绦柳雌株花芽分化1)
2018-06-15胡雨逸周元超王鲁北袁涛
胡雨逸 周元超 王鲁北 袁涛
(临沂市园林科学研究院,临沂,276000) (北京林业大学)
绦柳(Salixmatsudanaf.pendula)为杨柳科柳属落叶乔木,小枝黄色,光滑柔软且下垂,状若丝绦,常被误认为垂柳。由于生长速度快、适应性强、分布区域广,是我国北方地区重要的园林绿化、生态修复树种,广泛应用于各类城乡绿地绿化当中[1],栽植数量巨大。胡有宁[2]在调查研究城市森林系统时发现,绦柳已成为延安市除槐以外,栽植数量最多的行道树树种,占比高达14.81%。陈崇[3]、李想等[4]、秦仲[5]、赵松婷等[6]更是将其作为北京城市园林绿化代表树种开展相关应用性研究。
随着柳树的大量运用,柳絮的危害也日益凸显。目前研究出的飞絮解决方法主要有喷水降絮、更新雌株、阻断授粉、柳树高位嫁接、疏除雌花序、激素调控雌花芽分化等[7-14]。但这些方法成本高、工作量大、效率低等,一直没有有效地解决绦柳飞絮的问题。王建红等[13]提出在成花决定时间前使用“抑花一号”注射树干可有效防止次年飞絮,之后则无效果。因此,必须明确绦柳雌株花芽分化过程,准确确定成花决定时间,才能更好地防治飞絮。
目前,有关花芽分化的研究多集中在花原基外观形状描述、花芽分化所需的内源激素和营养物质以及特征基因检测方面[15-19]。前两者难以准确判定成花决定时间,后者则因技术难度大、成本高,难以广泛应用。王建红等[13]在成花决定时间前应分阶段注射“抑花一号”,确定了毛白杨的成花决定时间,但未能明确该状态下花芽的外部形态及其与花芽内部分化过程之间的相关性,从而也无法有效地从外观形态判定是否进入成花决定态。另外,“抑花一号”注射法虽然可以准确判定出杨柳属的成花决定时间,但是由于定期切片观察花芽分化所需期限较长,借助芽的形态特征判定更为高效。
本文应用石蜡切片法观察了注射“抑花一号”和未注射的绦柳雌株的花芽分化过程,观察花芽分化前后花芽的外部形态特征和分化过程中的内部结构的变化规律,为有效控制飞絮技术的推广提供理论依据。
1 材料与方法
10年生以上株龄绦柳雌株,生长一致,胸径10~40 cm,株高5~15 m,位于临沂市滨河景区北城新区北京路与滨河西路交汇处东南侧区段。
样本采集:自2016年4月26日至7月25日,选取生长状况和株龄一致的绦柳雌株,每5 d注射一次“抑花一号”。“抑花一号”为粉末状,微溶于水。每克药剂需先完全溶于10 mL 95%酒精,再加水40 mL稀释。然后选用与注射机匹配的钻头(1 cm),用电钻在树干胸径部位垂直钻孔,20 cm胸径以下打孔2个,20~30 cm胸径打孔3个,30~40 cm胸径打孔4个。最后用6HZ-1015型高压树木注射器,按1.75 mL·cm-1胸径药量平均注射到各注射孔中,每个处理设3个重复(3株树),每次采集3株树冠中部四个朝向12根50~100 cm长度的当年生健壮枝条的侧芽100~200个,共计3 349个。
同时选取与上文生长状况和株龄一致的绦柳雌株,不注射“抑花一号”,每5 d采样1次。每次采集5株树冠中部四个朝向12根50~100 cm长度的当年生健壮枝条的侧芽100~200个,共计3 287个,做对照。
花芽外观形态观察:样本采集后用OlympusSZX7型体视显微镜观察并拍照记录,利用CellSens Entry软件测量芽的长宽值,之后放入FAA固定液中固定。
花芽分化过程观察:样本固定2 d后剥芽处理初步观察,统计不同形态分化时期的数量,保存代表性芽体。7 d后脱水包埋,MicroHM315R型轮转式切片机切片,切片厚度10~12 μm,番红、固绿双重染色,中性树胶封片,OlympusBX53型荧光显微镜观察并照相。石蜡切片制作方法参考贺冰等[20]的方法,番红、固绿染色方法加以改进,由原来的切片复水后染色改为直接染色。
花芽分化进程观察:于2017年3月份,随机剪取对照组和“抑花一号”处理的雌株树冠中部东西南北四个方向12根50~100 cm长度的2年生健壮枝条,观察并记录其上柔荑花序及叶芽的数量。两者之和为枝条上总芽数。计算雌花芽数量所占百分比。
雌花芽百分比=(柔荑花序数量/总芽数(叶芽数量+柔荑花序数量))×100%
物候期观察:自2016年1月1日至7月25日,每3 d进行一次物候期观测。
数据分析方法:所得试验数据用Excel 2010软件进行整理、绘图,再利用SPSS 22.0软件做单因子差异性分析。
2 结果与分析
2.1 花芽外观形态特征
观测期间,绦柳雌花芽整体呈绿色,无明显颜色变化;外被绒毛,但明显毛量变化。由表1数据可见,观测期间,绦柳花芽的长度变化于1.88±0.24至2.67±0.40,P值为0.11,无显著差异,无法作为判定花芽分化进程的依据;宽度变化于1.15±0.14至1.62±0.31,P值为0.10,无显著差异,无法作为判定花芽分化的依据;长宽比变化于1.59至1.86,但呈现一定趋势变化规律,图1显示了从花芽分化开始到雌蕊分化结束时期,绦柳雌株花芽的外观大小变化,芽形状没有明显的变化,但在6月5日前芽的长宽比呈明显的下降趋势,6月5日长宽比降至最低值,随后呈升高趋势。这一变化规律与花芽分化进程相一致,但无法构成判定各分化过程的依据。
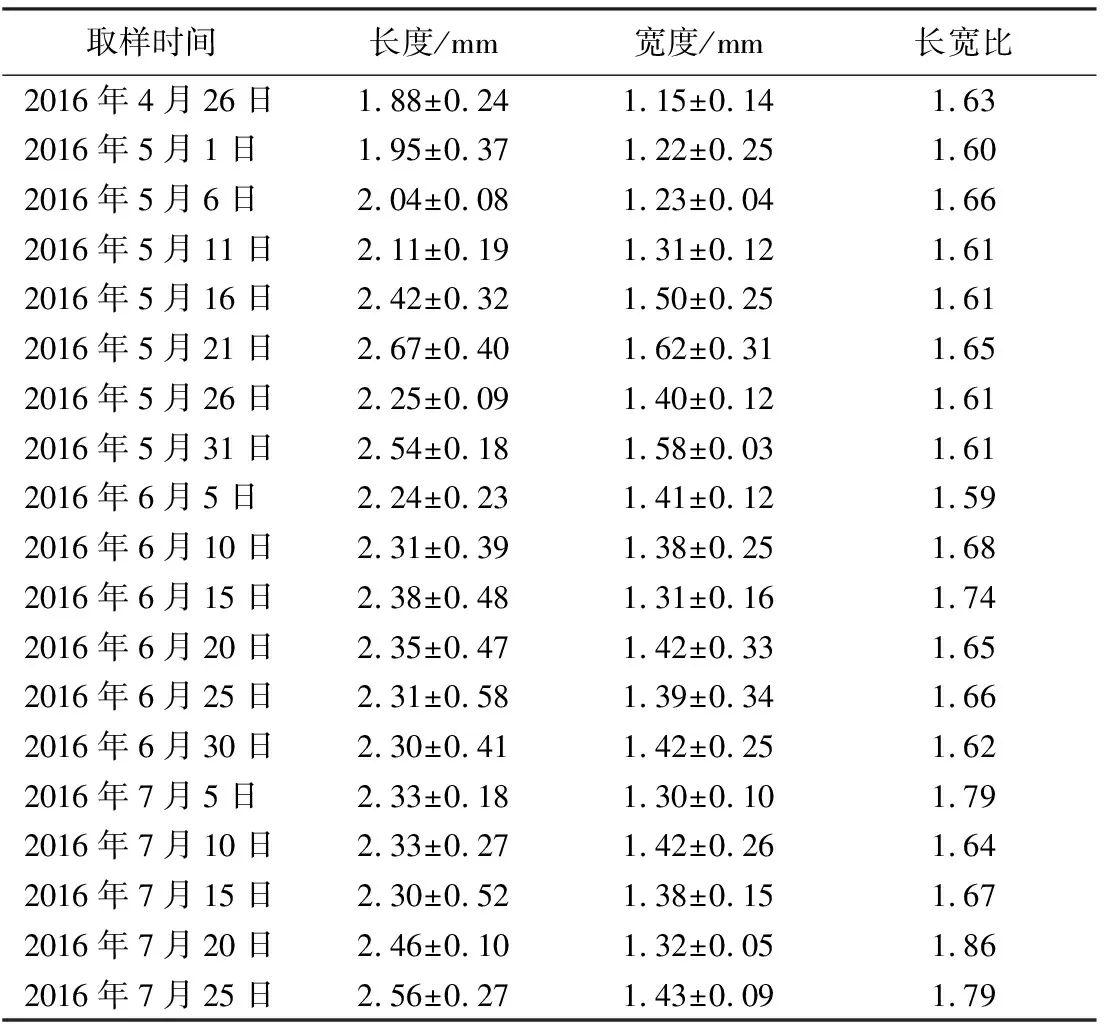
表1 绦柳雌花芽不同发育阶段的形态
2.2 花芽分化过程
据观察,绦柳雌株的芽为混合芽,雌花芽分化始于5月21日,观察期间经历了花序原基分化期、小花原基分化期、雌蕊分化期3个时期。
花序原基分化期:5月21日至5月31日,花芽外观较为干瘪瘦小,剥开芽鳞后绒毛尚未形成或较少。内部生长锥由尖变圆(图2A),形成花序轴(图2B),并在花序轴上开始形成小花原基突起(图2C)。
小花原基分化期:6月5日至7月10日,花芽基部渐宽、顶部明显伸长,整体圆润饱满,剥开芽鳞绒毛日趋增多。内部花序轴两侧开始分化出小苞片(图3A)。小苞片与花序轴之间的苞腋处生长点细胞分裂加剧,进而形成半球形隆起即小花原基(图3B、3C)。小花原基不断变大(图3D、3E、3F),与花序组织的细胞界线变明显(图3G、3H、3I)。雌花芽上所有小花原基在这一时期内基本全部形成(图3J)。
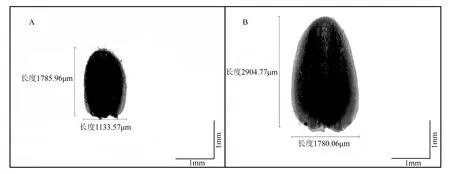
A.5月26日绦柳雌花芽,标尺=1 mm;B.7月20日绦柳雌花芽,标尺=1 mm。
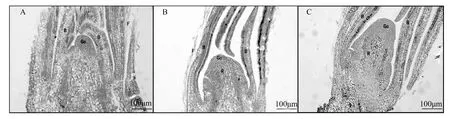
A.5月21日雌花芽,标尺=100 μm;B.5月26日雌花芽,标尺=100 μm;C.5月31日雌花芽,标尺=100 μm;Gc.生长锥;R.花序轴;B.苞片;F.绒毛。
雌蕊分化期:7月15日后,花芽外观形态较前期差异不明显。但内部小花原基突起不断增大,小苞片上下表面出现绒毛,之后形成雌蕊原基(图3K)。
此外,7月15日后注射“抑花一号”的雌株花芽的切片中,花序轴中下部的小花原基出现部分败育现象(图3L)。
2.3 花芽分化进程
根据图4数据,未处理的绦柳雌株花芽百分率为65.77%~65.79%。2016年6月5日以后“抑花一号”处理的雌株,雌花芽百分率开始升高;到7月15日以后,已有59.01%的侧芽分化为雌花芽。根据前文花芽分化阶段观察结果可知,6月5日至7月15日侧芽已分化出小花原基,并于7月15日前后进入雌蕊分化期。可见,6月5日后注射“抑花一号”已不能逆转侧芽向花芽分化的进程。而2016年6月5日前(即2016年4月26日—5月31日)注射“抑花一号”的绦柳雌株,2017年3月观察的雌花芽百分率为0~3.51%。由此推断,在临沂地区绦柳雌株的成花决定时间为6月5日至7月10日。
此外,在2017年3月份观察期间发现6月5日后注射“抑花一号”的雌株2年生枝条上部分柔荑花序有正常的花序轴和小苞片,但小花全部败育,无果实,不飞毛;随着时间推移,这样的花序从上到下越来越多(图5)。由此推断,绦柳雌株在6月5日,即小花原基分化期开始时进入花序决定态,花序轴及小苞片分化完成且无法逆转,但花序轴上的小花尚未进入小花决定态;在小花原基分化期应用“抑花一号”可使绦柳雌株花序上小花的分化发生逆转,无法完成全部分化,特别是雌蕊的分化,导致不开花或者小花败育,但花序的分化已完成,花序轴和其上的小苞片已形成无法逆转[21-22]。这与Ferguson et al.[23]豌豆分生组织或芽的测定结果一致。小花原基分化期是控制绦柳雌株成花决定的关键时期,抑制花序上小花的分化是防治次年飞毛的关键。
2.4 花芽分化时间与物候期
根据表2数据,绦柳雌花一般在果实开始脱落期(即飘絮开始期4月13日)后的38 d左右陆续进入花芽分化;52 d后完成花序决定态,进入小花原基分化期,开始花的决定;93 d后进入雌蕊原基分化期,从开始花芽分化到雌蕊分化完毕持续约103 d。采用化学药剂防治飞絮药物阻止绦柳雌株花芽分化的处理时间从花芽分化开始之前的25 d到小花原基分化前都有效果,即开始飞絮后的13~52 d。

A.6月5日雌花芽,标尺=100 μm;B.6月10日雌花芽,标尺=200 μm;C.6月15日雌花芽,标尺=100 μm;D.6月20日雌花芽,标尺=100 μm;E.6月25日雌花芽,标尺=100 μm;F.6月30日雌花芽,标尺=200 μm;G.7月1日雌花芽,标尺=200 μm;H.7月5日雌花芽,标尺=100 μm;I.7月10日雌花芽,标尺=100 μm;J.7月15日雌花芽,标尺=100 μm;K.雌蕊分化期,标尺=50 μm;L.小花原基败育,标尺=100 μm;R.花序轴;Fp.小花原基;b.小苞片;Pp.雌蕊原基;F.绒毛;Fa.小花败育。
图3绦柳花芽分化过程
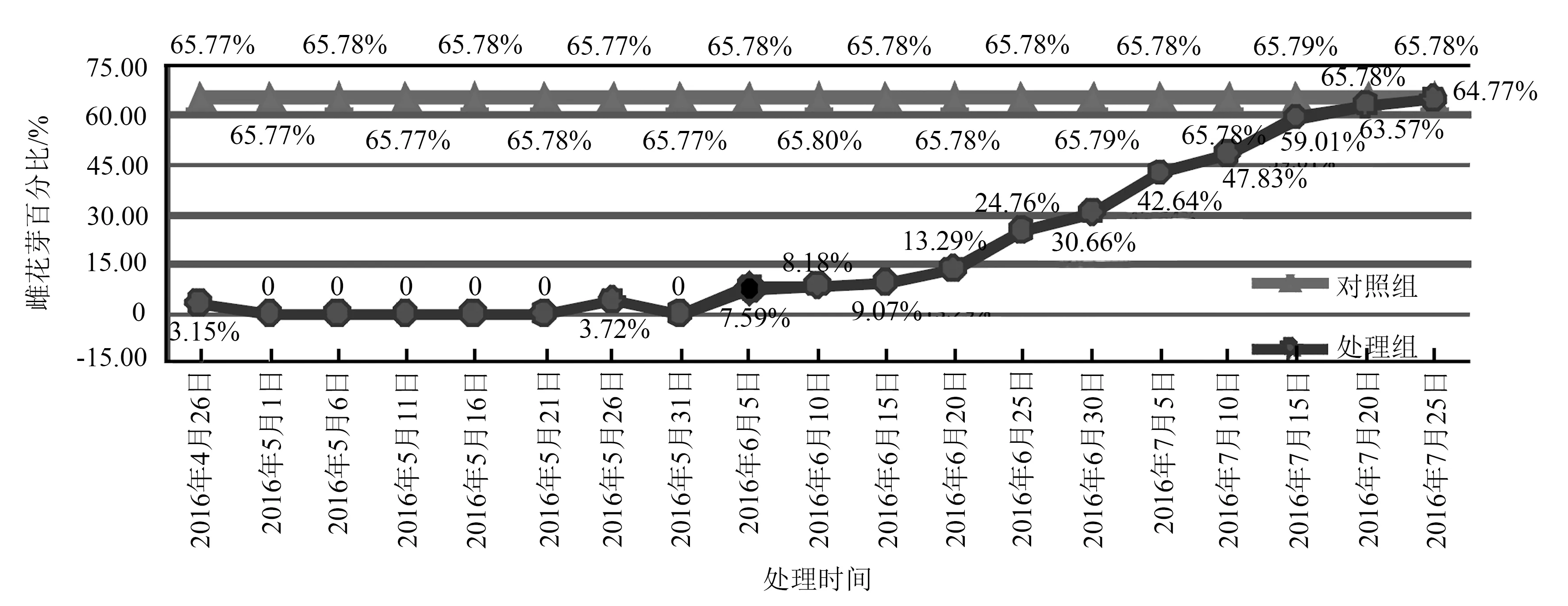
图4 2016年绦柳雌株雌花芽百分比(统计时间2017年3月份)

A.小花败育的雌花序,可见正常的花序轴和逆转成叶片功能的小苞片;B.雌花序,可见宿存的柱头;C.叶。

表2 2016年绦柳雌株物候观测记录
3 结论与讨论
临沂地区绦柳雌株成花决定时间为6月5日至7月10日。这与王建红等[14]在北京地区对垂柳的花芽分化发端时间(6月24日至7月14日)的试验结果差距较大。植物的发育是由遗传因素和环境条件共同决定的,其中光周期和温度是影响开花诱导的主要环境因子[24]。这种差异可能是由物种及气候的差异所致。
临沂地区5月份至8月份绦柳雌株花芽分化大致经历了花序原基分化期、小花原基分化期、雌蕊分化期3个时期。这一结果与张明照[16]的研究结果基本一致,但各时期开始时间略有提前,可能是由于临沂地区的环境气候较北京更为温暖、湿润,物候期整体提前有关。
试验期间绦柳物候期的观测记录表明:雌株一般在开始飘絮后的38 d左右进入花芽分化,52 d进入成花决定,整个过程持续约103 d。注射化学药剂物阻止绦柳雌花花芽分化的作用时间应在开始飞絮后的13~52 d。结合与郑景云等[26]做出的关于春季物侯期随地理位置的推移幅度规律,可以大致推算出北方其他地区的花芽分化发端及成花决定日期,建立了绦柳花芽分化进程与宏观物候变化的联系,打破飞絮控制技术应用的地域限制。
目前,利用石蜡切片法观察花芽分化的相关研究大多旨在建立花芽分化进程与花芽外观形态变化特征(如长度、宽度、长宽比、颜色等)的对应关系上,达到减少重复试验,指导生产应用的目的[15-16,25]。但是文中发现绦柳雌花芽外观形态变化并未呈现出如颜色变化、长宽比固定等显著特征。当雌株完成花序决定进入小花的决定(即小花原基分化期)时,芽外观一直为绿色并无明显颜色变化;芽体整体圆润饱满,长宽比无显著变化;此时剥开芽鳞可见被毛的幼叶,内部花序轴两侧小苞片及小花原基突起基本全部形成,但尚未分化出雌蕊原基。当雌蕊原基分化开始后,可见花序轴两侧小苞片上绒毛开始形成。因此,花序轴两侧小苞片上绒毛的有无可作为绦柳雌株成花决定期的判断依据之一,此时花序的小花原基进入雌蕊原基分化期。可以判断,此时小花原基已分化完毕。“抑花一号”对绦柳植株花芽分化的抑制,表明绦柳雌株花芽分化过程中,花序决定与小花决定是可以分开的。绦柳雌花芽可以形成花序,分化出花序轴且形成小苞片(当剥去雌株混合芽的芽鳞和幼叶,看见大量的绒毛包被着花序时),此时注射“抑花一号”仍可以有效抑制次年飞絮,但小花分化完成后(此时剥除芽鳞和幼叶,小苞片上形成绒毛,包被着小花原基时),注射“抑花一号”已无效果。这一结果与前任对金鱼草突变体、豌豆花序和花的分化观察结果基本一致[23,27]。
绦柳作为城市园林绿化常用树种,其飞絮治理问题一直影响着市民的生活。通过对绦柳花芽分化规律的研究,为飞絮控制技术的推广应用提供理论依据,在生态损益最小、社会影响最低的前提下,科学有效地解决飞絮问题。
参 考 文 献
[1] 王玉涛,李吉跃,胡东燕,等.常见绿化树种绦柳(Salixmatsudana‘Pendula’)耗水特性[J].生态学杂志,2008,27(12):2087-2093.
[2] 胡有宁.延安城市森林生态系统服务功能价值评估[D].杨凌:西北农林科技大学,2015.
[3] 陈崇.北京几个城市绿化树种生理生态特性的研究[D].北京:北京林业大学,2008.
[4] 李想,李海梅,马颖,等.居住区绿化树种固碳释氧和降温增湿效应研究[J].北方园艺,2008(8):99-102.
[5] 秦仲.北京常见园林植物群落的人体舒适度研究[C]//张启翔.中国观赏园艺研究进展.北京:中国林业出版社,2012:650-655.
[6] 赵松婷,李新宇,李延明.北京市常用园林植物滞留PM2.5能力的研究[J].西北林学院学报,2016,31(2):280-287.
[7] 周娟.浅谈杨柳飞絮抑制方法[J].西藏科技,2013(6):70-71.
[8] 乔建国,王晓媛.杨柳树飞絮的治理措施[J].河北林业科技,2008(5):46-47.
[9] 王世芬,张德银,郭建和,等.石硫合剂抑制杨柳飞絮实验研究[J].林业科技通讯,2015(3):28-29.
[10] 张志梅.杨柳飞絮控制新技术研究[J].中国农业信息,2014(11):41-42.
[11] 阚志霄,马俊荣.新法控制毛白杨飞絮[J].国土绿化,1999(6):43.
[12] 孙佳,刘占国.注干法抑制杨树飞絮试验[J].农村实用科技信息,2010(5):50.
[13] 王建红,车少臣,邵金丽,等.毛白杨飞絮控制技术的研究[C]//传统的继承与可持续发展:中国风景园林学会2010年会论文集(上册).北京:中国建筑工业出版社,2010:497-500.
[14] 王建红,车少臣,邵金丽,等.垂柳花芽分化发端时间的研究[J].中国农学通报,2008,24(12):173-178.
[15] 吴海.中国樱桃花芽分化规律的研究[J].林业科技开发,2007,21(5):27-29.
[16] 张明照.旱柳生殖生物学特性研究[D].北京:中国林业科学研究院,2014.
[17] 肖安琪.三角梅花芽分化的内源激素变化与花期调控研究[D].广州:华南农业大学,2016.
[18] SRIKANTH A, SCHMID M. Regulation of flowering time: all roads lead to rome[J].Cellular and Molecular Life Sciences,2011,68(12):2013-2037.
[20] 贺冰,李志岗,郝晓娟,等.植物材料快速石蜡制片方法[J].植物学报,2014,49(2):203-208.
[21] 傅永福,孟繁静.植物的成花决定[J].植物生理学通讯,1997,33(2):81-87,100.
[22] Battery N H, Lyndon R F. Changes in apical growth and phyllotaxis on flowering and reversion inImpatiensbalsaminaL.[J]. Ann Bot,1984,54(4):553-567.
[23] FERGUSON C J, HUBER S C, HONG P H, et al. Determination for inflorescence development is a stable state, separable from determination for flower development inPisumsativumL. buds[J]. Planta,1991,185(4):518-522.
[24] 岳彩鹏,韩锦峰,陈卫华.烟草开花研究进展[J].烟草科技,2001(9):36-40.
[25] 张美娇.蒙古柳(Salixlinearistipularis)雌雄株差异性分析[D].哈尔滨:东北林业大学,2016.
[26] 郑景云,葛全胜,赵会霞.近40年中国植物物候对气候变化的响应研究[J].中国农业气象,2003,24(1):28-32.
[27] COEN E S, ROMERO J M, DOYLE S, et al. Floricaula: a homeotic gene required for flower development inAntirrhinummajus[J]. Cell,1990,63(6):1311-1322.
