薇甘菊替代控制技术研究
2018-06-05张知晓泽桑梓季梅刘凌户连荣
张知晓,泽桑梓,季梅,刘凌,户连荣
(1.云南省林业科学院,云南 昆明 650201;2.云南省林业有害生物防治检疫局,云南 昆明 650051)
薇甘菊(Mikaniamicrantha)是菊科(Asteraceae)假泽兰属(Mikania)的一种入侵杂草[1]。因其严重影响农林畜牧业的生产,被国家林业局列入21种林业检疫性有害生物之一[2]。近年来,人们对其物理、化学和生物防治技术进行大量研究[3]。但由于各项技术均有其弊端,在薇甘菊防治方面目前尚未看到较理想的防治方法[4-6]。
替代控制技术,是选择一种或多种适应性强、生长快、具经济价值、在短时间内可以达到较高郁闭度的植物,种植在外来入侵植物大面积危害的地方,以取代外来物种种群优势的方法[7]。选择乡土植物与杂草混种,对杂草的防控可起到长久持续的效果[8],且能创造较高的经济效益。目前,已有幌伞枫(Heteropanaxfragrans)[9]、血桐(Macarangatanarius)[10]、红薯(Lpomoeabatatas)[11]成功控制薇甘菊的案例,这为进一步开发薇甘菊的替代控制技术提供重要参考。中国滇西地区薇甘菊发生面积广,但至今还没有成套的替代控制技术规范[12]。为此,在当地筛选和研究可替代控制薇甘菊的乡土植物及替代模式,探究薇甘菊替代控制技术,对实现薇甘菊在当地的可持续控制具有极为重要的科学价值和现实意义。
1 材料与方法
1.1 供试材料
7种牧草分别为柱花草(Stylosantheshumilis)、东非狼尾草(Pennisetumclandestinum)、多年生黑麦草(Loliumperenne)、鸭茅(Dactylisglomerata)、杂三叶(Trifoliumhybridum)、紫花苜蓿(Madicagosativa)、苇状羊茅(Festucaarundinacea)。
5种林木分别为麻竹(Dendrocalamuslatiflorus)、勃氏甜龙竹(Dendrocalamusbrandisii)、杉木(Cunninghamialanceolata)、西南桦(Betulaalnoides)、旱冬瓜(Alnusnepalensis)。
1.2 草本植物替代控制薇甘菊试验
2011年6月-2014年10月,在德宏州瑞丽市姐勒水库(23°3′6.5″N,97°53′52.8″E,海拔800m),进行草本植物替代控制薇甘菊试验。试验前,采取人工清除薇甘菊的方法,将试验地块内的薇甘菊及其它杂草清除干净,翻地、整平。将试验地块分成1m×1m的若干小样方,样方间隔0.5m。为提高发芽率,采用变温浸种方法打破种子休眠。牧草种子播种密度梯度为3 000粒/m2、6 000粒/m2、9 000粒/m2,选择长度为0.15m、木质化程度基本一致的薇甘菊茎节,扦插密度为25枝/m2,牧草与薇甘菊茎节混播,每梯度设置3次重复,并设1组牧草空白对照及1组薇甘菊空白对照,自然条件下生长。60d后,测量薇甘菊的主茎长度、分枝数量、生物量以及牧草的株高,分别与对照组进行比较,筛选出对薇甘菊替代控制作用较强的优势植物种类及替代模式。
1.3 林木替代控制薇甘菊试验
2011年6月,在德宏州瑞丽市帕色林地(24°2′13.8″N,97°21′20.6″E,海拔502m),选取薇甘菊危害盖度平均达85%的废弃撂荒坡地5块,各0.67hm2,进行林木替代控制薇甘菊试验。种植的林木苗为1年生,初始栽植密度分别为麻竹4m×5m、勃氏甜龙竹5m×6m、杉木1m×1m、西南桦2m×3m、旱冬瓜1.5m×2m;种植当年施底肥,以后每年年底追肥1次;种植后3年内对幼苗进行人工抚育。2014年11月,调查麻竹、勃氏甜龙竹、杉木、西南桦和旱冬瓜5种薇甘菊-林木的替代控制试验基地,记录各种林木的郁闭度和林下的薇甘菊盖度。
2 结果与分析
2.1 草本植物替代控制薇甘菊的效果
2.1.1 牧草生长状况分析
7种牧草与薇甘菊混种时,各替代植物株高见图1。在3 000粒/m2、6 000粒/m2和9 000粒/m2种植密度下,柱花草与薇甘菊混种时株高分别为0.49m、0.48m和0.46m,均显著高于其它6种草本植物(P<0.05);其次为东非狼尾草,株高分别为0.31m、0.31m和0.28m,显著高于其它5种植物(P<0.05)。7种替代植物中,以杂三叶的株高最低,3种种植密度下,分别为0.07m、0.07m和0.05m(图1)。比较不同种植密度对同种植物株高影响发现,随替代植物种植密度增大,各植物株高降低,差异不显著(P>0.05)。
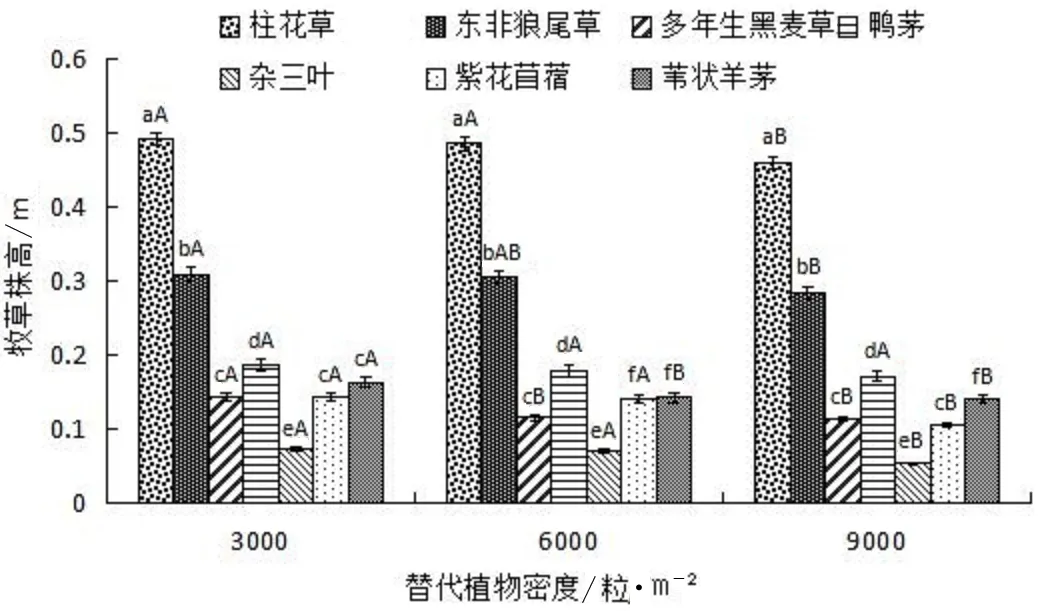
图1 不同密度7种替代植物株高分析
当替代植物与薇甘菊混种时,调查生境中其它植物种类的情况,结果见表1。柱花草和东非狼尾草样地中其它植物种类最少,其中柱花草生境中无其它植物,东非狼尾草生境中仅有阔叶丰花草(Borrerialatifolia)1种植物;在多年生黑麦草和鸭茅样地中,其它植物种类最多,多达8种。

表1 牧草中其它物种种类
注:柱花草(Stylosantheshumilis)、东非狼尾草(Pennisetumclandestinum)、多年生黑麦草(Loliumperenne)、鸭茅(Dactylisglomerata)、杂三叶(Trifoliumhybridum)、紫花苜蓿(Madicagosativa)、苇状羊茅(Festucaarundinacea)、阔叶丰花草(Borrerialatifolia)、含羞草(Mimosapudica)、野茼蒿(Crassocephalumcrepidioides)、胜红蓟(Ageratumconyzoides)、牛筋草(Eleusineindica)、毛豨莶(Siegesbeckiapubescens)、金腰箭(Synedrellanodiflora)、刺苋(Amaranthusspinosus)。
2.1.2 牧草在不同密度下对薇甘菊的影响
比较薇甘菊与几种不同种植密度牧草混种时的主茎长度,结果见图2。
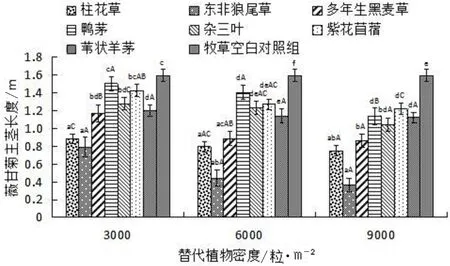
图2 不同密度7种替代植物对薇甘菊主茎长度的影响
由图2可知,牧草空白对照组的平均主茎长度为0.02m,薇甘菊与牧草混种时的主茎长度均小于薇甘菊单种时的主茎长度。在3 000粒/m2、6 000粒/m2、9 000粒/m2种植密度下,薇甘菊与东非狼尾草混种时主茎长度最低,分别为0.79m、0.44m和0.37m;其次是与柱花草混种,主茎长分别为0.88m、0.80m和0.75m。与两种植物混种时薇甘菊主茎长差异不显著(P>0.05),而均显著低于其它几种植物。在替代植物密度为3 000粒/m2和6 000粒/m2条件下,与鸭茅混种的薇甘菊主茎最长,分别为1.50m和1.41m。而在9 000粒/m2条件下,薇甘菊与紫花苜蓿混种时主茎最长(1.22m)。就同种替代植物不同种植密度而言,随着替代植物种植密度逐渐增大,薇甘菊主茎长度逐渐减小,但并无显著差异。
牧草种类对薇甘菊分枝数量影响见图3。
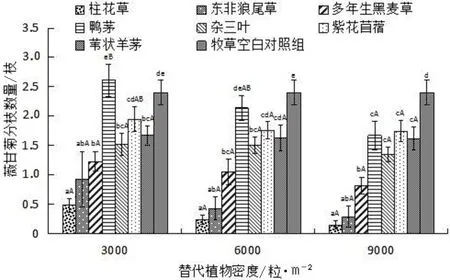
图3 不同密度7种替代植物对薇甘菊分枝数量的影响
由图3可知,薇甘菊对照组中平均分枝数量为2.40枝,在3种替代植物密度条件下,与柱花草混种的薇甘菊分枝数均最少,分别为0.48枝、0.24枝和0.15枝;其次为与东非狼尾草混种的薇甘菊分枝数,分别为0.93枝、0.43枝和0.29枝。二者对薇甘菊分枝的控制作用无显著差异。在替代植物密度为3 000粒/m2和6 000粒/m2条件下,与鸭茅混种的薇甘菊分枝最多,而在9 000粒/m2条件下,与紫花苜蓿混种的薇甘菊分枝数最多。就同种替代植物不同种植密度而言,随着替代植物种植密度逐渐增大,薇甘菊分枝数逐渐减小,但并无显著差异。
比较薇甘菊与7种牧草在不同牧草种植密度条件混种时的生物量,结果见图4。牧草空白对照组的生物量为0.53g/m2,薇甘菊与牧草混种时的生物量均小于薇甘菊单种时的生物量。在3 000粒/m2、6 000粒/m2和9 000粒/m2种植密度下,薇甘菊与柱花草混种时生物量分别为0.06g/m2、0.06g/m2和0.04g/m2;与东非狼尾草混种时生物量分别为0.07g/m2、0.03g/m2和0.01g/m2。与2种植物混种时薇甘菊生物量差异不显著(P>0.05),而均显著低于其它几种植物。在替代植物密度为3 000粒/m2和6 000粒/m2条件下,与鸭茅混种的薇甘菊生物量最大。而在9 000粒/m2条件下,薇甘菊与苇状羊茅混种时生物量最大。就同种替代植物不同种植密度而言,随着替代植物种植密度逐渐增大,薇甘菊生物量逐渐减小,但并无显著差异。
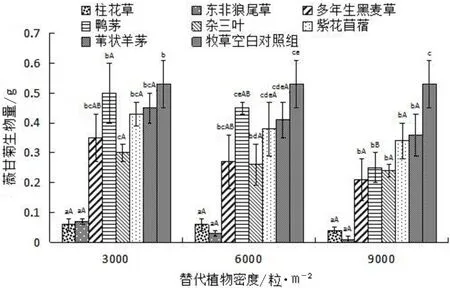
图4 不同密度7种替代植物对薇甘菊生物量的影响
2.2 林木替代控制薇甘菊的效果
由图5可知,对薇甘菊盖度和替代植物郁闭度进行方差分析比较,勃氏甜龙竹、麻竹、杉木、旱冬瓜及西南桦的郁闭度分别为98.00%、97.80%、97.2%、75.2%及30.60%,其中,勃氏甜龙竹、麻竹和杉木三者间的郁闭度差异不显著(P>0.05),旱冬瓜的郁闭度显著低于勃氏甜龙竹、麻竹和杉木的郁闭度(P<0.05),以西南桦的郁闭度最低,显著低于其它4种(P<0.05)。
替代植物的郁闭度不同,对薇甘菊的控制作用就不同,勃氏甜龙竹、麻竹、杉木、旱冬瓜、西南桦5种植物林下及对照组中薇甘菊盖度分别为0.00%、0.11%、0.12 %、10.17 %、87.80 %和90.20%,其中,旱冬瓜林下薇甘菊盖度显著小于对照组薇甘菊盖度,而显著大于勃氏甜龙竹、麻竹、杉木3种植物林下的薇甘菊盖度,而以西南桦和对照组中薇甘菊盖度最高,显著高于以上其它4种植物林下的薇甘菊盖度(P<0.05)。
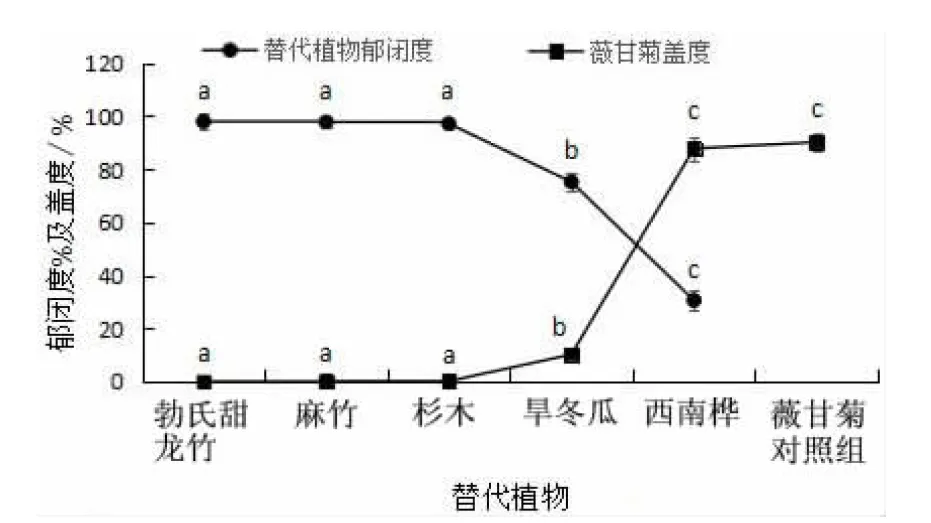
图5 不同替代植物郁闭度和林下薇甘菊盖度
3 结论与讨论
3.1 草本植物替代控制薇甘菊
植物的株高、生物量以及样方内其它植物的种类和数量是反映植物竞争力大小的重要指标[13-14]。通过分析比较所选替代植物与薇甘菊混种时的株高,可以得知,柱花草和东非狼尾草相较其它草本植物具有更强的株高优势,且二者生境中其它植物种类最少,表现出很强的环境适应性和种间竞争性,这对能够有效替代控制薇甘菊而言是相当重要的。除柱花草之外,其它替代植物样方中均有阔叶丰花草的生长,至于在特定条件下,阔叶丰花草能否用来控制薇甘菊有待进一步研究。
在试验材料及条件基本一致的情况下,分析各替代植物对薇甘菊生长情况的控制作用发现,当与牧草混种,薇甘菊的主茎长度、分枝数量和生物量均受到影响。在3个密度下,柱花草和东非狼尾草效果均为最佳,能够有效地抑制薇甘菊主茎的生长;分析同种牧草不同种植密度的影响发现,供试替代植物种类对薇甘菊几项指标的影响随着替代植物种植密度增大呈递增趋势,这与徐高峰等[13]报道随着竞争物种种植密度的增大,薇甘菊株高和分枝数受到的抑制作用明显增强相一致,研究结果证实替代植物密度对控制薇甘菊有重要影响。
总而言之,柱花草与东非狼尾草能以较低密度对薇甘菊主茎长度、分枝数量和生物量起到很强的控制作用,可以选为替代控制薇甘菊的草本植物,且在利用二者控制薇甘菊的同时,又能收获具有较高经济效益的牧草。柱花草和东非狼尾草混合播种对薇甘菊的控制作用是否增加,以及两者混种是否存在种间竞争从而影响对薇甘菊控制作用有待进一步研究。
3.2 林木替代控制薇甘菊效果评价
赵燕波等[15]指出,林冠郁闭程度高,对林下的光照、水分拦截较多,将影响林下耐阴性较弱以及需水较多的植物生长;此外郁闭度高,林下凋落物较多,对其他植物的化感作用较强,也会影响其生长。李双喜等[16]研究马褂木(Liriodendronchinense)林分郁闭度对林下草本植物的影响,结果表明,喜光性植物的平均盖度随林分郁闭度增大而减小。本研究通过调查勃氏甜龙竹、麻竹、杉木、旱冬瓜和西南桦替代控制薇甘菊试验基地发现,前4种植物形成的郁闭度显著大于薇甘菊盖度,对薇甘菊有显著的控制作用,西南桦形成的郁闭度(30%)很小,不能有效防控薇甘菊。旱冬瓜为落叶乔木,勃氏甜龙竹、麻竹和杉木的替代控制作用略强于旱冬瓜,其中,以勃氏甜龙竹效果最佳。利用勃氏甜龙竹、麻竹和杉木对薇甘菊进行替代控制,持续性较强,在防控薇甘菊的同时,还可促进树种的资源化利用,获得生态效益的同时,还能创造更大的经济效益[17-19]。
已有调查发现,台湾相思(Acaciaconfusa)-鸭脚木(Scheffleraoctophylla)+潺槁(Litseaglutinosa)群落中薇甘菊的分枝格局及生物量明显低于芒草(Miscanthussinensis)单优群落[20],采用乔木、草本等不同植物形态间作,最大限度地增加林间郁闭度,可以有效控制薇甘菊的生长。综上所述,采用柱花草、东非狼尾草、勃氏甜龙竹、麻竹、杉木对薇甘菊进行替代控制,从种植时间及植物形态方面考虑,采用乔木-草本相结合的方式,不同植物种类相互补充,最大限度地保证周年郁闭,使替代植物的种群优势显著高于薇甘菊,具有生态环保、高效防控的优点。
致谢:西南林业大学2015届硕士研究生张新瑜、徐小伟参加了本研究的全程试验,在此一并致谢。
参考文献:
[1] 李秋玲,张峰,肖辉林.外来入侵植物薇甘菊的危害现状及治理途径[J].北京农业,2011(33):129.
[2] 李娟,崔永三,宋玉双,等.我国林业检疫性和危险性有害生物新名单的特点[J].中国森林病虫,2013,32(5):42-47.
[3] 李正洪,谷芸,郭芯瑜,等.外来杂草薇甘菊在云南德宏州的危害及防控措施[J].杂草科学,2013,31(1):69-70.
[4] 李鸣光,鲁尔贝,郭强,等.入侵种薇甘菊防治措施及策略评估[J].生态学报,2012,32(10):3240-3251.
[5] 徐小伟,泽桑梓,杨斌,等.薇甘菊的分布危害、生物防治及资源化利用研究现状与展望[J].热带农业科学,2014,34(12):75-84.
[6] 刘晓燕,曹坳程,李园,等.几种除草剂对薇甘菊的防控效果[J].生物安全学报,2012,21(3):216-220.
[7] 黄云鹏,范繁荣,王邦富,等.4种不同林分类型对多花黄精生长的影响[J].西部林业科学,2016,45(5):132-135.
[8] 郭连金,徐卫红,孙海玲,等.空心莲子草入侵对乡土植物群落组成及植物多样性的影响[J].草业科学,2009,26(7):137-142.
[9] 成秀媛.幌伞枫对薇甘菊的化感作用[D].广州:中山大学,2006:25-26.
[10] 王慧.血桐的化感作用及构筑型对薇甘菊入侵的影响[D].广州:中山大学,2009:17.
[11] 申时才,徐高峰,张付斗,等.红薯对薇甘菊的竞争效应[J].生态学杂志,2012,31(4):850-855.
[12] 张国云,杨俊华,毕生斌,等.德宏州入侵杂草薇甘菊调查监测及风险分析[J].中国农学通报,2015,31(15):187-192.
[13] 徐高峰,张付斗,李天林,等.不同密度五种植物对薇甘菊幼苗的竞争效应[J].生态环境学报,2011,20(5):798-804.
[14] 张永亮,王建丽,胡自治,等.杂花苜蓿与无芒雀麦混播群落种间竞争及稳定性[J].草地学报,2007,15(1):43-51.
[15] 赵燕波,张丹桔,张健,等.不同郁闭度马尾松人工林林下植物多样性[J].应用与环境生物学报,2016,22(6):1048-1054.
[16] 李双喜,朱建军,张银龙,等.人工马褂木林下草本植物物种多样性与林分郁闭度的关系[J].生态与农村环境学报,2009,25(2):20-24.
[17] 杜凡.云南重要经济竹种特性及其生产中存在问题[J].西南林学院学报,2003,23(2):26-30.
[18] 朱凯,曹少元,陈科,等.杉木油的提取及干馏油的精制[J].农业工程学报,2012,28(9):282-286.
[19] 李大伟,陈宏伟,史富强,等.云南旱冬瓜的生物学、生态学特性及地理分布[J].林业调查规划,2008,33(5):25-28.
[20] 张炜银, 王伯荪, 李鸣光,等.台湾相思林和芒草草丛中薇甘菊枝构件的分枝格局及其生物量[J].植物生态学报,2002,26(3):346-350.
