应用二类调查数据对塔林林场森林碳储量的估算
2018-06-15黄韵宁
黄韵宁
(北京林业大学,北京,100083)
森林生态系统以全球27%的陆地面积储存着80%以上的全球植被碳储量和40%以上的全球土壤碳储量[1],其不仅是陆地生态系统重要的碳库,更是全球碳循环与碳平衡研究的重要基础,因此现阶段森林生态系统碳计量研究正受到越来越多的重视。在全球、区域尺度上,Pan et al.[2]、方精云等[3]、王效科等[4]分别依据材积源生物量模型和森林资源连续清查统计数据估计全球和全国尺度森林碳储量,并发现全国尺度森林碳密度与地形、气候及人口密度等存在显著关系;此后,焦燕等[5]、李士美等[6]、甄伟等[7]在该方法基础上继续研究了我国不同省份碳储量及其动态变化特征,这些研究虽然为区域尺度森林碳储量估计提供了有效方法,但其所用材积源生物量模型存在建模样本数量、模型数量不足以及精度不高等问题。在此基础上,李海奎等[8]采用全国范围内主要树种单木相容性生物量模型和第七次(2004—2008年)森林资源连续清查样地数据,估算了全国范围内乔木林碳储量。在林分尺度上,我国学者开展了广泛研究,其优势与国外同行相比主要体现在相容性模型及思想的应用上,如王蒙等[9]采用单木相容性生物量模型和固定监测样地数据研究了黑龙江省落叶松(Larixgmelinii)人工碳储量及其动态变化特征;董利虎等[10]采用非线性似乎不相关模型构建了基于林分变量和生物量换算系数的相容性林分生物量模型,但这些研究多是对森林资源监测体系中固定样地数据的挖掘和应用,忽略了对森林资源监测体系中面妆数据的挖掘。
森林资源规划设计调查(简称二类调查)是我国森林资源监测体系的重要组成部分,其是以国有林业局(场)、自然保护区、森林公园等森林经营单位或县级行政区域为调查单位,其主要任务是查清森林、林地和林木资源的种类、数量、质量与分布,进而为森林资源经营管理决策提供数据支持[11]。在二类调查中,小班是最基本的调查单元(即面妆数据),调查过程中需要测定和测算的指标包括平均胸径、平均树高、林分密度、单位蓄积等,因此如何充分有效地借助该类数据实现林场尺度森林碳储量的估计是当前林业研究的重点。目前,周伟等[12]、崔传洋等[13]、唐才富等[14]分别用材积源生物量换算模型研究了不同地区典型林场的碳储量及其分布特征,但其所用模型和含碳率均为全国通用水平,未考虑不同气候、林型、起源及经营水平等因素对估算结果的影响。同时,他们的研究也未分别乔木林不同器官进行估算,使估算结果的实用性存在较大缺陷。为此,本研究以大兴安岭地区塔林林场为研究对象,基于2008年森林资源二类调查数据,运用邻近区域实测数据和所建材积源生物量转换模型研究该地区森林碳储量和碳密度及其分布特征,摸清该地区森林碳储量和碳汇能力,为构建区域尺度碳汇交易市场和建立碳汇效益补偿标准提供理论依据。
1 研究区概况
研究区域位于大兴安岭地区塔河林业局塔林林场(52°19′9″N,124°41′43″E),总经营面积1.39×105hm2,森林覆盖率达82.76%。全区属典型低山丘陵地貌,平均海拔535 m,平均坡度9°,约80%的小班处于斜坡及以下等级;气候属寒温带大陆性季风气候,年平均降水量463 mm,年平均气温-2.4 ℃,极端高温37.2 ℃(1992年),极端低温-45.8 ℃(1980年),年有效积温(≥10 ℃)1 276~1 969 ℃;区内土壤主要以棕色针叶林土为主,此外还伴有少量的草甸土、沼泽土等;区域内植被以兴安落叶松(Larixgmelini)、樟子松(Pinussylvestrisvar.mongolica)、白桦(Betulaplatyphylla)、山杨(Populusdavidiana)等树种为主,此外还有少量云杉(Piceakoraiensis)、蒙古栎(Quercusmongolica)、柳(Salixbabylonica)等。根据2008年森林资源二类调查数据,区域内林型主要以天然落叶松林、天然白桦林、阔叶混交林和针阔混交林为主,占区域总面积和总蓄积的76.69%和89.45%(表1)。

表1 塔林林场主要林分类型基本特征
注:*包括落叶松和樟子松人工林。表中除样本、总面积、总蓄积数据外均为平均值±标准差。
2 研究方法
2.1 碳储量估算
采用连续生物量扩展因子法(CBEF)研究林场尺度森林碳储量估计问题[14],其基本思路为:以森林资源二类调查数据中各小班林分蓄积为计算基础,通过各林型不同器官(包括树干、树枝、树叶和树根)CBEF函数计算其不同成分生物量,然后再乘以该林型各器官碳含率计算其累计碳储量。显然,基于该方法计算出来的是以立木为主体的森林生物量碳储量和碳密度,其计算公式为:
式中:C为林分单位面积总碳储量,即总碳密度,单位为t·hm-2;Ci为林分第i个器官碳储量,包括树干、树枝、树叶和树根,单位均为t·hm-2;ci为林分第i个器官碳含率,因林分类型和器官而异;V为林分蓄积,单位为m3·hm-2;ai和bi分别为待估参数,同样因林型和器官而异;BEF,i为第i个器官生物量扩展因子,计算公式为BEF,i=生物量(Bi)/蓄积量(Vi))[15]。研究区域各林型生物量换算系数采用董利虎提供的大兴安岭主要森林类型CBEF模型[16],各林型不同器官碳含率采用贾炜玮提供的东北地区主要树种实测数据[17]。
2.2 影响因素分级
林分碳储量和碳密度是林地内生物、非生物以及人为干扰等多种因素长期综合作用的结果,因此根据国内外相关研究成果[12-14]以及森林资源二类调查数据的可用性,定量分析不同因素对林分碳密度的影响。生物因素主要包括林分类型、龄组等,鉴于天然林是区域内森林资源的主体(表1),因此本研究不探讨起源的影响。非生物因素则涉及海拔、坡度、坡向、坡位等,这些因素可通过调节植被生境所需的水、热、光照等微气候影响森林碳储量和碳密度。由于研究区域森林已经经历了长达百年的无序采伐且经营档案严重缺失,因此人为干扰(主要指采伐)可通过上述地形因子间接来反映。实践中,海拔越高、坡度越陡、坡位越高则受人为影响的概率越低。林分类型根据区域内各树种分布面积以及CBEF模型的完整性划分为天然落叶松林、天然白桦林、天然樟子松、天然杨树林、针叶混交林、针阔混交林、阔叶混交林、人工林以及其他9种类型,将区域内水域、道路、居民点、沼泽地等归为其他。将龄组划分为幼龄林、中龄林、近熟林、成熟林和过熟林5类,将坡度分为平坡、缓坡、斜坡、陡坡、急坡和险坡6类,将坡位分为山脊、上坡、中坡、下坡、谷地和平地6类。根据区域内地形特征和经营实践,将海拔划分为<400,[400,500),[500,600),[600,700),[700,800),≥800 m共6个等级,将坡向划分为无坡向、阴坡、半阴坡、半阳坡和阳坡5级。
3 结果与分析
研究结果表明,区域内总平均碳密度和各器官碳密度均呈现出显著的破碎化趋势(图1),说明区域内森林资源遭到严重人为干扰。截至到2008年塔林林场全境森林碳储量为4.29×106t,有林地平均碳密度为37.06 t·hm-2,其中人工林总碳储量仅为0.24×103t,平均碳密度也仅为1.43 t·hm-2,远低于同地区的天然林,这可能是由于区域内人工林现阶段多为造林未成林地所致。因此,后续仅分析相关生物和非生物因素对天然林碳储量和碳密度的影响。
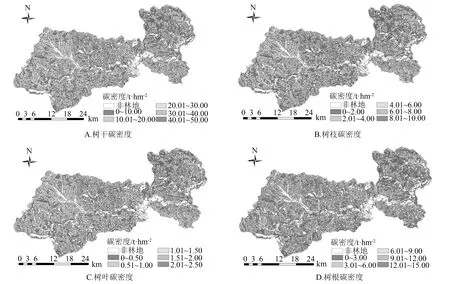
图1 塔林林场森林各器官碳密度空间分布特征
3.1 生物因素作用
区域内各林型碳储量和碳密度如表2所示。在天然林中,阔叶混交林的碳储量最高,达1.07×106t,其次分别为针阔混交林(0.97×106t)、天然白桦林(0.96×106t)和天然落叶松林(0.90×106t),这4种林型共占区域内总碳储量的90.92%。从各林型碳密度整体看,天然樟子松林的碳密度最高,达48.62 t·hm-2,其次为阔叶混交林44.80 t·hm-2,而天然白桦林碳密度最低,仅为27.81 t·hm-2。各林型不同小班碳密度平均变异系数为32.81%,其中天然白桦林最高(60.80%),针阔混交林最低(25.43%)。各林型不同器官碳密度由高到低依次为树干(24.02 t·hm-2)、树根(8.23 t·hm-2)、树枝(3.44 t·hm-2)、树叶(1.58 t·hm-2)。各林型树干碳密度所占比例平均达到64.95%,其中天然樟子松林树干碳密度所占比例最高(67.09%),其次为天然杨树林(66.99%),而阔叶混交林树干碳密度所占比例最低(61.82%)。
对区域内有林地小班分龄组统计结果表明(表3),各龄组树干和树根平均碳密度整体均随林分年龄的增加而增加,而树枝和树叶碳密度则无明显变化趋势。此外,由于树干和树根碳密度在全树整体碳密度中占绝对优势(87.09%),因此各龄组平均碳密度仍表现出随林分年龄的增加而增加的趋势,其中幼龄林、中龄林、近熟林、成熟林和过熟林分别为27.36、39.54、43.68、47.23、53.50 t·hm-2。各龄组不同小班碳密度平均变异系数为38.23%,其中幼龄林最高(61.00%),近熟林最低(26.44%)。在树木各器官内,树叶碳密度变异系数最大(62.74%),而树干变异系数最小(38.04%)。结合各龄组面积发现,中龄林碳储量最高,达2.78×106t,其次为幼龄林(1.16×106t),两者占区域总体的91.87%。
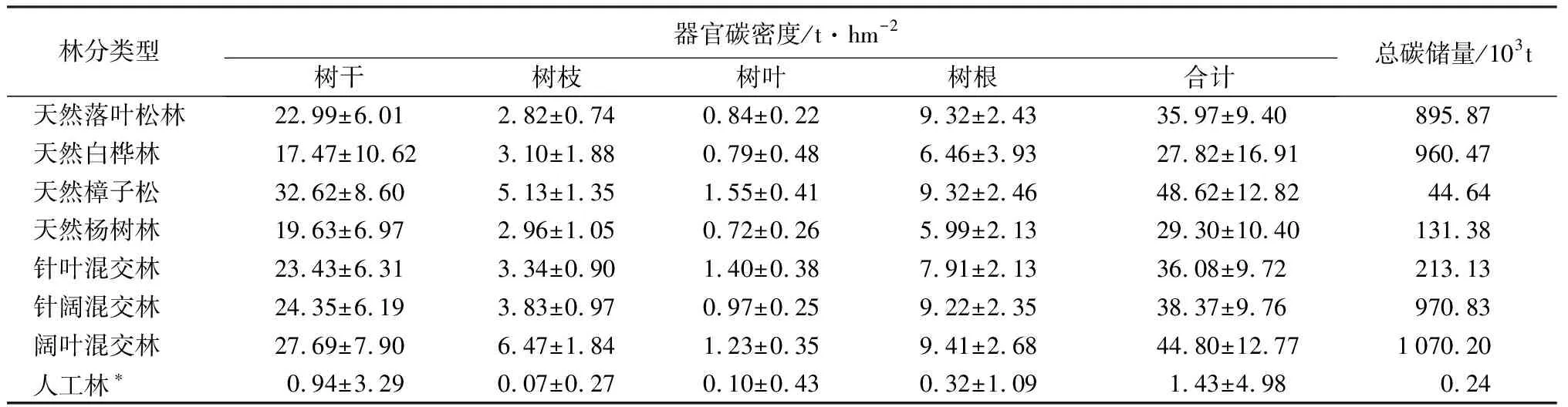
表2 塔林林场主要林分类型碳储量和碳密度特征
注:*包括落叶松和樟子松人工林。表中除总碳储量数据外均为平均值±标准差。
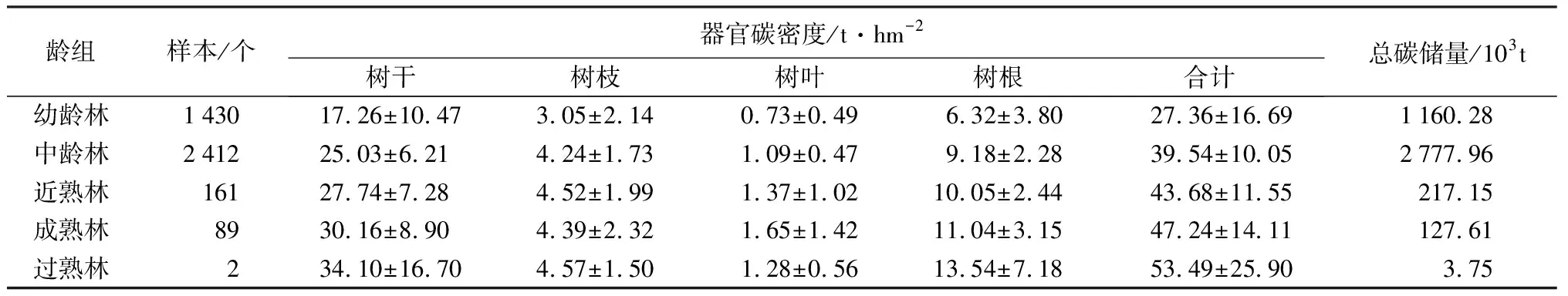
表3 塔林林场各龄组碳储量和碳密度特征
注:表中数据除样本和总碳储量数据外均为平均值±标准差。
3.2 非生物因素作用
地形因素对区域内森林碳密度的影响如表4所示。海拔500 m以下的平均碳密度明显较低,平均仅为35.05 t·hm-2,而高海拔(>500 m)地区碳密度则明显偏高(40.03 t·hm-2);坡度同样显著影响碳密度,其中平坡处碳密度最低(33.06 t·hm-2),而陡坡处碳密度最高(46.41 t·hm-2),但急坡处碳密度则略有下降(42.34 t·hm-2),这可能是随着坡度的不断增加,土壤厚度、水分、肥力等生境逐渐变差所致;从坡位来看,平地处碳密度整体偏高(42.19 t·hm-2),之后碳密度则随着坡位的增加呈显著增加趋势;从坡向来看,除无坡向处碳密度明显偏低外(33.92 t·hm-2),其余坡向对碳密度的影响并不显著,这可能是由于早期采用的木材生产策略所致。
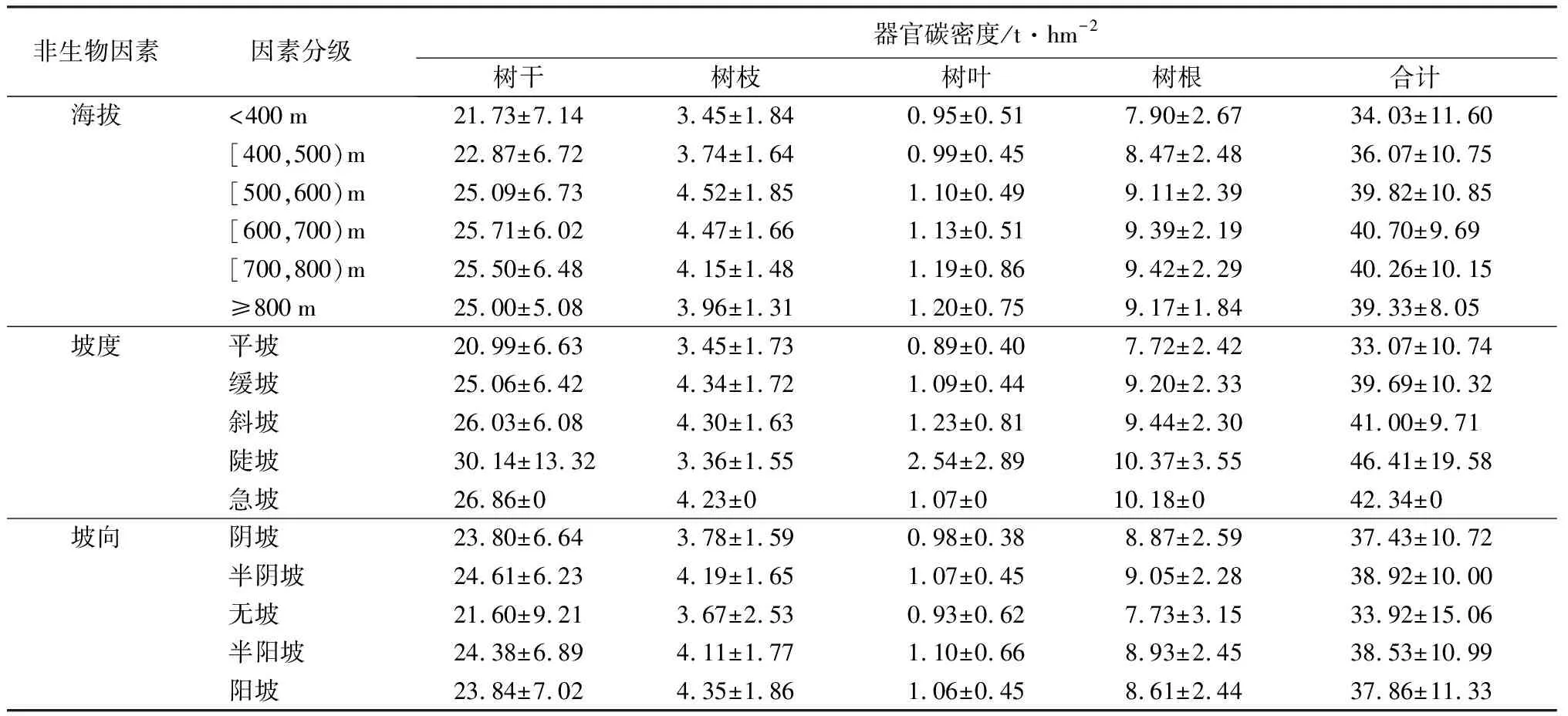
表4 塔林林场森林碳密度对地形因素的响应

续(表4)
注:表中数据为平均值±标准差。
4 结论与讨论
根据估算结果,大兴安岭塔林林场森林碳储量为4.29×106t,折合CO2约为15.89×106t,平均碳密度为37.06 t·hm-2,低于全国平均水平(42.82 t·hm-2),更远低于黑龙江省的平均水平(45.06 t·hm-2)[8],但接近于大兴安岭地区森林平均碳密度(36.46 t·hm-2)[18]。区域尺度内各器官碳密度呈现出明显的破碎化趋势,这一方面可能是由于当地复杂地形条件引起的微气候、微立地等生境差异所致,另一方面也可能由于当地长期无序的、高强度的森林采伐所致。在不同森林类型中,天然樟子松林的碳密度最高,达48.62 t·hm-2,其次为阔叶混交林44.80 t·hm-2,而天然白桦林碳密度最低,仅为27.81 t·hm-2,这与贾炜玮等[20]研究结果基本一致。由于区域内天然落叶松林、天然白桦林、针阔混交林和阔叶混交林的面积和蓄积处于明显优势地位,因此这4种林型总碳储量也最高,分别达到0.90×106t、0.96×106t、0.97×106t、1.070×106t,占区域总碳储量的90.92%。各林型不同器官碳密度按大小依次为树干(24.02 t·hm-2)、树根(8.23 t·hm-2)、树枝(3.44 t·hm-2)、树叶(1.58 t·hm-2),各器官所占比例平均依次为64.59%、22.23%、8.90%和4.29%,这与多数学者研究某种具体林型时碳的分配格局一致[9-10,19]。
乔木林平均碳密度与林分年龄呈显著正相关关系,这与贾炜玮等[17]、胡海清等[20]研究结果一致,但本研究进一步发现不同器官碳密度随林分年龄变化趋势有所差异,其中树干、树根碳密度整体随林分年龄增加,而树叶和树枝随林分年龄变化则无明显趋势。任继勤等[21]研究表明,黑龙江省18种主要森林类型碳密度和林分年龄均可用逻辑斯蒂模型拟合,且多数模型确定系数均在0.80以上。基于这些模型和《黑龙江省林地保护利用规划(2010—2020)》,进一步对黑龙江2014—2020年森林碳密度进行了预测,结果表明与2014年相比,全省范围内碳密度将增加4.90%,碳储量增加7.30%。因此,考虑到区域内森林目前主要以中幼龄林为主(93.84%),其正处于快速生长阶段,可认为该地区森林未来碳汇潜力巨大。但众所周知,该地区森林现阶段多为典型的过伐林,其结构、功能和质量均发生了显著退化,因此在后续管理中积极的人为干预措施(如森林抚育、林分结构优化)仍必不可少,以加速该地区森林资源的正向演替。
地形因素同样可显著作用于森林碳密度。研究结果表明,在一定范围内,乔木林平均碳密度整体随着海拔高度、坡度和坡位的增加而增加,这可能是随着地形特征的变化,人为干扰变得越来越困难,从而有利于林木生长和避免遭到采伐所致,同时地形因子也可间接影响植被生境所需的水热光照条件;但在极端条件下(如海拔≥800 m或急坡),乔木林碳密度也会略有降低,这可能是随着地形的变化,适于林木生长的土壤、水分等生境变差所致,这些结论与前人研究结论[22-23]基本一致。范叶青等[22]研究显示坡向、坡位对毛竹(Phyllostachysheterocycla)林生态系统碳储量及其空间分配均有一定影响,坡向对植被碳储量影响显著(P<0.05),对土壤和生态系统碳储量影响较显著(P<0.10),而坡位对植被碳储量影响极显著(P<0.01),对土壤和生态系统碳储量影响显著(P<0.05),但两者交互作用不显著;彭守璋等[23]研究表明,青海地区水热条件的差异使青海云杉(Piceacrassifolia)林生物量在空间上存在巨大差异,经度每增加1°,其生物量增加3.12 t·hm-2,纬度每增加1°,生物量减小3.80 t·hm-2,海拔每升高100 m,生物量减小0.05 t·hm-2。但因数据所限,本研究暂时还未能揭示多种因素对森林碳密度的交互影响。
与其他学者在研究区域尺度森林碳储量和碳密度估算时普遍采用国家尺度生物量扩展系数和碳含率(多为0.50或0.45)不同[2-4,12-14],本研究采用了基于实测数据的大兴安岭地区主要林型CBEF模型和碳含率实测数据,研究了区域范围内特定地块(即塔林林场)的森林碳储量和碳密度分布特征,研究结果能够较好地反映该地区森林的真实碳汇水平。但限于二类数据调查指标的限制,本研究暂未涉及土壤层、枯落物层、林下植被层的碳储量,因此可待数据进一步收集完备后,逐渐更深入细致地研究,以便更准确地把握林场尺度内森林碳储量及其动态变化。
参 考 文 献
[1] CAO M, WOODWARD F I. Net primary and ecosystem production and carbon stocks of terrestrial ecosystems and their responses to climate change[J]. Global Change Biology,1998,4(2):185-198.
[2] PAN Y D, BIRDESY R A, FANG J Y, et al. A large and persistent carbon sink in the world’s forests[J]. Science,2011,333:988-993.
[3] 方精云,郭兆迪,朴世龙,等.1981—2000年中国陆地植被碳汇的估算[J].中国科学D辑:地球科学,2007,37(6):804-812.
[4] 王效科,冯宗炜,欧阳志云.中国森林生态系统的植物碳储量和碳密度研究[J].应用生态学报,2001,12(1):13-16.
[5] 焦燕,胡海清.黑龙江省森林植被碳储量及其动态变化[J].应用生态学报,2005,16(12):2248-2252.
[6] 李士美,杨传强,王宏年,等.基于森林资源清查资料分析山东省森林立木碳储量[J].应用生态学报,2014,25(8):2215-2220.
[7] 甄伟,黄玫,翟印礼,等.辽宁省森林植被碳储量和固碳速率变化[J].应用生态学报,2014,25(5):1259-1265.
[8] 李海奎,雷渊才,曾伟生.基于森林清查资料的中国森林植被碳储量[J].林业科学,2011,47(7):7-12.
[9] 王蒙,李凤日,贾炜玮,等.黑龙江省落叶松人工林碳储量动态研究[J].植物研究,2013,33(5):623-628.
[10] 董利虎,李凤日.三种林分生物量估算方法的比较[J].应用生态学报,2016,27(12):3862-3870.
[11] 国家林业局.森林资源规划设计调查主要技术规定[M].北京:中国标准出版社,2011.
[12] 周伟,王晓洁,关庆伟,等.基于二类调查数据的森林植被碳储量和碳密度:以徐州市为例[J].东北林业大学学报,2012,40(10):71-74,88.
[13] 崔传洋,李建华,冯燕,等.基于二类调查数据的县级森林碳储量及碳密度测算:以山东省泗水县为例[J].山东农业大学学报(自然科学版),2017,48(2):279-283.
[14] 唐才富,张莉,罗艳,等.基于森林资源二类调查的青海乔木林碳储量分析[J].西部林业科学,2017,46(2):1-7.
[15] GUO Z D, FANG J Y, PAN Y D, et al. Inventory-based estimates of forest biomass carbon stocks in China: a comparison of three methods[J]. Forest Ecology and Management,2010,259(7):1225-1231.
[16] 董利虎.东北林区主要树种及林分类型生物量模型研究[D].哈尔滨:东北林业大学,2015.
[17] 贾炜玮,李凤日.东北林区各林分类型森林生物量和碳储量[M].哈尔滨:黑龙江科学技术出版社,2014.
[18] 韦昌雷,赵希宽,李慧仁.黑龙江大兴安岭森林碳储量与碳汇估算[J].防护林科技,2016(6):51-53.
[19] 任军辉,刘建军,刘斌,等.宁夏贺兰山天然油松林碳储量和碳密度[J].东北林业大学学报,2011,39(5):108-110.
[20] 胡海清,罗碧珍,魏书精,等.大兴安岭5种典型林型森林生物碳储量[J].生态学报,2015,35(17):5745-5760.
[21] 任继勤,夏景阳.基于碳密度-林龄关系的黑龙江省森林碳汇潜力预测[J].环境科学研究,2017,30(4):552-558.
[22] 范叶青,周国模,施拥军,等.坡向坡位对毛竹林生物量与碳储量的影响[J].浙江农林大学学报,2012,29(3):321-327.
[23] 彭守璋,赵传燕,郑祥霖,等.祁连山青海云杉林生物量和碳储量空间分布特征[J].应用生态学报,2011,22(7):1689-1694.
