毛乌素沙地3种典型灌木生物量分配与土壤含水量特征
2018-06-05王伟峰段玉玺李少博张立欣王博刘宗奇
王伟峰,段玉玺,李少博,张立欣,王博,刘宗奇
(内蒙古自治区林业科学研究院,内蒙古 呼和浩特 010010)
灌木生物量的组成及其特征是研究灌木群落演替及其生态系统效应的一个重要内容,也是评价植被生产力的一个重要参数,特别是研究灌木生物量与土壤养分和水分的耦合关系对指导荒漠生态系统的植被恢复具有重要科学意义[1-3]。地处农牧交错带的毛乌素沙地是我国北方重要的生态屏障,也是我国四大沙地之一[4]。灌木作为毛乌素沙地的优势物种,对于毛乌素沙地的自然环境改善以及沙害治理有着重要作用[5]。目前,在灌木生物量方面已有不少学者做了研究,如巨文珍等[6]采用全部收获法分别获取灌木的枝、干、叶、根鲜重,并选择地径和高2个因子与各器官生物量拟合,分别建立了生物量模型;宁晨等[7]采用直接收获法和实测数据,以天然灌木林内木本和草本植物、凋落物及土壤为研究对象,研究了灌木林生态系统的生物量、碳含量及碳储量;曾伟生[8]对国内外灌木生物量模型的研究现状进行了综述,从数据采集、建模方法和模型评价3个方面系统总结了灌木生物量模型研建的技术要点;曾慧卿等[9]以中国科学院千烟洲生态试验站林下常见的16种物种作为研究对象,构建了单一物种以植冠面积为变量的二次方程和以植冠投影体积为变量的乘幂方程来估算物种生物量;杨昊天等[10]选取腾格里沙漠东南缘荒漠草地的4种优势灌木种,采用全挖法在个体水平上研究了各器官的生物量以及地下/地上生物量关系(根冠比),揭示了生物量在各器官中以及地下/地上的分配格局;牛海[11]以毛乌素沙地4个水分梯度植物群落为研究对象,探讨了植物地上/地下生物量分配及根冠比随生长进程而表现出的变化趋势;肖春旺[12]以毛乌素沙地沙柳(Salixpsammophila)、杨柴(Hedysarumfruticosumvar.mongolicum)、油蒿(Artemisiaordosica)和柠条锦鸡儿(Caraganakorshinskii)幼苗为研究对象,模拟了温度和降水对不同植被类型的影响;刘陟等[13]以毛乌素沙地优势灌木油蒿为研究对象,利用其易测指标株高、冠幅面积、株高与冠幅面积乘积等参数作为变量,结合实地获取的油蒿地上生物量与地下根系生物量,进行回归分析构建油蒿生物量模型。综上分析,目前对毛乌素沙地灌木生物量的研究还不够深入,特别是缺乏不同灌木生物量分配与土壤含水量特征的相关研究。因此,基于挖掘法实测毛乌素沙地主要灌木类型的生物量,并就各器官生物量分配、含水率以及土壤含水量等进行综合分析,对科学评价荒漠生态系统的生态服务功能具有重要意义。
1 研究区概况与研究方法
1.1 研究区概况
研究区位于毛乌素沙地西北缘,地理坐标为106°50′00.30″-106°51′31.03″E,38°48′34.82″-38°49′44.45″N。研究区属于典型的温带大陆性季风气候,日照丰富,四季分明,无霜期短,降水少,蒸发量大。该区年平均气温7.1℃,最热月(7月)平均气温22.4℃,最冷月(1月)平均气温-10.2℃,极端高温为36.7℃,极端低温为-36.1℃,全年≥10℃积温大约2900℃。降水主要集中在7-9月,多年平均降水量为260mm左右,年平均蒸发量2 514.8mm,蒸发量远大于降水量,是降水量的9-10倍,且降水量的季节和年际变化非常大。年平均日照2 700-2900h,无霜期134d,大风、沙尘暴天气较多,尤以春季最多,年平均风速2.8-3.8m/s,大风(≥8级)年平均25-30d。植被主要由油蒿、杨柴、柠条锦鸡儿、沙米(Agriophyllumsquarrosum)、沙鞭草(Psammochloavillosa)等植物为主的沙生灌丛和半灌丛所组成。研究区土壤类型均为风沙土,土壤矿质部分几乎全由细砂颗粒组成,剖面层次分化不明显,仅有淋溶层和母质层,缺乏沉积层,土壤风蚀严重。
1.2 研究方法
1.2.1 样地设置及标准株取样
在毛乌素沙地西北部的鄂托克旗境内选取具有代表性的3种沙生灌木群落进行样地设置,3种植被类型分别为柠条锦鸡儿群落、油蒿群落和杨柴群落,在每个植被类型中分别设置1块标准样地。样地范围内及周边地貌以起伏的沙丘为主,包括植被覆盖的固定沙地、半流动沙地和裸露的流动沙地,沙丘高度10-50m不等,沙丘形态多为复合型沙丘,其中流动沙地形态多以沙丘链和格状沙丘为主,其次为复合型沙丘,而固定沙丘的形态多为梁窝状沙丘和灌丛沙堆,迎风坡面较为平缓,背风坡面则较为陡峭,坡度为10°-45°,坡位为中下坡和丘间洼地,海拔为1320-1370m,每块样地面积为100m×100m,在每块样地内随机设置5m×5m的调查样方3个(以固定沙地为主),对每个样方进行植被群落调查,调查内容包括植物种类、高度、盖度和密度等,根据样方调查数据在每个样方内选取1株灌木作为标准株(考虑大、中、小代表性),采用电子称分别称重每一灌木标准株的根、茎、叶鲜重,然后选取部分样品用干纸包好标记后带回实验室。在实验室中置于恒温烘箱65℃条件下烘至恒重,分别称取每株标准株各器官干重,计算各器官干重占整株标准株干重的比例以及植物含水量等。此外,分别在每块样地内随机挖取3个1m×1m的土壤自然剖面,分别在0-10cm、10-20cm、20-40cm、40-60cm土层取样,采用烘干法测定土壤含水量,取每个剖面各层土壤含水量的平均值作为该样地土壤含水量值。
1.2.2 数据分析
采用Microsoft Excel 2007和SPSS 20.0进行数据整理及图件绘制。对3种灌木标准株整株和各器官生物量及其分配比例、土壤含水量等进行单因素方差分析(P<0.05为显著,P< 0.01为极显著),利用双变量法进行相关性分析。
2 结果与分析
2.1 3种沙生灌木各器官生物量分配特征
如图1 所示,研究区3种灌木标准株整株生物量与各器官生物量均为柠条锦鸡儿﹥油蒿﹥杨柴,且均达到了显著性差异水平(P<0.05),其中柠条锦鸡儿标准株整株生物量分别是油蒿与杨柴的5.42倍和10.92倍;根系生物量分别是油蒿与杨柴4.57倍和8.11倍;茎干生物量分别是油蒿与杨柴的6.11倍和11.9倍;而叶片生物量是油蒿与杨柴的4.57倍和14.88倍。3种灌木各器官生物量及生物量分配比例均为茎﹥根﹥叶,其中柠条锦鸡儿茎、根、叶各器官分配比例分别是62%、25%与13%;油蒿茎、根、叶各器官分配比例分别是53%、30%与17%;杨柴茎、根、叶各器官分配比例分别是55%、35%与10%。3种灌木根系生物量分配比例为杨柴﹥油蒿﹥柠条锦鸡儿,其中杨柴与柠条锦鸡儿根系生物量分配比例达到了显著差异;茎干生物量分配比例为柠条锦鸡儿﹥杨柴﹥油蒿,柠条锦鸡儿与油蒿茎干生物量分配比例差异显著;叶片分配比例为油蒿﹥柠条锦鸡儿﹥杨柴,油蒿叶片生物量分配比例显著高于杨柴叶片生物量的分配比例。这可能与3种灌木长期与干旱的环境相互作用,而产生不同的生存策略,致使各部分器官的复杂异速生长有关。
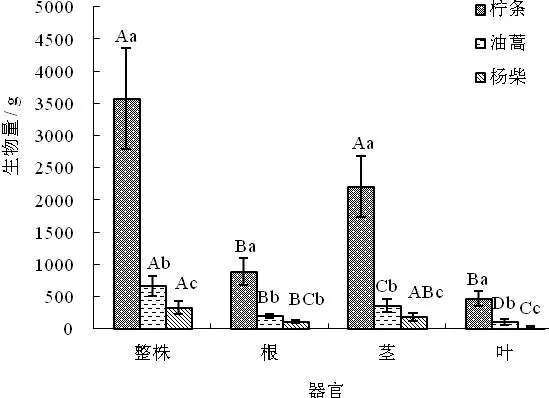
图1 3种灌木整株及各器官生物量分配特征
注:大写字母表示同一灌木不同器官间的差异性,小写字母表示同一器官不同灌木间的差异性,显著性水平P< 0.05,图2同。
Fig.1 Biomass distribution characteristics of whole plant and their organs of three shrubs
2.2 3种沙生灌木含水率特征
如图2所示,3种灌木间比较,杨柴整株及各器官含水率均显著高于柠条锦鸡儿和油蒿整株及各器官含水率(P<0.05),且3种灌木根、茎、叶各器官含水率规律具有明显差异。其中,杨柴各器官含水率为叶﹥根﹥茎,叶片的含水率是根和茎的1.22倍和1.25倍,叶片含水率显著高于根茎的含水率;柠条锦鸡儿各器官含水量与杨柴各器官含水率具有相同规律,叶的含水率分别是根和茎的1.09倍和1.36倍,根、茎、叶各器官间含水率均有显著性差异;而油蒿各器官含水率则为根﹥茎﹥叶,根含水率分别是茎和叶含水率的1.24倍和1.27倍,根系含水率显著高于茎和叶的含水率(P<0.05)。3种沙生灌木长期生长在干旱的环境中,在形态适应的同时,对其生境的资源利用也有不同的生态对策,表现出不同的生理生态特征。尤以植被保水及光合特征最为显著,3种灌木在生长发育过程中,既要进行光合作用来满足自身新陈代谢的需要,又要保持足够体内水分进行物质循环与完成自身的生理活动。杨柴和柠条锦鸡儿生长在干旱环境中,在对叶片生物量分配比例较少的情况下,为保证最大限度光合作用来满足自身新陈代谢的需要,获取更多碳水化合物,因此将体内更多水分分配到叶片中;而油蒿有些不同,在干旱环境中,对叶片生物量分配比例较大的情况下,为保证最大限度减少水分丧失的前提下促进CO2的吸收,即单位CO2收获下最小的水分散失,因此将体内较少水分分配到叶片中来完成对干旱生境的适应。
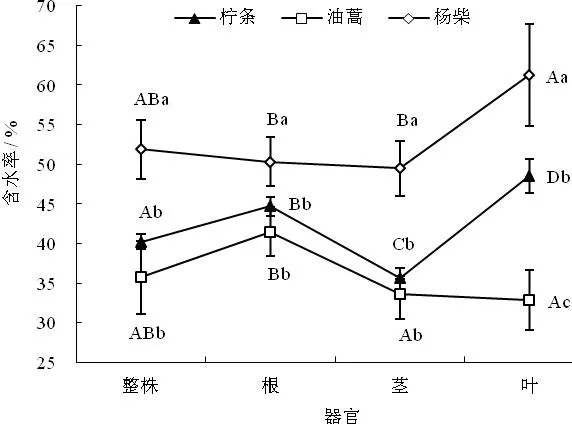
图2 3种灌木整株及各器官含水率特征
2.3 3种沙生灌木含水率与土壤含水量的相关性
如图3所示,柠条锦鸡儿、油蒿和杨柴3种灌木除叶片外(R2=0.45),3种灌木标准株整株(R2=0.74)、根(R2=0.66)的含水率均与土壤含水量存在着显著的正相关性(P<0.05),而茎(R2=0.86)的含水率与土壤含水量之间达到了极显著的正相关(P<0.01)。作为与干旱生境长期作用的灌木而言,既要保证光合作用有效进行,又要保证体内充足水分来完成正常的生理代谢活动,而用于植物光合作用所需要水分占叶片蒸腾很小一部分。因此,长期生活在干旱生境中的植物,为了适应干旱环境其叶片均具有较强的保水功能,来减少由于叶片蒸腾而造成的体内水分损耗。由于植物自身通过渗透物质对叶片含水率的反馈调节,导致叶片含水率与土壤含水量相关性不显著。在干旱生境中,植物自身不仅要从土壤中获取充足的水分,而且要有较强的保水功能,而根与茎分别是植物水分吸收、储存与运输的主要器官,它们的水分代谢活动与土壤含水量息息相关,是植物从土壤获取充足水分以及植物自身保水的基础。因此,根、茎含水率与土壤含水量呈显著正相关性。

图3 3种灌木含水率与土壤含水量的相关性
3 讨论与结论
荒漠区具有干旱、高温、强光复合逆境的特点,荒漠灌木为了适应其特定的生境条件,在长期的进化过程中形成了一系列抵御逆境的生理生态适应机制,包括形态结构特性、水势、光合特性、气孔导度、蒸腾特性、渗透调节与抗氧化保护等多种抗旱途径或方式[14]。柠条锦鸡儿、油蒿和杨柴是毛乌素沙地中3种优势沙生灌木。研究结果表明,3种沙生灌木在形态结构和水分调节利用方面采取不同的方式来适应干旱的生境。在根、茎、叶生物量分配及其比例方面,3种沙生灌木虽均表现为茎﹥根﹥叶,但各器官分配比例具有明显的差异;而在含水率方面,杨柴和柠条锦鸡儿2种灌木各器官含水率表现为叶﹥根﹥茎,而油蒿则表现为根﹥茎﹥叶,这可能是因3种灌木长期与干旱的环境相互作用,对保水与CO2吸收进行权衡,从而达到对环境最大适应而采用不同生理生态对策而造成的[15]。3种沙生灌木长期生长在干旱、炎热的环境中,在形态适应的同时,对其生境的资源利用也有不同的生态对策,表现出不同的生理生态特征,尤以植被保水及光合特征最为显著[16]。3种灌木在生长发育过程中,既要进行光合作用来满足自身新陈代谢的需要,又要保持足够体内水分进行物质循环与完成自身的生理活动[17]。杨柴和柠条锦鸡儿生长在干旱的环境,在对叶片生物量分配比例较少的情况下,为保证最大限度光合作用来满足自身新陈代谢的需要,获取更多碳水化合物,因此将体内更多水分分配到叶片中;而油蒿有些不同,在干旱环境中,对叶片生物量分配比例较大的情况下,为保证最大限度减少水分丧失的前提下促进CO2的吸收,即单位CO2收获下最小的水分散失,因此将体内较少水分分配到叶片中来完成对干旱生境的适应。
此外,为了更好适应干旱、风力与特殊土壤条件相互作用造成强烈风蚀的荒漠生境,3种沙生灌木的根、茎、叶生物量分配比例方面,均表现为茎﹥根﹥叶,这可能是由3种沙生灌木各器官行使的功能导致的[10]。茎作为沙生灌木水分营养传输及主要支撑器官,面对风蚀异常强烈的特殊生境,3种沙生灌木必须有强大的茎作为支撑才能更好地适应荒漠生境。而植物根系主要功能是进行固定与吸收,对于荒漠这一干旱以及强烈风蚀的生境来说,有强大的根系来固定植物自身及从土壤中吸收充足的水分对于沙生灌木是非常重要的。叶片作为植物器官的一部分,主要是进行光合同化以及水分蒸腾等生理生化活动,在干旱的荒漠生境中,既要进行光合作用来满足自身新陈代谢的需要,又要保持足够体内水分进行物质循环与完成自身的生理活动[18]。而不同植物在保水及光合两个方面采取不同的生态适应对策,表现为杨柴和柠条锦鸡儿2种灌木各器官含水率为叶﹥根﹥茎,而油蒿则为根﹥茎﹥叶。因此,植物的各器官生物量分配比例及含水率是沙生灌木适应干旱气候、强烈风蚀生境的有效适应策略,两者的相互影响共同决定了3种沙生灌木的生理生态习性。
参考文献:
[1] 钱洲.毛乌素沙地飞播造林植被恢复特征及土壤性质变化[D].南京:南京林业大学,2013.
[2] 邱开阳,谢应忠,许冬梅,等.毛乌素沙地南缘沙漠化临界区域土壤水分和植被空间格局[J].生态学报,2011,31(10):2697-2707.
[3] 张军红.毛乌素沙地油蒿群落土壤水分分布与动态[D].北京:中国林业科学研究院,2013.
[4] 赵宣,韩霁昌,王欢元,等.毛乌素沙漠——黄土高原过渡带土壤养分空间异质性[J].生态学报,2016,36(22):7446-7452.
[5] 苗恒录.毛乌素沙地土壤养分空间变异性初步研究[D].呼和浩特:内蒙古农业大学,2010.
[6] 巨文珍,张伟,方颖琨,等.广西石山灌木生物量估算模型的建立[J].林业调查规划,2012,37(5):1-4.
[7] 宁晨,闫文德,宁晓波,等.贵阳市区灌木林生态系统生物量及碳储量[J].生态学报,2015,35(8):2555-2563.
[8] 曾伟生.国内外灌木生物量模型研究综述[J].世界林业研究,2015,28(1):31-36.
[9] 曾慧卿,刘琪璟,冯宗炜,等.红壤丘陵区林下灌木生物量估算模型的建立及其应用[J].应用生态学报,2007,18(10):2185-2190.
[10] 杨昊天,李新荣,刘立超,等.荒漠草地4种灌木生物量分配特征[J].中国沙漠,2013,33(5):1340-1348.
[11] 牛海.毛乌素沙地不同水分梯度植物群落生物量的研究[D].呼和浩特:内蒙古农业大学,2008.
[12] 肖春旺.毛乌素沙地优势植物对全球气候变化的响应研究[D].北京:中国科学院植物研究所,2001.
[13] 刘陟,黄奇,周延林,等.毛乌素沙地油蒿生物量估测模型研究[J].中国草地学报,2014,36(4):24-30.
[14] 马成仓,高玉葆,李清芳,等.内蒙古高原锦鸡儿属植物的形态和生理生态适应性[J].生态学报,2011,31(21):6451-6459.
[15] 钱多,查天山,吴斌,等.毛乌素沙地参考作物蒸散量变化特征与成因分析[J].生态学报,2017,37(6):1966-1974.
[16] 贾晓鹏,石晓萌,王海兵.降尘过程指示的沙尘循环过程[J].中国沙漠,2012,32(3):600-603.
[17] 程栋梁.植物生物量分配模式与生长速率的相关规律研究[D].兰州:兰州大学,2007.
[18] 冯丽,张景光,张志山,等.腾格里沙漠人工固沙植被中油蒿的生长及生物量分配动态[J].植物生态学报,2009,33(6):1132-1139.
