[11]CALLAWAY R M,CIPOLLINI D,BARTO K,et al.Novel weapons:invasive plant suppresses fungal mutualists in America but not in its native Europe [J].Ecology,2008,89(4):1043 - 1055.
[12]KLIRONOMOS J N.Feedback with soil biota contributes to plant rarity and invasiveness in communities [J].Nature,2002,417(6884):67 - 70.
[13]REINHART K O,CALLAWAY R M.Soil biota facilitate exotic Acer invasions in Europe and North America [J].Ecol Appl,2004,14(6):1737 - 1745.
[14]REINHART K O,TYTGAT T,van der PUTTEN W H,et al.Virulence of soil-borne pathogens and invasion by Prunus serotina [J].New Phytol,2010,186(2):484 - 495.
[15]侯永侠,周宝利,吴晓玲,等.土壤灭菌对辣椒抗连作障碍效果[J].生态学杂志,2006,25(3):340-342.HOU Yongxia,ZHOU Baoli,WU Xiaoling,et al.Effects of soil sterilization on preventing continuous pepper cropping obstacles [J].Chin J Ecol,2006,25(3):340 - 342.
[16]张春英.云锦杜鹃菌根及其菌根真菌多样性研究[D].北京:北京林业大学,2008.ZHANG Chunying.Diversity of Ericoid Mycorrhiza and the Related Fungi Isolated from Hair Roots of Rhododendron fortune L.[D].Beijing: Beijing Forestry University,2008.
[17]PHILLIPS J M, HAYMAN D S.Improved procedures for clearing roots and staining parasitic and vesicular-arbusular mycorrhizal fungi for rapid assessment of infection [J].Trans Bri Mycol Soc,1970,55(1):158 - 161.
[18]MCGONIGLE T P,MILLER M H,EVANS D G,et al.A new method which gives an objective measure of colonization of roots by vesicular-arbuscular mycorrhizal fungi[J].New Phytol,1990,115(3):495 - 501.
[19]曾端香,袁涛,王莲英.AM真菌接种剂与栽培基质对牡丹容器苗丛枝菌根侵染的影响[J].中国农学通报,2011,27(10):108-112.ZENG Duanxiang, YUAN Tao, WANG Lianying.The effects of AM fungus and culture substrates on colonization of arbuscular mycorrhizal for tree peony container seedling [J].Chin Agric Sci Bull,2011,27(10):108-112.
[20]LIU Yu,YU Shixiao,XIE Zhiping,et al.Analysis of a negative plant-soil feedback in a subtropical monsoon forest[J].J Ecol,2012,100(4):1019 - 1028.
[21]HOVATTER S,BLACKWOOD C B,CASE A L.Conspecific plant-soil feedback scales with population size in Lobelia siphilitica (Lobeliaceae) [J].Oecologia,2013,173(4):1295 - 1307.
[22]肖博,周文,刘万学,等.紫茎泽兰入侵地土壤微生物对紫茎泽兰和本地植物的反馈[J].中国农业科技导报,2014,16(4):151 - 158.XIAO Bo,ZHOU Wen,LIU Wanxue,et al.Feedback of Ageratia adenophora soil microbe on A.adenophora and native plants [J].J Agric Sci Technol,2014,16(4):151 - 158.
[23]NIJJER S,ROGERS W E,SIEMANN E.Negative plant-soil feedbacks may limit persistence of an invasive tree due to rapid accumulation of soil pathogens [J].Proc R Soc B,2007,274(1625):2621 - 2627.
[24]NIJJER S,ROGERS W E,SIEMANN E.The effect of mycorrhizal inoculum on the growth of five native tree species and the invasive Chinese tallow tree (Sapium sebiferum) [J].Tex J Sci,2004,56(4):357 - 368.
[25]NIJJER S,ROGERS W E,LEE C T A,et al.The effects of soil biota and fertilization on the success of Sapium sebiferum [J].Appl Soil Ecol,2008,38(1):1 - 11.
[26]NIJJER S.Understanding Belowground Community Regulation in an Invaded Forest [D].Houston:Rice University,2006.
[27]McCARTHY-NEUMANN S,KOBE R K.Conspecific and heterospecific plant-soil feedbacks influence survivorship and growth of temperate tree seedlings [J].J Ecol,2010,98(2):408 - 418.
[28]MITCHELL C E,POWER A G.Release of invasive plants from fungal and viral pathogens [J].Nature,2003,421(6923):625 - 627.
[29]刘润进,陈应龙.菌物学[M].北京:科学出版社,2007.
[30]柏艳芳,郭绍霞,李敏.入侵植物与丛枝菌根真菌的相互作用[J].应用生态学报,2011,22(9):2457-2463.BAI Yanfang,GUO Shaoxia,LI Min.Interactions between invasive plants and arbuscular mycorrhizal fungi:a review[J].Chin J Appl Ecol,2011,22(9):2457 - 2463.
[31]李立青,张明生,梁作盼,等.丛枝菌根真菌促进入侵植物紫茎泽兰的生长和本地植物竞争效应[J].生态学杂志, 2016,35(1):79 - 86.LI Liqing,ZHANG Mingsheng,LIANG Zuopan,et al.Arbuscular mycorrhizal fungi enhance invasive plant,Ageratina adenophora growth and competition with native plants [J].Chin J Ecol,2016,35(1):79 - 86.
