水稻种子休眠特性与耐藏性的关系
2018-06-22陈志超黄婷婷吴洪恺陈璋琦陶若芙
陈志超,黄婷婷,王 洋,吴洪恺,陈璋琦,陶若芙
(1.浙江农林大学 农业与食品科学学院 浙江省农产品品质改良技术研究重点实验室,浙江 杭州311300;2.浙江省舟山市定海区农林与海洋渔业局,浙江 舟山 316000)
水稻Oryza sativa是中国最重要的粮食作物之一。中国因储藏问题引起的粮食损失达350亿kg·a-1,因此,如何选育耐藏性好的种子一直是种子科学研究的重点。在杂交水稻制种过程中,通过选育具有强休眠性的保持系和不育系可以解决杂交稻制种穗发芽的危害,延长种子的储藏时间。侯雅杰[1]认为,休眠种子能通过长时间储藏而保持活力,适应性较强。但也有学者认为休眠性强的种子可能会影响种子的萌发,并不一定更耐储藏[2]。种子休眠性和耐藏性之间的关系研究较少,仍不明确。种子休眠性与种子活力密切相关。近年来,已有部分学者对种子休眠性与种子活力的关系进行了研究[3-6],普遍认为高活力种子耐藏性好,通过测定种子活力水平即能反映种子的耐藏性好坏。人工加速老化试验是研究种子老化过程中活力变化和种子耐藏性的最常见的方法之一。董国军等[7]研究发现,水稻人工老化方法能较好地模拟种子的自然老化过程。本研究采用40℃,100%相对湿度(RH)高温高湿人工加速老化法(AA)[8],对2个具有休眠特性差异的籼稻Oryza sativa subsp.sativa品种4K58(Ⅱ-32B休眠)和4K59(Ⅱ-32B)及其各自与不育系(Ⅱ-32A)的杂交F2代种子(C178,C179)进行人工加速老化,获得不同老化天数的水稻种子,通过测定与种子活力相关指标,探讨水稻种子休眠特性和耐藏性的关系,以期为选育耐藏性强的水稻品种提供理论依据。
1 材料与方法
1.1 试验材料
供试材料为籼稻品种4K58(Ⅱ-32B休眠),4K59(Ⅱ-32B),C178和C179的种子。4K58种子具有休眠性;C178和C179分别是4K58和4K59与不育系(Ⅱ-32A)杂交获得的F2代种子。
采用人工加速老化方法对供试材料进行处理:相对湿度100%,温度40℃,老化3,6,9 d。老化结束后将种子放在室温下晾干24 h,用纸袋包装,4℃保存备用。以未老化种子为对照(ck)。
1.2 试验方法
1.2.1 种子电导率测定 参照CAO等[9]方法,分别取不同老化天数处理的种子各50粒,设置重复试验3次·组-1,35℃烘2 h,取出冷却后称量;用蒸馏水冲洗3次,去除表面水分后置于100 mL烧杯中,加蒸馏水100 mL,保鲜膜封口后,静置于25℃恒温培养箱24 h。用电导仪测定种子浸出液电导率。电导率(μS·cm-1·g-1)=(样品值-对照值)/样品种子质量(g)。
1.2.2 种子单核苷酸浸出液质量浓度测定 分别取经不同老化天数处理的种子各1粒,设置重复试验10次·组-1;每粒种子置于装有10 mL蒸馏水的20 mL试管中,将试管置于25℃恒温培养箱中24 h。取3 mL 浸出液测定其在 260 nm 下的吸光度 D(260)。 核苷酸 D(260)=40 mg·L-1[10]。
1.2.3 种子发芽试验 种子经质量分数为0.1%的次氯酸钠溶液消毒15 min后,在垫有2层湿润发芽纸的发芽盒中于20~30℃变温条件发芽,高温8 h,低温16 h,高温时段有光照,光合光子照度为250 μmol·m-2·s-1。3次重复,种子100粒·重复-1。每天记录发芽种子数,第5天计算发芽势, 第14天计算发芽率。 参照 ZHANG 等[11]的方法, 计算发芽指数(gi)=∑(nt/td), 平均发芽时间(tg)=∑(nt×td)/∑nt。 其中:nt为第t天的发芽种子数,td为发芽日数。发芽结束后,随机取幼苗10株·盒-1,分别测量苗高和鲜质量。
1.2.4 统计分析 所得数据用SAS进行统计分析,多重比较采用最小显著差(LSD)法,显著性水平为P<0.05, 百分率数据在分析前进行反正弦转换, y=arcsin [sqrt(x/l00)]。
2 结果
2.1 不同老化天数处理对种子电导率的影响
未经老化处理的种子(ck)中,C179的电导率值显著高于4K59和C178;4K58显著高于4K59,约比4K59高18.8%(图1)。老化3 d后,4个水稻品种的电导率值无显著差异;老化6 d后,4K58的电导率值显著高于C179;老化9 d后,C178和C179的电导率值显著高于4K58和4K59;C178和C179之间,4K58和4K59之间均无显著差异。随老化时间的延长,4个品种的种子电导率值均呈增加的趋势;其中4个品种老化9 d的种子电导率值均显著高于老化6 d,增幅达29.0%~71.1%;4K58和4K59老化6 d的电导率值显著高于老化3 d;4K59和C178老化3 d的电导率值显著高于ck。
2.2 不同老化天数处理对种子单核苷酸浸出液质量浓度的影响
未老化的C178种子核苷酸浸出液质量浓度显著高于其他3个品种(图2),而3个品种间无显著差异。老化3 d后,4K58和C178的种子核苷酸浸出液质量浓度分别显著高于4K59和C179;4K58和C178之间,4K59和C179之间均无显著差异。老化3 d后4K58和4K59的种子核苷酸浸出液分别比对照增加了52.3%和3.6%,两者相差近14.5倍。老化6 d后,4K59的核苷酸浸出液质量浓度显著低于其余3个品种;C179显著低于4K58和C178;4K58和C178之间无显著差异。老化9 d后,4K59的核苷酸浸出液质量浓度显著低于其他3个品种;4K58显著高于其他3个品种;C178和C179之间无显著差异。随老化时间的延长,4个品种的种子核苷酸浸出液质量浓度均呈增加的趋势;4K58,4K59和C179老化9 d的种子核苷酸浸出液显著高于老化6 d,4个品种老化6 d的核苷酸浸出液与老化3 d无显著差异;老化3 d的4K58种子核苷酸浸出液质量浓度显著高于ck。

图1 不同水稻品种老化处理后种子的电导率值Figure 1 Conductivity of accelerated aging seeds of four rice cultivars
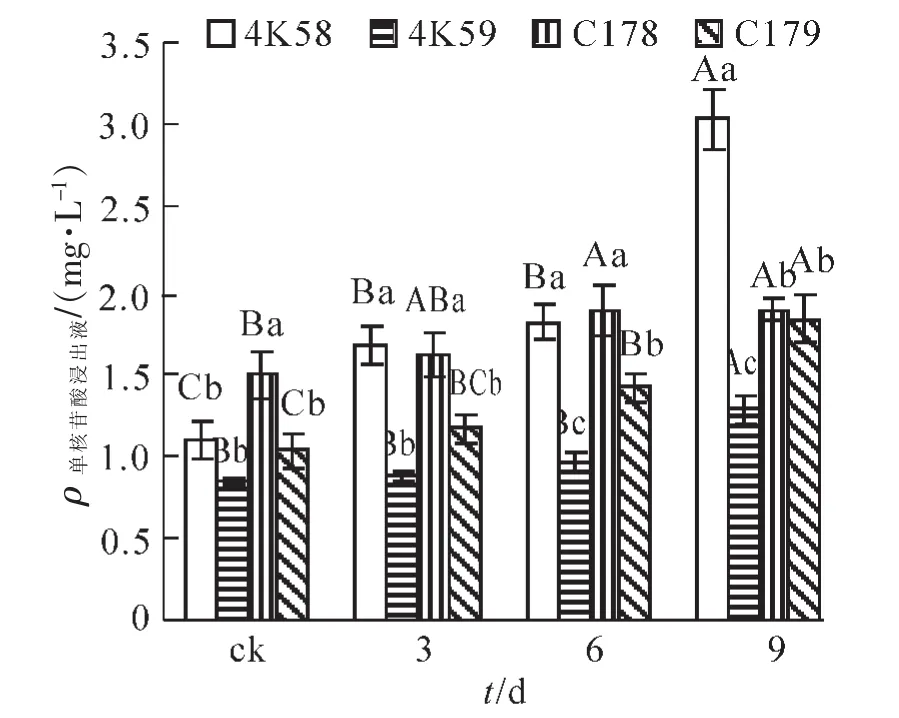
图2 不同水稻品种老化处理后种子核苷酸浸出液质量浓度比较Figure 2 Single nucleotide extract content of accelerated aging seeds of four rice cultivars
2.3 不同老化天数处理对种子萌发的影响
2.3.1 发芽势 由表1可知:4个品种未经老化处理(ck)的种子发芽势均无显著差异。老化3 d和6 d后,4K58种子的发芽势显著低于其余3个品种,其他3个品种均无显著差异;老化9 d后,C179种子的发芽势显著高于其他3个品种,4K58的发芽势显著低于其他3个品种,4K59和C178之间无显著差异。与未老化种子相比,老化9 d后4K58的发芽势下降幅度约为4K59的2.1倍,C178约为C179的2.5倍。随老化时间的延长,4个品种的种子发芽势呈下降趋势,但4个品种老化3 d的发芽势与ck无显著差异。4K58老化9 d的发芽势显著低于老化6 d,老化6 d的发芽势显著低于老化3 d;其他3个品种老化9 d的发芽势显著低于老化6 d,老化6 d的发芽势与老化3 d无显著差异。
2.3.2 发芽率 未经老化处理的4个品种种子发芽率均无显著差异(表2)。老化3 d后,C178种子的发芽率显著低于其余3个品种,而3个品种之间均无显著差异;老化6 d后,C178的发芽率显著低于C179,与4K58和4K59无显著差异;老化9 d后,4K58的发芽率显著低于C179,与4K59和C178无显著差异。与未老化种子相比,老化9 d后4K58的发芽率下降幅度约为4K59的1.4倍,C178约为C179的3.0倍。4个品种的未老化种子、老化3 d和老化6 d种子的发芽率之间均无显著差异;老化9 d后,4K58,4K59和C178种子的发芽率均分别显著低于老化6 d的发芽率。

表1 不同水稻品种老化处理后种子发芽势比较Table 1 Germination energy of accelerated aging seeds of four rice cultivars
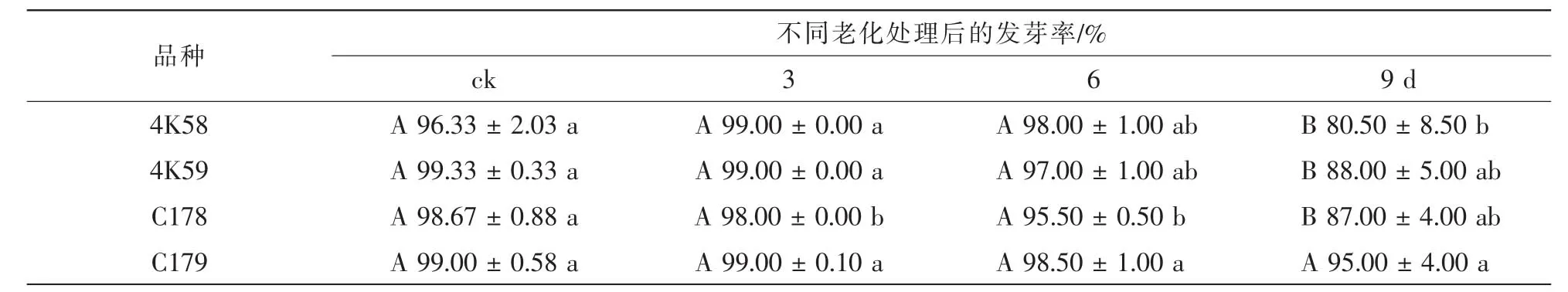
表2 不同水稻品种老化处理后种子发芽率比较Table 2 Germination percentage of accelerated aging seeds of four rice cultivars
2.3.2 发芽指数 从表3可知:未经老化处理的4个品种种子发芽指数均无显著差异。老化3 d后,C179种子的发芽指数显著高于4K58和4K59,与C178无显著差异。老化6 d后,4K59和C179的发芽指数均显著高于4K58和C178;4K59和C179之间,4K58和C178之间均无显著差异。老化9 d后,C179的发芽指数显著高于C178;C178显著高于4K59;4K59显著高于4K58。随老化时间的延长,4个品种种子的发芽指数均呈显著下降趋势,即4个品种的未老化种子发芽指数显著低于老化3 d;老化3 d的发芽指数显著低于老化6 d;老化6 d显著低于老化9 d。与未老化种子相比,老化9 d后4K58的发芽指数下降幅度约为4K59的1.1倍,C178约为C179的1.1倍。

表3 不同水稻品种老化处理后种子发芽指数比较Table 3 Germination index of accelerated aging seeds of four rice cultivars
2.3.3 平均发芽时间 4个品种的未老化种子平均发芽时间无显著差异(表4)。老化3 d后,4个品种的种子平均发芽时间也无显著差异。老化6 d后,4K58的平均发芽时间显著高于C178;C178显著高于4K59;4K59显著高于C179。与未老化种子相比,老化6 d后4K58的平均发芽时间上升幅度约为4K59的1.5倍,C178约为C179的1.4倍。老化9 d后,4K58和4K59的种子平均发芽时间均显著高于C178和C179;4K58和4K59之间,C178和C179之间均无显著差异。随老化时间的延长,4个品种的种子平均发芽时间均呈增加的趋势;其中4个品种老化9 d的种子平均发芽时间显著高于老化6 d;老化6 d显著高于老化3 d;C178和C179老化3 d的种子平均发芽时间显著高于未老化处理的种子。

表4 不同水稻品种老化天数处理后种子平均发芽时间比较Table 4 Mean germination time of accelerated aging seeds of four rice cultivars
2.4 不同老化天数处理对幼苗生长的影响
2.4.1 苗高 种子发芽试验结束后,分别测定了幼苗的苗高和鲜质量。结果显示:未老化和老化9 d的C179幼苗苗高显著高于其余3个品种(图3);C178的幼苗苗高显著高于4K58和4K59;4K58和4K59之间无显著差异。老化3和6 d后,C178和C179的苗高均显著高于4K58和4K59;C178和C179之间,4K58和4K59之间均无显著差异。随老化时间的延长,4个品种的幼苗苗高均呈下降趋势。其中4K58和C178老化9 d的幼苗苗高显著低于未老化幼苗,而老化6 d,3 d和ck之间均无显著差异。4K59的幼苗苗高随老化处理时间延长下降较明显,老化9 d的苗高显著低于老化6 d,老化6 d显著低于老化3 d和ck。未老化的C179幼苗苗高显著高于老化3,6和9 d;后三者间无显著差异。
2.4.2 鲜质量 未老化处理的种子中,C179品种的幼苗鲜质量显著高于4K58,与4K59和C178无显著差异(图4)。老化3 d后,C179的幼苗鲜质量显著高于4K59和4K58;C178的鲜质量显著高于4K58;4K58和4K59之间无显著差异。老化6 d和9 d后,C179的幼苗鲜质量显著高于其他3个品种,而3个品种之间均无显著差异。随老化时间的延长,4个品种的幼苗鲜质量均呈下降趋势,但4K58和C179品种不同老化处理后的幼苗鲜质量均无显著差异。C178老化9 d的幼苗鲜质量显著低于其他3个处理,而3个处理之间均无显著差异。4K59未老化处理的幼苗鲜质量显著高于老化3,6和9 d,后三者之间均无显著差异。
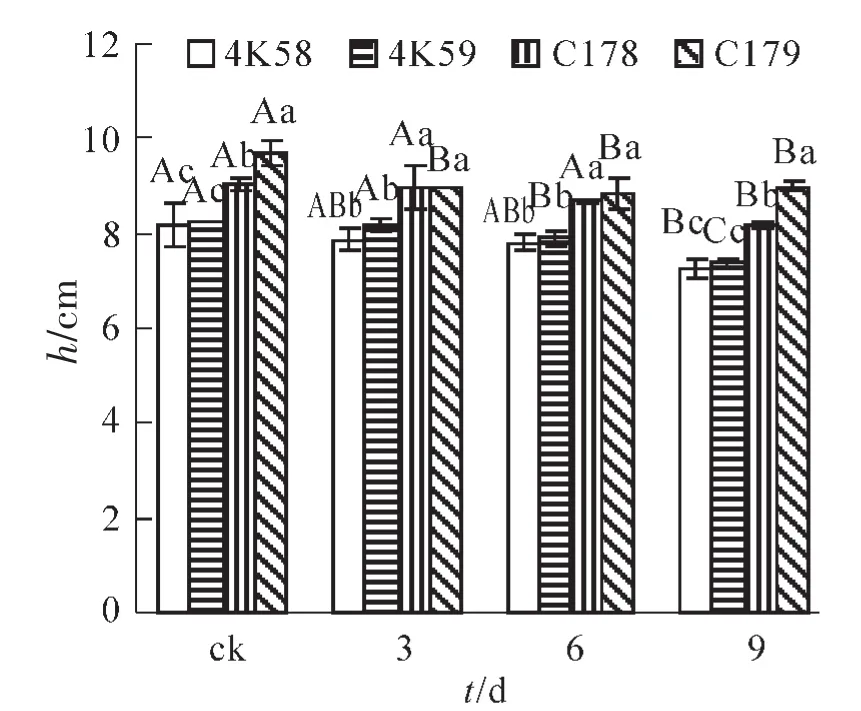
图3 不同水稻品种老化处理后发芽14 d的幼苗苗高Figure 3 Shoot lengths of accelerated aging seeds of four rice cultivars

图4 不同水稻品种老化处理后发芽14 d的幼苗鲜质量Figure 4 Fresh weights of accelerated aging seeds of four rice cultivars
3 讨论
种子休眠是指有生活力的种子在适宜的萌发条件下不能萌发的现象,是植物生长发育过程中的一个暂停生长的阶段[12]。种子休眠是影响种子耐藏性的原因之一。一般认为,休眠种子具备更高的抵抗能力,通过长时间储藏仍能保持活力。然而,关于种子休眠性和耐藏性之间的关系研究仍旧较少,且在不同的物种研究中存在着相反的结论。
人工加速老化是研究种子耐藏性常用的方法。人工加速老化的原理是在人工创造的条件下加速种子的劣变,使种子活力出现差异,通过测定种子活力相关指标的变化来判断种子的耐藏性[13]。由于能在短时间内获得不同老化程度的种子,因而人工加速老化广泛应用于种子耐藏性研究。
细胞膜是维持细胞内环境稳态的一道屏障,细胞膜系统的生理生化变化与种子活力密切相关。种子老化后,细胞膜功能受损,膜透性增加,大量可溶性营养物质及生理活性物质外渗,电导率增大[14]。本研究发现:随老化时间延长,4个品种的种子电导率值均呈增加趋势,说明种子老化时间越长,劣变越严重,活力越低,这与SWAMY等[15]的结果一致。此外,未老化的4K58的电导率值显著高于4K59;老化后4K58的电导率值也高于4K59,表明老化后4K58的种子活力比4K59低,耐藏性更差。
核苷酸浸出液质量浓度的高低反映细胞膜的裂变程度,核苷酸浸出液质量浓度越高,说明种子细胞膜裂变越严重,活力越低[16]。本研究发现,随老化时间的延长,4个品种的核苷酸浸出液质量浓度均呈增加趋势,也说明种子活力随老化程度加深而逐渐降低。未老化处理的4K58和4K59种子核苷酸浸出液无显著差异;但老化处理(3,6,9 d)后,4K58的核苷酸浸出液均显著高于4K59,说明老化后4K58种子活力比4K59更低,不耐储藏。此外,C178种子的核苷酸浸出液质量浓度也显著低于C179,似乎表明这种储藏特性可遗传。
张瑛等[13]指出,通常可采用发芽率、发芽势、发芽指数、平均发芽时间等指标作为判断种子耐藏性的标准。本研究发现,随老化时间的延长,4个品种种子的发芽势、发芽率、发芽指数均逐渐下降,平均发芽时间逐渐增加,这与陈良碧[17]的研究结果一致。未经老化处理的种子,4个品种的发芽势、发芽率、发芽指数和平均发芽时间等均无显著差异。随着老化程度加深,4个品种的发芽指标逐渐出现显著差异。例如老化9 d后,4K58的发芽势显著低于4K59,C178也显著低于C179;老化6 d后,C178的发芽率显著低于C179;老化6和9 d后,4K58和C178种子的发芽指数均分别显著低于4K59和C179;老化6 d后,4K58和C178种子的平均发芽时间均分别显著高于4K59和C179。以上这些结果均表明,经老化后4K58的种子活力比4K59更低,说明4K58的储藏性更差,且其杂交后代也保留了这种特性。同时,本研究也发现:4K58和4K59的杂交F2种子(C178和C179)活力均比4K58和4K59更高,说明这些种子更耐储藏。
种子发芽结束后的幼苗生长指标表明,随老化程度加深,幼苗的苗高和鲜质量均逐渐下降,说明种子活力逐渐下降。所有处理的4K58幼苗苗高和鲜质量均与4K59无显著差异;老化9 d后的C178幼苗苗高和老化6和9 d的幼苗鲜质量均显著低于C179。这些结果说明幼苗的生长指标不能明显反映4K58的储藏性比4K59更差,但两者的杂交F2种子耐藏性存在差异。
综上所述,经人工加速老化处理后,具有休眠特性的4K58种子由于其电导率值和核苷酸浸出液质量浓度均高于无休眠特性的4K59种子;而其发芽指标均低于4K59,推测具有休眠特性的水稻种子更不耐储藏。同时,4K58与不育系的杂交F2种子的耐藏性也低于4K59与不育系的杂交F2种子,表明这种储藏特性可遗传至下代。由于本研究采用的是人工加速老化方法来测定种子的耐藏性,并不能完全反映自然储藏过程中的变化,且只选用了2个供试材料。因此,有关种子休眠特性与耐藏性关系研究还需在更多类型的植物种子以及自然老化的种子材料中进行。
4 参考文献
[1]侯雅杰.种子休眠与耐藏关系探析[J].现代农业科技,2015(23):51.HOU Yajie.Study on relationship between seed dormancy and storage tolerance [J].Mod Agric Sci Technol,2015(23):51.
[2]鱼小军,徐长林,王芳,等.草玉梅种子休眠原因及解除休眠方法[J].生态学杂志,2014,33(1):65-70.YU Xiaojun,XU Changlin,WANG Fang,et al.The cause of Anemone rivularis seed dormancy and the methods for breaking dormancy [J].Chin J Ecol,2004,33(1):27 - 29.
[3]李振华,王建华.种子活力与萌发的生理与分子机制研究进展[J].中国农业科学,2015,48(4):646-660.LI Zhenhua,WANG Jianhua.Advances in research of physiological and molecular mechanism in seed vigor and germination [J].Sci Agric Sin,2015,48(4):646 - 660.
[4]徐保钦,陆作楣.籼稻强休眠保持系Ⅱ 112B与双亲休眠性的比较[J].南京农业大学学报,2008,31(4):8-12.XU Baoqin,LU Zuomei.Comparison on seed dormancy between high dormancy maintainer lineⅡ112B and its parents in indica rice (Oryza sativa L.) [J].J Nanjing Agric Univ,2008,31(4):8 - 12.
[5]李金华,王丰,廖亦龙,等.水稻种子活力的生理生化及遗传研究[J].分子植物育种,2009,7(4):772-777.LIN Jinhua,WANG Feng,LIAO Yilong,et al.Adcance on seed vigor physiological-biochemical and genetic mechanisms in rice [J].Mol Plant Breed,2009,7(4):772 - 777.
[6]兰海,冷亦峰,周树峰,等.强休眠玉米种子休眠前后的蛋白差异表达[J].植物遗传资源学报,2015,16(1): 23 - 28.LAN Hai,LENG Yifeng,ZHOU Shufeng,et al.Proteomic analysis of storage substances during after-ripening of dormant seeds with dry ripening process in maize inbred line 08-641 [J].J Plant Genet Resour,2015,16(1):23 - 28.
[7]董国军,胡兴明,曾大力,等.水稻种子人工老化和自然老化的比较研究[J].浙江农业科学,2004(1):27-29.DONG Guojun,HU Xingming,ZENG Dali,et al.Research on comparison between artificial aging and natural aging of rice seed [J].J Zhejiang Agric Sci,2004(1):27 - 29.
[8]DELOUCHE J C,BASKIN C C.Accelerated aging techniques for predicting the relative storability of seeds lots [J].Seed Sci Technol,1973,1(2):427 - 452.
[9]CAO Dongdong,HU Jin,GAO Canhong,et al.Chilling tolerance of maize (Zea mays L.) can be improved by seed soaking in putrescine [J].Seed Sci Technol,2008,36(1):191 - 197.
[10]DESWAL D P,SHEORAN I S.A simple method for seed leakage measurement:applicable to single seeds of any size[J].Seed Sci Technol,1993,21(1):179 - 185.
[11]ZHANG Sheng,HU Jin,ZHANG Yin,et al.Seed priming with brassinolide improves lucerne (Medicago sativa L.)seed germination and seedling growth in relation to physiological changes under salinity stress [J].Austr J Agric Res,2007,58(8):811 - 815.
[12]XU Shengchun,LI Yongping,HU Jin,et al.Responses of antioxidant enzymes to chilling stress in tobacco seedlings[J].Agric Sci China,2010,9(11):1594 - 1601.
[13]张瑛,滕斌,吴敬德,等.水稻种子高温高湿人工加速老化试验方法研究[J].中国粮油学报,2010,25(10): 8 - 13.ZHANG Ying,TENG Bin,WU Jingde,et al.Study on accelerated aging test of rice seeds with high temperature and humidity [J].J Chin Cereal Oil Ass,2010,25(10):8 - 13.
[14]胡晋.种子学[M].北京:中国农业出版社,2014.
[15]SWAMY Y M I, SOWBHAGYA C M, BHATTACHARYA K R.Changes in the physicochemical properties of rice with aging [J].J Sci Food Agric,1978,29(7):627 - 639.
[16]HAMPTON J G,LEEKS C R F,McKENZIE B A.Conductivity as a vigour test for Brassica species [J].Seed Sci Technol,2009,37(1):214 - 221.
[17]陈良碧.杂交水稻种子生理特点及耐贮藏性的研究[J].种子,1994(4):19-24.CHEN Liangbi.Research on physiological characteristics and storage tolerant of hybrid rice [J].Seed,1994(4):19-24.
