2种生态环境下糯玉米单倍体加倍方法的比较
2018-06-08,,,,
, , , ,
(1.吉林农业科技学院农学院, 吉林 吉林 132101; 2.吉林省科学技术信息研究所, 长春 130033)
糯玉米(ZeamaysL.certainaKulesh)又称粘玉米,亦称蜡质玉米,起源于中国,是西南地区的硬质玉米发生突变,经人工选育而逐渐显现了糯质类型[1]。籽粒中有较粗的蜡质状胚乳,较像硬质玉米和马齿型玉米有光泽的玻璃质(透明)籽粒,其化学性状和物理性状受单个隐性基因(WX)控制,这个基因位于第9染色体上[1]。糯玉米富含蛋白质、维生素A、维生素B1等营养物质,口感香甜细腻、风味独特,深受人们青睐[2]。糯玉米已成为新形势下推动我国农业生产高效、农民增收的一项新兴产业。我国不同生态地区糯玉米种质资源极其丰富,但对于糯玉米育种的研究起步较晚,育种家对大量的种质资源缺乏有效的育种技术手段,从而造成了糯玉米育种田间工作量大、育种效率较低[3]。因此,迫切需要通过新型育种手段选育品质好、综合抗逆性优良的品种类型,以满足多元化市场需求。
目前,单倍体技术已经成为普通玉米的主流选系技术,但将该技术运用于糯玉米育种中的研究报道较为鲜见。玉米DH(Doubled Haploid)育种中的关键环节之一,即加倍问题,已成为各大科研院所的研究重点[4-5]。当前玉米单倍体的化学加倍法主要是指通过配置不同秋碱药剂浓度对单倍体进行人工加倍的一种加倍方法[6-7]。而玉米单倍体的自然加倍法则指未经任何人工处理时其染色体自发加倍的过程,是一种自然的、安全的加倍方式。在大量秋碱化学加倍普通玉米单倍体的试验中,采取滴心叶法进行单倍体的加倍时对植株伤害较小,且加倍率亦较高[8]。鉴于此,本试验采用高频孤雌生殖诱导系NY 10对东华北鲜食糯玉米区域试验对照品种京科糯569进行杂交诱导,以获得糯玉米单倍体,通过设计海南崖州、吉林公主岭2个加倍环境,采取秋碱滴心叶法及自然加倍法对单倍体进行加倍处理,进一步探讨糯玉米单倍体加倍的实用技术,以期为加快糯玉米DH育种进程提供参考。
1 材料与方法
1.1 试验地概况
吉林公主岭地理位置为北纬43°11′~44°09′,东经124°02′~125°18′,属中温带湿润地区大陆性季风气候,年平均气温5.6 ℃,年均降水594.8 mm。
海南崖州地理位置为北纬18°09′~18°37′,东经108°56′~109°48′,属热带海洋性季风气候,年均气温25.7 ℃,年均降水1 347.5 mm,有“天然温室”之称。
1.2 试验材料
糯玉米杂交种京科糯569(N 39/白糯6)、高频孤雌生殖诱导系NY 10均由吉林农业大学特用玉米育种团队提供。秋水仙素原药购自长春长生生物科技股份有限公司。
表2 不同环境下滴心叶法与自然加倍散粉率的方差分析
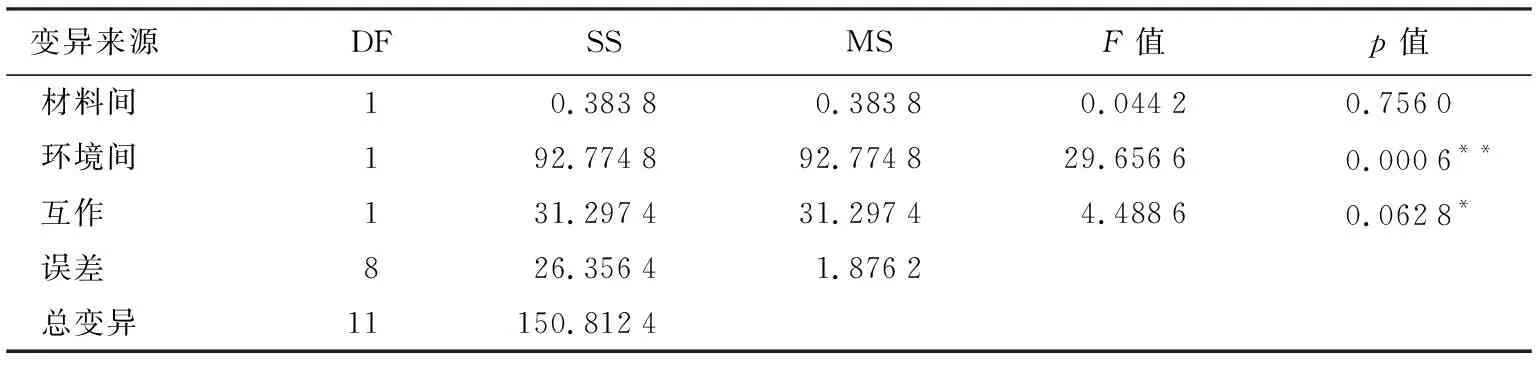
变异来源DFSSMSF值p值材料间10.38380.38380.04420.7560环境间192.774892.774829.65660.0006**互作131.297431.29744.48860.0628*误差826.35641.8762总变异11150.8124
注:“**”表示差异极显著(p<0.01),“*”表示差异显著(p<0.05)。下同。
表3 不同环境下滴心叶法与自然加倍法的结实率比较
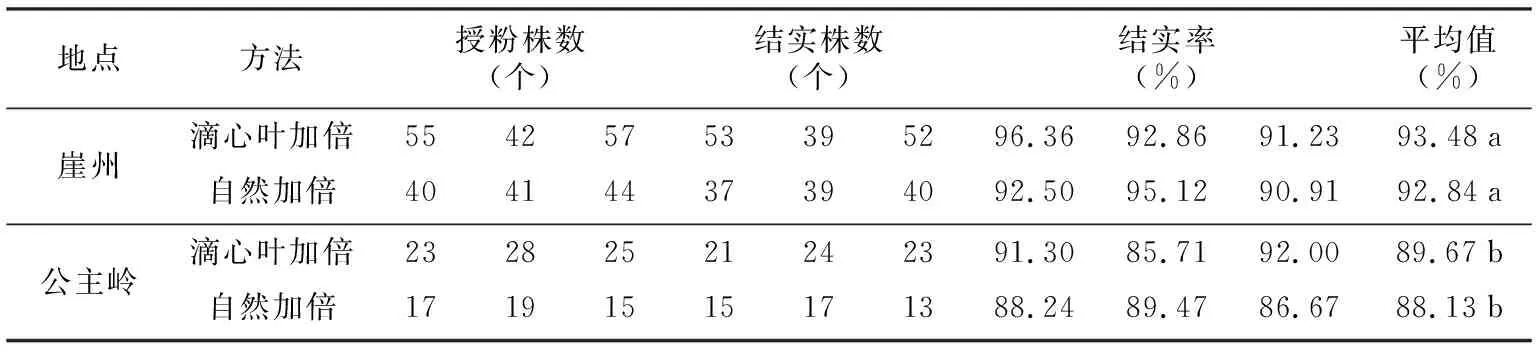
地点方法授粉株数(个)结实株数(个)结实率(%)平均值(%)崖州滴心叶加倍55425753395296.3692.8691.2393.48a自然加倍40414437394092.5095.1290.9192.84a公主岭滴心叶加倍23282521242391.3085.7192.0089.67b自然加倍17191515171388.2489.4786.6788.13b
表1 不同环境下滴心叶法与自然加倍法的散粉率比较

地点方法单倍体株数(个)散粉株数(个)散粉率(%)平均值(%)崖州滴心叶加倍49749648860476212.079.4812.7011.42a自然加倍49049448352534910.6110.7310.1410.50a公主岭滴心叶加倍4854734852729305.576.136.195.96b自然加倍4804814701720163.544.163.403.70c
注:小写字母表示差异显著(p<0.05)。下同。
1.3 试验方法
1.3.1 单倍体的诱导
2016年春在吉林长春种植母本糯玉米杂交种京科糯569共20行,行长5 m,每行20株,行距65 cm。父本诱导系NY 10进行错期播种,分别于母本播种的20 d、25 d后进行播种以保证父母本花期相遇,对母本去雄后进行杂交诱导。收获杂交诱导果穗,进行糯玉米单倍体的挑选。
1.3.2 单倍体籽粒鉴定
根据Chase[9]提出的单倍体籽粒Navajo遗传标记鉴定法进行单倍体籽粒的室内鉴定,以获得拟单倍体籽粒;根据R-nj植株标记鉴定法对拟单倍体籽粒进行田间种植鉴定[10],去杂去伪后以确定最终糯玉米单倍体株数。
1.3.3 单倍体加倍处理方法
2016年冬在海南崖州,将挑选的500粒拟单倍体种子播种于田间,进行糯玉米单倍体植株的田间鉴定以确定糯玉米单倍体株数。待糯玉米单倍体幼苗长到4~5片叶时,用微量移液器抽取3μL的0.5 mg/mL秋水仙素+2.5% DMSO+5%甘油溶液滴于幼苗心叶处。记录及统计糯玉米单倍体植株的散粉率及结实率;将挑选的另一份500粒拟单倍体种子种于同一地块,田间管理水平与上述滴心叶法加倍处理相同,进行糯玉米单倍体植株的田间鉴定以及观察单倍体植株的自然加倍情况,记录及统计糯玉米单倍体植株的散粉率及结实率。每个处理重复3次。2017年春在吉林公主岭进行与2016年冬试验过程同样的试验处理。
1.3.4 数据统计与分析
糯玉米单倍体的散粉率及结实率的计算方法如下[8]:
散粉率(%)=散粉株数/单倍体总株数×100%;
结实率(%)=单倍体自交结实株数/单倍体总株数×100;
综合利用Excel 2013软件和DPS 7.05软件进行数据统计分析[11]。
2 结果与分析
2.1 不同环境下滴心叶法与自然加倍法的散粉率比较
将不同环境下滴心叶法与自然加倍法散粉率以及方差分析结果列于表1和表2。由表1、表2可知,海南崖州糯玉米单倍体植株散粉率较吉林公主岭有较大提高且达极显著水平,其中,滴心叶法提高了5.46%,自然加倍法提高了6.80%。在海南崖州,滴心叶法与自然加倍法均超过10%,二者间未达到显著水平,但是环境和处理间的互作达到了显著水平,说明环境和加倍方式都能够对糯玉米单倍体的雄穗散粉产生影响。海南崖州地区的2种加倍处理方式均显著高于吉林公主岭地区2种加倍处理方式,表明海南崖州试验地种植的单倍体植株更容易加倍,是较吉林公主岭更为适宜的环境。
2.2 不同环境下滴心叶法与自然加倍法的结实率比较
本试验中统计单倍体结实株率时先统计实际雌雄协调株数,即花期相遇能够正常授粉的株数,将实际授粉株数作为统计结实率的依据。将不同环境下滴心叶法与自然加倍法结实率以及方差分析结果列于表3和表4。由表3、表4可知,在海南崖州,滴心叶法与自然加倍的结实率没有明显差异,均高于80%,且均高于吉林公主岭对应的处理方法。结实率方差分析表明,相对于处理方法,环境是影响糯玉米单倍体结实率更为重要的因素,适宜的田间环境能够有效提高结实率,而不同的加倍处理方法对糯玉米单倍体的结实率没有显著影响。
表4 不同环境下滴心叶法与自然加倍结实率的方差分析
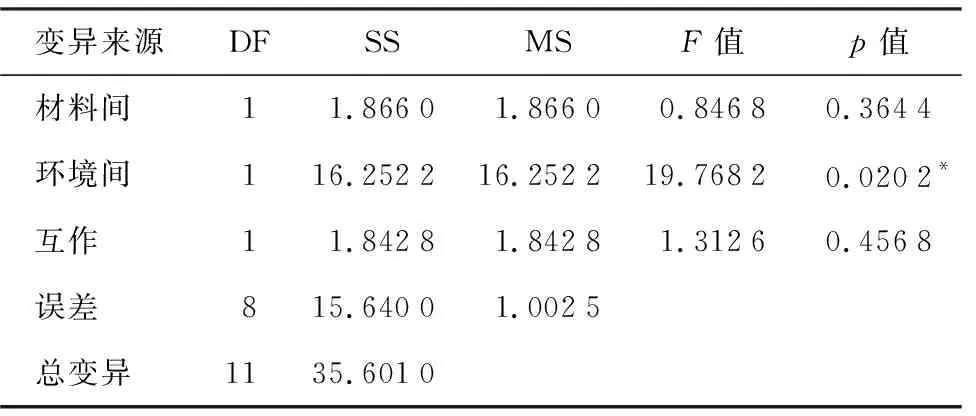
变异来源DFSSMSF值p值材料间11.86601.86600.84680.3644环境间116.252216.252219.76820.0202*互作11.84281.84281.31260.4568误差815.64001.0025总变异1135.6010
3 讨 论
通过高频孤雌生殖诱导系诱导目标母本基础材料可产生单倍体植株,经自然(化学)加倍成为纯合双单倍体-DH系,以此作为玉米育种的原始材料,加速了玉米自交系的纯合过程,大大缩短了育种进程。在玉米DH育种中,单倍体加倍方法是近年来育种家的研究重点,其中自然加倍法以具有节约育种成本、对人畜安全等突出的优点,正逐渐被广大玉米育种工作者所接受[12-13]。
才卓等研究表明,玉米自身遗传系统具有单倍体雄穗自然加倍遗传恢复(修复)能力,利用自然加倍法获得更多的纯系是可能的[14]。文科研究表明,普通玉米单倍体雄穗育性的恢复与环境温度有关,生长前期昼夜温差较小的自然环境对单倍体雄穗育性恢复不利,而昼夜温差较大的自然环境有利于雄穗育性的恢复[15];刘志增等研究认为,单倍体雄穗育性的恢复与其生长分化时期所处的田间环境温度具有负(或正)相关趋势[16];段民孝等研究表明,与其他地点相比,北京春季和海南冬季进行田间播种更有利于玉米单倍体的自然加倍[17]。本试验结果表明,不同环境与加倍处理方法间的互作有显著差异,在海南崖州环境下,自然加倍法对提高散粉率、结实率均优于吉林公主岭,与前人在普通玉米上的研究结果一致。
海南崖州冬季光、温、肥水充足,易于糯玉米单倍体植株生长;而吉林公主岭糯玉米单倍体生长季节多发生干旱、田间冷凉,使得单倍体植株生长受阻,相比之下海南更适合糯玉米单倍体发育,这可能是海南自然加倍率高的原因。综上,在进行糯玉米单倍体DH育种时,通过筛选适宜的生态环境并提高田间管理水平能够有效加快糯玉米DH育种进程。
参考文献:
[1]景桂昕,姜龙,王薪淇,等.东北地区几个糯玉米DH系单株产量的配合力分析[J].种子,2015,34(9):82-85.
[2]李艳茹,吉士东,大浩,等.糯玉米的营养价值和发展前景[J].延边大学农学学报,2003,25(2):146-148.
[3]李向永,姜龙,王薪淇,等.糯玉米单倍体诱导和加倍的研究[J].西北农林科技大学学报(自然科学版),2016,44(8):91-96.
[4]王文洁,周联东,孙佩,等.玉米单倍体加倍方法研究进展[J].安徽农业科学,2014,25:8 542-8 543.
[5]王江浩,尤帅,赵爱菊,等.玉米单倍体的诱导、加倍技术及其应用研究[J].河北农业科学,2016(1):70-75.
[6]邓昆鹏,李向永,邢政,等.高效糯玉米单倍体加倍方法的研究[J].种子,2017,36(1):82-85.
[7]陈绍江,黎亮,李浩川,等.玉米单倍体育种技术(第2版)[M].北京:中国农业大学出版社,2012,3.
[8]姜龙,王薪淇,李向永,等.爆裂玉米单倍体诱导和加倍方法的研究[J].作物杂志,2015(3):52-56.
[9]Chase S.S.Produciton of homozygous diploids of maize from monoploids[J].Agron J,1952(44):263-267.
[10]姜龙.玉米单倍体诱导和加倍技术的优化及DH系的遗传性研究[D].长春:吉林农业大学,2014.
[11]唐启义,冯明光.实用统计分析及其DPS数据处理系统[M].北京:科学技术出版社,2002:304-311.
[12]王志永,郑淑云,吴金鑫,等.玉米单倍体自然加倍与化学加倍研究[J].玉米科学,2015(6):18-20.
[13]吴鹏昊,任姣姣,田小龙,等.玉米单倍体自然加倍若干问题探讨[J].玉米科学,2016(4):7-11.
[14]才卓,徐国良,任军,等.玉米单倍体雄穗自然加倍性轮选遗传修复与高加倍率材料的创制[J].玉米科学,2016,24(4):1-6.
[15]文科.玉米单倍体诱导和加倍方法及其SSR和ISSR标记分离研究[D].北京:中国农业大学,2003.
[16]刘志增,宋同明.玉米单倍体雌雄育性的自然恢复以及染色体的化学加倍[J].作物学报,2000,26(6):947-952.
[17]段民孝,赵久然,刘新香,等.不同种植地点对玉米单倍体自然加倍率的影响[J].作物杂志,2012(2):68-70.
